 2010, Vol. 26
2010, Vol. 26

 , 付萍, 洪牧
, 付萍, 洪牧 家蝇(Mu sca dom estica)是兼性滞育昆虫,又是重要的医学昆虫。现己证实其体内具有独特的免疫防御系统,可产生具有抗细菌、抗病毒、抑肿瘤细胞、原虫等多种生物活性〔1〕。利用家蝇幼虫血淋巴进行免疫研究的报道很多,但国内外诱导家蝇3龄末期幼虫滞育的研究正在不断深入。本研究拟利用人工条件(温度和光周期)诱导家蝇3龄幼虫滞育,观察温度、光周期和亲代经验对子代滞育的影响,优化诱导家蝇幼虫滞育的条件,为滞育家蝇幼虫免疫活性物质的进一步研究提供理论和实验依据。
1 材料与方法 1.1 材料家蝇(M usca dom estica)为本实验室饲养传代,幼虫按常规饲养至3龄(孵化后5 d)备用。
1.2 方法 1.2.1 滞育时间标准的确立收集25℃、长日照3 h之内的家蝇卵,分别放入不同温度(25,20,17.5,15℃)与不同光周期(长日照L 16:D 8,短日照L 12:D 12)各组合条件下饲养,每12 h观察一次幼虫进食情况,记录幼虫停止进食时间。设立25℃下幼虫停止进食到成蛹的最长时间为对照。同上收集家蝇卵,随机分为5组,(1) L 4:D 20;(2) L 8:D 16;(3) L 12:D 12;(4) L 16:D 8;(5) L 20:D 4。温度均为25#,计算幼虫停止进食到成蛹的时间。同上设立对照组。
1.2.2 家蝇幼虫滞育的判断标准幼虫历期是正常发育幼虫最短历期的3倍以上为滞育〔2〕。
1.2.3 亲代、子代的光周期和温度对子代滞育的影响按饲养条件的不同将亲代随机分为4个组:(1)25℃,L 16:D 8; (2)25℃,L 12:D 12;(3)20℃,L 16:D 8;(4)20℃,L 12:D 12。分别收集各组家蝇卵,将子代分别饲养于2个光周期(L 16:D 8,L 12:D 12)和4个温度(25,20,17.5,15℃)共32种组合条件下,计算各组幼虫滞育率。每个条件重复3次。
1.2.4 温度对家蝇幼虫滞育的解除将进入滞育状态7 d的幼虫随机分为20℃和25℃2组,光周期均为L 12:D 12,计算各组幼虫48 h内的成蛹率。
1.3 统计分析利用SPSS 10.0软件对数据进行单因素多变量方差分析,分析各个因素对子代幼虫滞育的影响。
2 结 果 2.1 温度和光照与幼虫停止进食的关系在25℃、不同光周期下3龄幼虫停止进食时间均为3.5 d,提示停止进食时间和光周期无关,只与温度有关。
2.2 发育速率方程根据不同温度下幼虫停止进食的时间计算发育速率,利用线性回归绘制温度和发育速率关系方程: y=-0.182+0.01856x,r2=0.996。根据方程计算出不同温度条件下幼虫的发育速率和最低生长温度,当发育速度率为 0时的温度是9.81℃。
2.3 确定进入滞育状态的时间(表 1)| 表 1 不同温度下滞育总时间 |
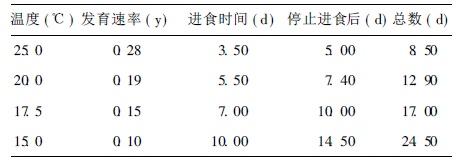
25℃条件下正常3龄幼虫停止进食后5 d内均成蛹,故将25℃条件下,停止进食后5 d还未成蛹的3龄家蝇幼虫确认为已经进入滞育状态。以此为标准,根据有效积温公式计算出其他温度下正常 3龄幼虫从停止进食到成蛹的时间。
2.4 亲代、子代的光周期和温度对子代滞育的影响4种培养条件下的亲代所产子代,在不同温度和光周期的32种组合条件下培养后,其3龄幼虫滞育结果提示:L 12:D 12和两代均低温通常会有高滞育率,亲代和子代都在L 12:D 12(20 ℃)时幼虫滞育率较高;两代长日照或/和两代高温的情况下饲养的幼虫滞育率几乎为0。经单变量多因素方差分析,组合中的4个因素(亲代和子代的光周期、温度)均会分别对子代幼虫的滞育率产生影响(P < 0.01),亦受亲代光周期和子代温度联合作用影响(P < 0.01)。提示人工诱导家蝇幼虫滞育的最佳方案为亲代20℃,L 12:D 12培养;其子代在15℃,L 12:D 12条件下培养会得到较高的滞育率。
2.5 温度对解除滞育的影响25℃条件下,停止滞育的幼虫第10 h开始成蛹,第18 h达到高峰,到第24 h,成蛹率达到 88.28%,第31 h则完全成蛹。在20℃条件下,幼虫在第19 h开始成蛹,第24 h成蛹率为2.28%,第31 h为41.09%,此时 25℃下已完全成蛹,48 h成蛹率为96.48%。提示随着温度的升高,滞育幼虫发育为蛹的时间缩短。
3 讨 论昆虫滞育的环境影响因素很多,包括光照、温度、湿度、水、食料等。有研究报道,光周期是诱导滞育的关键,而低温是维持滞育的关键,如丽蝇滞育幼虫的短期维持的温度是11℃〔3〕,长期维持的温度是4℃,滞育事件可以维持到150 d〔4〕;对草地螟幼虫,滞育的最有效光周期是L 12:D 12,随着温度的升高,临界光周期呈缩短趋势〔5〕。本结果表明,25℃下,昆虫停止进食的时间与光周期无关,只与温度有关。本观察中 25℃下幼虫从孵化到进入滞育状态共需8.5 d的时间。
实验结果还表明,亲代的短日照较长日照更有利于子代幼虫滞育,亲代的低温环境比高温更容易使幼虫滞育。子代幼虫本身的短日照亦增加了滞育机会。统计分析发现,亲代在短日照低温条件下产卵,子代在短日照15℃条件下滞育率最高,得到滞育幼虫大约需50 d。要加快速度,亲代条件不变,子代在20℃下喂养也有很好效果,时间可以缩短12 d,滞育率达到82%~100%。如果将家蝇长期在短日照低温下饲养,省去培养亲代的时间,得到滞育幼虫的时间将大大缩短,但具体条件还需进一步研究。本实验观察到,将滞育幼虫从12℃转入25℃和20℃环境后,2 h以上才开始活动,过一段时间后开始成蛹。幼虫25 ℃条件开始成蛹时间较20℃早了9 h,可见幼虫结束滞育后的发育速率与温度有关。
| 〔1〕 | Gu LJ,Wu WJ,Su XQ,et a.l Isolat ion and pu rification of novel antifungal peotides from hemolymph of mimunized larvae of housefly,Musca domestica[J].Korean Joumal of Life Science,2006,16(3):387-395. |
| 〔2〕 | Yata O,Shmia H,Saigusa T,et a.l Photoperiodic response of four Japanese species of the genus Pieris Pieris(Lepidoptera:Pieridae)[J].Kontyu,1979,47(2):185-190. |
| 〔3〕 | Chemysh I,Kmi SI.Antiviral and antitum or peptides from in sects[J].PANS,2002,99:12628-12632. |
| 〔4〕 | Chemysh SI,Natalia AF,Natalia SC,et al. Cytotoxic activity of blow fly Calliphora vicina hemocytes[J].Joumal of Insect Physiology,2004,50:777-781. |
| 〔5〕 | 黄少虹,江幸福,罗礼智.光周期和温度对草地螟滞育诱导的影响[J].昆虫学报,2009,52(3):274-280. |