 2010, Vol. 26
2010, Vol. 26

已有研究表明,慢性应激可影响脑的认知功能,尤其是学习记忆功能,导致学习和记忆损害〔1〕。研究发现,适度的体力训练有益于学习记忆等脑功能〔2〕。细胞外信号调节蛋白激酶(extracellular signal-regulated protein kinase,ERK)通路对突触的可塑性和长时程记忆的形成起重要作用;环磷酸腺苷反应元件结合蛋白(cAMP response element binding protein,CREB)是ERK通路中重要的下游转录靶信号,介导了短时程记忆向长时程记忆的转化,而海马、额叶皮质、纹状体都是与学习记忆相关的脑区,本研究采用慢性束缚应激模型,观察运动对应激大鼠不同脑区ERK-CREB信号通路的变化,探讨运动提高慢性应激状态下学习记忆能力的机制。
1 材料与方法 1.1 试剂与仪器兔抗p-ERK及p-CREB单克隆抗体、生物素标记羊抗兔IgG (北京中山公司);链亲和素-过氧化物酶复合物(武汉博士德公司)。Olympus BX-51显微镜(日本奥林巴斯公司);LEICA-CM1850冷冻切片机(上海徕卡仪器有限公司)。
1.2 动物与分组雄性SD大鼠(四川大学实验动物中心) 40只,2~3月龄,体重(210 ± 15) g。随机分为4组:对照组、应激组、运动+应激组、运动组,每组10只。所有动物自由摄食和饮水。实验室温度20~22℃、湿度50%,室内光/暗周期为12 h。
1.3 慢性应激模型及运动方案采用慢性束缚方式建立应激模型〔3〕,6 h/d,共21 d。运动方案为无负重游泳〔4〕,60min/d,总运动时间8周。应激组、运动+应激组在8周运动结束1 d后,开始21 d束缚应激造模。运动组在8周运动结束后,同期饲养22 d,对照组同期饲养。
1.4 取材造模结束1 d后,10%水合氯醛(30mg/kg)腹腔注射麻醉大鼠,断头处死,剥离脑组织,经矢状位切开,随机取一侧半球,迅速置于冷冻切片机中,行15 μm连续矢状切片,取其中包含海马、纹状体及额叶皮层等脑结构的切片,置于 -20℃冰箱中备用。
1.5 指标检测免疫组化染色检测海马、纹状体及额叶皮层 p-ERK及p-CREB。正常羊血清37℃孵育20 min;加入兔抗 p-ERK及p-CREB单克隆抗体,37℃孵育30 min,4℃冰箱过夜;滴加生物素标记羊抗兔IgG,37℃孵育40 min;滴加链亲和素-过氧化物酶复合物,37℃孵育30 min;二氨基联苯胺室温显色,终止反应。图像采集和分析分别采用SPOT ADVANCE和IMAGE-PRO-PLUS分析软件,每张切片随机选取 4个高倍视野(400 ×),统计每个视野中阳性细胞面积、密度、吸光度(A),以总吸光度值进行统计。
1.6 统计分析采用SPSS13.0软件分析数据,组间两两比较采用单因素方差分析。
2 结 果 2.1 各组大鼠不同脑区p-ERK表达变化(表 1)| 表 1 各组大鼠海马、额叶皮层、纹状体p-ERK 变化(x ± s,A 值,× 10 000) |
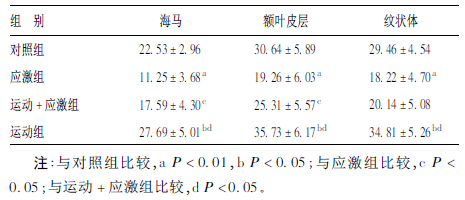
与对照组比较,应激组海马、额叶皮层、纹状体p-ERK表达明显下降,差异有统计学意义(P < 0.01)。与应激组比较,运动+应激组海马、额叶皮层p-ERK表达明显升高(P < 0.05)。与运动组比较,运动+应激组各脑区p-ERK表达仍明显降低(P < 0.05)。运动组各脑区p-ERK变化高于对照组,差异有统计学意义(P < 0.05)。
2.2 各组大鼠不同脑区p-CREB表达变化(表 2)| 表 2 各组大鼠海马、额叶皮层、纹状体p-CREB(x ± s,A 值,× 10 000) |
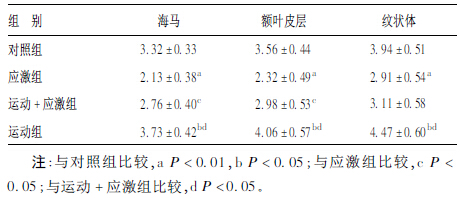
与对照组比较,应激组海马、额叶皮层、纹状体区p-CREB表达明显降低(P < 0.01)。运动+应激组海马、额叶皮层p-CREB表达较应激组明显升高(P < 0.05)。运动+应激组各脑区p-CREB仍明显低于运动组(P < 0.05)。运动组各脑区p-CREB表达较对照组明显升高(P < 0.05)。
3 讨 论ERK广泛分布于中枢神经系统,在海马和前额叶皮质的表达尤其显著,其磷酸化形式能够介导胞外信息向胞核内转移,在调节神经元的增殖、分化和提高学习记忆等方面起重要作用〔5, 6〕。CREB是ERK最重要的下游信号分子之一,其调节转录活性及磷酸化与细胞内长期记忆的构成密切相关〔7〕。 ERK信号通路在调节CREB磷酸化方面起关键作用,这种 ERK活化伴随CREB的活化同时也支持诸如长时程增强的适应过程〔8, 9〕。本研究结果表明,应激组海马、额叶皮层、纹状体区p-ERK/p-CREB表达均低于对照组(P < 0.01),表明 ERK-CREB通路下调可能参与了慢性应激致学习记忆受损的机制。比较运动+应激组和应激组结果,8周运动明显上调p-ERK/p-CREB在海马及额叶皮层的表达(P < 0.05),提示运动可能通过激活细胞内ERK通路,p-ERK经核转位进入细胞核后,对其下游的CREB进行磷酸化从而增强多种与学习记忆相关的下游靶基因的转录,在一定程度上达到减缓应激性记忆损伤的作用。此外,运动组p-ERK/p-CREB表达较对照组明显升高(P < 0.05),表明运动能使中枢神经系统 ERK与CREB的磷酸化水平持续升高,通过目标基因的表达使脑学习记忆功能发生有益变化。
| 〔1〕 | Wright RL,Conrad CD.Chronic stress leaves novelty-seeking behavior intact while impairing spatial recognition memory in the Y-maze[J].Stress,2005,8(2):151-154. |
| 〔2〕 | Chen L,Gong S,Shan LD,et al.Effects of exercise on neurogenesis in the dentate gyrus and ability of learning and memory after hippocampus lesion in adult rats[J].Neurosci Bull,2006,22(1): 1-6. |
| 〔3〕 | Wood GE,Young LT,Reagan LP,et al.Acute and chronic restraint stress alter the incidence of social conflict in male rats[J]. Horm Behav,2003,43(1):205-213. |
| 〔4〕 | 张银娥,宋辉,杨惠芳,等.运动对慢性心理应激大鼠影响[J]. 中国公共卫生,2007,23(4):472-473. |
| 〔5〕 | Li L,Yun SH,Keblesh J,et al.Egr3,a synaptic activity regulated transcription factor that is essential for learning and memory[J]. Mol Cell Neurosci,2007,35(1):76-88. |
| 〔6〕 | Feld M,Dimant B,Delorenzi A,et al.Phosphorylation of extra-nuclear ERK/MAPK is required for long-term memory consolidation in the crab Chasmegnathus[J].Behav Brain Res,2005,158(2): 251-261. |
| 〔7〕 | 文涛,刘阳,孙黎光,等.慢性铅暴露对小鼠pCREB蛋白表达影响[J].中国公共卫生,2007,23(5):602-603. |
| 〔8〕 | Song XS,Xu YB,Cao JL,et al.cAMP response-element binding protein participates in the phosphorylated extracellular signal-regulate kinase mediated neuropathic pain[J].Sheng Li Xue Bao, 2005,57(2):139-146. |
| 〔9〕 | Grinkevich LN,Lisachev PD,Baranova KA,et al.Comparative analysis of the activation of MAP/ERK kinases in the CNS of animals with different learning abilities[J].Neurosci Behav Physiol, 2007,37(7):715-720. |