 2010, Vol.26
2010, Vol.26

 , 宋春花, 李艳青, 王淑玲, 张卫东, 郗园林
, 宋春花, 李艳青, 王淑玲, 张卫东, 郗园林 幽门螺杆菌MEL2HP27 (HP27) 、PET30 (郑州大学公共卫生学院分子流行病教研室保存) ;大肠埃希 菌( Escherichia coli) DH5a、BL21 (郑州创生生物工程有限公 司) ; pMD18 - T载体(日本TaKaRa公司) 。
1.1.2 主要试剂和设备基因组DNA提取试剂盒、小质粒 提取试剂盒、凝胶回收试剂盒(天根生物有限公司) ; T4DNA 连接酶试剂盒和限制性内切酶NdeⅠ、XhoⅠ (日本TaKaRa 公司) ;镍离子纯化柱(美国Invitrogen公司) ;凝胶图像分析 仪(美国基因有限公司) ; PCR仪(德国Biomtre公司) ;台式恒 温振荡器(上海跃进医疗器械厂) ; TGL - 16G台式高速离心 机(上海安亭科学仪器厂) 。
1.2 方法 1.2.1幽门螺杆菌Hp27基因组提取 将保存的幽门螺杆 菌涂板培养后,使用DNA提取试剂盒,参照说明提取DNA。
1.2.2 PCR扩增目的基因及片段回收 参照美国国立生物 技术信息中心(NCB I)中幽门螺杆菌超氧化物歧化酶( Fe2 SOD)基因序列,用软件p rimier 510自行设计基因的引物P1: 5’C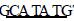 TTTACTTTACGAGAG3’; P2:
TTTACTTTACGAGAG3’; P2: 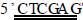 TTTATG2 CACCAACTCA3’。P1引物5’端引入酶切位点NdeⅠ (划线部 分) ,P2引物5’端引入酶切位点XhoⅠ (划线部分) 。引物由 赛百盛生物公司合成。PCR扩增程序: ( 1) 9410 ℃,5 min; (2) 9410 ℃,30 s; (3) 5611 ℃,30 s; ( 4) 7210 ℃,70 s; ( 5) 7210 ℃,10 min。重复(2) ~ ( 4) 35次。将PCR产物进行琼 脂糖凝胶电泳,使用凝胶回收试剂盒,参照说明回收目的片 段。
TTTATG2 CACCAACTCA3’。P1引物5’端引入酶切位点NdeⅠ (划线部 分) ,P2引物5’端引入酶切位点XhoⅠ (划线部分) 。引物由 赛百盛生物公司合成。PCR扩增程序: ( 1) 9410 ℃,5 min; (2) 9410 ℃,30 s; (3) 5611 ℃,30 s; ( 4) 7210 ℃,70 s; ( 5) 7210 ℃,10 min。重复(2) ~ ( 4) 35次。将PCR产物进行琼 脂糖凝胶电泳,使用凝胶回收试剂盒,参照说明回收目的片 段。
构建重组质粒PMD18 - SOD转化DH5a 参照PMD18 - T载体试剂盒(日本TaKaRa公司)说明,将凝胶回收的目 的基因与载体连接并转化到DH5a大肠埃希菌中。经蓝白斑 筛选,挑取阳性菌株做菌液PCR鉴定,正确后,送北京三博生物公司测序。
1.2.4 重组质粒PET30 - SOD的构建 1.2.4.1 PMD18 - SOD和pET30 ( a)的酶双切与回收提取 经测序鉴定正确的重组质粒PMD18 - SOD和载体质粒pET30 ( a) ,并对重组质粒和载体质粒双酶切回收产物(载体质粒用 018%琼脂糖凝胶回收,目的基因用112%琼脂糖凝胶回收) 。 重组质粒PMD18 - SOD 和载体质粒pET30 ( a)用Nde Ⅰ和 XhoⅠ双酶切的反应总体积为100 μL,各组分为: NdeⅠ 14 μL; XhoⅠ 14μL; 10 ×Buffer 28μL;重组质粒PMD18 - SOD 或载体质粒pET30 ( a) 90 μL; ddH2O 54 μL。具体步骤参照 内切酶使用说明。酶切过夜后,使用018%琼脂糖凝胶,参照 凝胶回收试剂盒说明回收目的片段。
1.2.4.2 目的基因片段与pET - 30a质粒的连接与转化经 NdeⅠ和XhoⅠ双酶切后,将回收的产物进行连接,具体步骤 参照T4DNA连接酶试剂盒,16 ℃连接过夜。将连接产物- 重组质粒PET302sod转化到DH5a大肠埃希菌中,均匀地涂抹 到含卡那霉素的LB培养板上。挑取阳性菌株做PCR鉴定,鉴定正确后送北京三博生物公司测序。
1.2.5 SOD蛋白表达和纯化将测序鉴定正确的重组质粒 PET30 - SOD转化到表达宿主菌E•coli BL21 (DE3) ,用卡那 霉素筛选阳性克隆,PCR鉴定后,首先参照《现代分子生物学 实验技术》〔6〕进行目的蛋白的优化诱导表达。经优化后,对 目的蛋白做可溶性检测,参照美国Invitrogen公司镍离子纯化 柱操作手册进行分离纯化。取分离纯化的目的蛋白50μL,进行十二烷基硫酸钠- 聚丙酰胺( SDS - PAGE)凝胶电泳。
2 结 果 2.1 幽门螺杆菌sod基因克隆与鉴定(图 1)以HP27基因 染色体DNA为模板,PCR扩增获得约640 bp的DNA片段,经 琼脂糖电泳获得预期大小目的基因片段,表明SOD基因克隆 成功。
 | 注: 1: 2 000 bp marker; 2:退火温度5611 ℃的产物; 3:退火温度5713 ℃的产物; 4:退火温度5815 ℃的产物。 图 1 SOD基因PCR扩增 |
将目的 基因与载体连接并转化到DH5a大肠埃希菌中,转接到涂有氨苄青霉素、5 - 溴- 4 - 氯- 3 - 吲哚- β - D - 半乳糖苷、异 丙基-β - D - 硫代半乳糖苷( IP IG)的培养板上,经蓝白斑筛 选,挑取阳性菌株增菌后进行菌液PCR。琼脂糖电泳获得预 期大小目的基因片段,并送北京三博生物公司测序。测序显 示,MEL - HP27 SOD基因编码序列长度为639 bp,编码213 氨基酸残基,与美国国家生物技术信息中心收录的幽门螺杆 菌SOD基因比较,核苷酸同源性达97% ~,而所编码的蛋白 质氨基酸序列同源性达100%。表明重组质粒PMD18 - SOD 构建成功。
 | 注: 2 000 bp marker; 2~4:不同菌株PCR产物。 图 2 重组质粒PMD 182sod菌液PCR鉴定 |
提取重组质粒PMD18 - SOD和载体质粒PET30 (a) ,双酶切过夜,再连接过夜转化DH5a大肠埃希菌,在含卡那霉素的培养板 上培养,挑取阳性克隆菌株进行菌液PCR检测,经琼脂糖电 泳获得预期大小目的基因片段(图 4中,下方模糊条带可能 是由于引物加入过多而产生的二聚体,对实验无影响,不予考 虑) ,并送北京三博生物公司测序,序列完全正确。表明重组 质粒PET30 - SOD的构建成功。
 | 注: 1:PMD 18-sod 双酶切;2:PET30(a)双酶切;3:PET30(a)质粒;4:15000bp marker。 图 3 PMD 18-SOD与PET30(a)双酶切电泳 |
 | 注: 2000bp matker;2~6:为不同菌株PCR产物 图 4 重组质粒PET30-菌SOD液PCR鉴定 |
提取重组质粒PET30 - sod转化到表达宿主菌BL21。按照优化的条件大量 诱导表达并通过镍离子纯化柱分离提纯。经SDS - PAGE凝 胶电泳获得预期大小目的条带,表明表达的SOD蛋白正确。
 | 注:M: 9712 KD蛋白marker; 1:纯化前的蛋白电泳; 2, 3:纯化后的蛋白电泳。 图 5 纯化前后SOD蛋白电泳 |
本研究表明,以本教研室分离并保存的幽门螺杆菌MEL - HP27的DNA组为模板,成功克隆出639 bp的幽门螺杆菌 超氧化物歧化酶( Fe - SOD)基因,并成功地将Fe - SOD基因 克隆到pET30a ( + )质粒中。测序表明,Fe - SOD基因处于正 确的阅读框架内。IPTG诱导表达表明,Fe - SOD得到高效表 达,其表达目的蛋白达到全菌蛋白含量的75%。SOD是普遍 存在于生物体内的能清除超氧阴离子自由基的一类金属酶,对于Hp在氧化应激状态下的生长和存活非常重要〔7, 8〕。按 其结合的金属离子的不同,主要可分为Cu /Zn - SOD、Mn - SOD和Fe - SOD 3种〔9, 10, 11〕。本实验中,幽门螺杆菌SOD蛋白 为Fe - SOD,序列分析发现,SOD无信号肽序列,为非分泌型 蛋白。其晶体结构最明显的特点是它的活性部位结构保守,尤其在二聚体表面相互作用部位非常保守。与其他有生物活 性的Fe - SOD结构比较,幽门螺杆菌Fe - SOD有20个C - 末端尾巴,呈α - 螺旋结构。可以推测,这个结构与SOD吸 附在细菌表面有关,并且也可能是SOD磷酸化的部位。至于 Hp SOD蛋白翻译后修饰的意义,以及与其定位的关系还需 进一步研究〔12, 13, 14〕。
| 〔1〕 | Parsonnet J, Friedman GD, Vandersteen DP, et al.Helicobacter pylori infection and the risk of gastric carcinoma[J]. N Engl JMed, 1991, 325 (16) : 11127 - 11131. |
| 〔2〕 | Huang ZG, Duan GC, Fan QT, et al.Mutation of cytotoxin2associat2 ed gene A affects exp ressions of antioxidant p roteins of Helicobacter pylori[J]. World J Gastroenterol, 2009, 15 (5) : 599 - 606. |
| 〔3〕 | AkopyantsNS, Clifton SW, Kersulyte D, et al.Analyses of the cag pathogenicity island of Helicobacter pylori[J]. MolMicrobiol, 1998, 28 (1) : 37 - 53. |
| 〔4〕 | Hsu P I, Hwang IR, CittellyD, et al.Clinical p resentation in relation to diversity within the Helicobacter pylori cag pathogenicity island [J]. Am J Gastroenterrol, 2002, 97 (9) : 2231 - 2238. |
| 〔5〕 | 段秀杰, 闻平.幽门螺杆菌毒力相关蛋白研究进展[J].世界感 染杂志, 2006, 6 (2) : 179 - 181. |
| 〔6〕 | 卢圣栋.现代分子生物学实验技术[M].北京:高等教育出版 社, 1993: 412 - 414. |
| 〔7〕 | Seyler RJ, Olson JW, Maier RJ.Superoxide dismutase2deficientmu2 tants of Helicobacter pylori are hypersensitive to oxidative stress and defective in host colonization[J]. Infect Immun, 2001, 69 ( 6 ) :4034 - 4040. |
| 〔8〕 | Wang G, Alamuri P, Maoer RJ.The diverse antioxidant systems of Helicobacter pylori[J]. MolMicrobiol, 2006, 61 (4) : 847 - 860. |
| 〔9〕 | 陈惠芳, 王琦, 付学池, 等.超氧化物歧化酶( SOD)的分子生物 学[J].生命的化学, 2003, 23 (4) : 291 - 293. |
| 〔10〕 | 周赞虎, 刘仁海, 章军, 等.超氧化物歧化酶基因突变在大肠埃 希菌中表达[J].中国公共卫生, 2004, 20 (9) : 1052 - 1053. |
| 〔11〕 | 陆耀良, 陶炳根.超氧化物歧化酶的生物学作用及其酶活性的 检验方法[J].中国公共卫生, 1995, 11 (6) : 281 - 284. |
| 〔12〕 | Ernst FD, Homuth G, Stoof J, et al.Iron2responsive regulation of the Helicobacter pylori iron2cofactored superoxide dismutase SodB ismediated by Fur[J]. J Bacteriol, 2005, 187 (11) : 3687 - 3692. |
| 〔13〕 | Carpenter BM, Gancz H, Gonzalez2Nieves RP, et al.A single nuc2 leotide change affects fur2dependent regulation of sodB in H.pylori [J]. PLOS ONE, 2009, 4 (4) : e5369. |
| 〔14〕 | Esposito L, Seydel A, Aiello R, et al.The crystal structure of the su2 peroxide dismutase from Helicobacter pylori reveals a structured C2 terminal extension[J]. Biochim Biophys Acta, 2008, 1784 ( 11 ) : 1601 - 1606. |