 2009, Vol. 25
2009, Vol. 25

 , 叶丽平1, 曹师承2
, 叶丽平1, 曹师承2 2. 中国医科大学基础医学院生物化学与分子生物学教研室
重金属铅是一种普遍存在的环境神经毒物, 具有很强的神经亲和性。目前铅神经毒作用机制研究中的一个重要内容是探讨铅对中枢神经细胞信号转导过程中蛋白激酶功能的影响〔1〕。本课题组前期研究发现, 急、慢性铅中毒可以扰乱正常小鼠整体脑海马及脑片细胞外信号调激酶(ERK2)活性〔2〕。本研究进一步使用Ca2+/钙调蛋白依赖的蛋白激酶Ⅱ (Ca2+/Calmodulin dependent protein kinaseⅡ, CaMKⅡ)的激活剂、抑制剂KN-93及铅对大鼠海马脑片进行处理, 通过检测CaMKⅡ的下游信号分子ERK2活性及表达总量来探讨CaMKⅡ在大鼠海马急性铅中毒中作用机制, 以期寻求新的治疗铅中毒靶点。
1 材料与方法 1.1 实验材料Wistar大鼠(中国医科大学实验动物中心)。醋酸铅(沈阳化学试剂厂, 分析纯)。兔磷酸及非磷酸化ERK2多克隆抗体(上海Promega公司); 辣根过氧化酶标记的山羊抗兔二抗(北京中山生物技术有限公司); 牛血清白蛋白(联星生物工程公司); KN-93(上海Sigma公司); 谷氨酸(上海试剂厂)。Alpha Innotech Corporation 5500型自动凝胶成像分析系统及ChemiImager 5500 V 2.03(美国自然基因有限公司)。人工脑脊液〔ACSF (mmo l/L) NaCl 124, KCl 4.4, CaCl2 2.5, MgSO4 1.3, NaH2PO4 1, NaHCO3 26, 葡萄糖10〕。
1.2 实验步骤 1.2.1 大鼠脑片培养出生30 d健康大鼠, 颈部脱臼法处死, 迅速取出海马, 放入预冷的ACSF液中。将海马横切为350 μm脑片放到6孔培养板中培养。用预先通以95%O2 + 5%CO2的混合气体的ACSF灌流, 流速1 mL/min, 温度32.5~33.5 ℃。稳定培养2 h。将培养的大鼠海马脑片放到0.5 g/L四甲基偶氮噻唑蓝(MTT)中, 室温观察15 min, 如颜色由白变黑则证明存活〔3〕。
1.2.2 谷氨酸、KN-93及铅处理培养脑片6孔培养皿ACSF液稳定培养2 h后分组:对照组为普通ACSF液; 加谷氨酸的ACSF液组(谷氨酸终浓度为5 μmol/L); 加KN-93的ACSF液组(KN-93终浓度为2 μmol/L 〔4〕); 加谷氨酸及醋酸铅的ACSF液组(谷氨酸终浓度为5 μmol/L, 醋酸铅浓度为20 μmol/L), 培养30 min后收集脑片。
1.2.3 ERK2活性测定将收集的样品放到4 ℃预冷的裂解缓冲液中。4 ℃超声粉碎后, 20 000 g离心60 min, 取上清分装。用酚试剂法测蛋白含量, 以牛血清白蛋白为标准。用10%的十二烷基磺酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE)分离蛋白质, 每孔蛋白含量为30 μg。用磷酸化抗体, 常规蛋白免疫印迹(Western blots)法测定ERK2的磷酸化状态下活性及总量的表达。将蛋白印迹显影图扫描, 再利用Chemi Imager 5500 V 2.03图像分析软件进行分析。灰度值代表酶含量, 以对照组酶含量为100, 其他组与其对比。
1.3 统计分析采用SPSS 11.5软件统计对实验数据经内参校正后进行方差分析。
2 结果 2.1 谷氨酸对ERK活性及总量ERK表达的影响(图 1, 表 1)

|
注: 1:对照; 2:谷氨酸; 3: K N-93; 4:谷氨酸+醋酸铅。 图 1 Western blots测定ERK2活性及表达结果 |
| 表 1 CaMKⅡ活性改变对ERK2活性及表达的影响(x±s, n=3) |
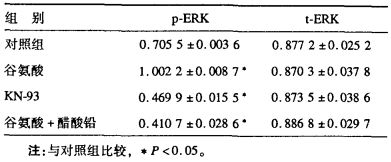
谷氨酸通过激活CaMKⅡ活性使磷酸化的ERK2活性与对照组比较增加42%, 差异有统计学意义(P < 0.05);对非磷酸化的ERK2表达无明显影响。
2.2 KN-93抑制CaMK Ⅱ活性对ERK活性及总量ERK表达的影响(图 1, 表 1)KN-93通过抑制CaMKⅡ活性, 使磷酸化的ERK2活性比对照组降低23%, 差异有统计学意义(P < 0.05);对总量ERK2表达无明显影响。
2.3 铅对ERK2活性及总量E RK表达的影响(图 1, 表 1)铅可拮抗谷氨酸的作用, 降低了谷氨酸引起的ERK2活性升高使磷酸化的ERK2活性与对照组比较降低28%, 差异有统计学意义(P < 0.05);对总量ERK2表达无明显影响。
3 讨论铅是环境中影响婴幼儿智力发育、儿童学习记忆功能的一种重要的神经毒, 铅中毒机制的研究一直是铅中毒防治领域的热点问题, 尽管获得了大量的研究结果, 但其机制仍未完全清楚。
目前研究表明, CaMKⅡ在脑组织中高度表达, 是突触后颗粒的主要成分, 而学习记忆的理想模型LTP 〔5〕的诱导过程中, 研究突触后成分变化发现, 胞浆CaMKⅡ的Thr 286磷酸化而被激活, 这种磷酸化可以使其在Ca2+浓度下降情况下依然保持活性, 因此, 被认为是学习记忆的分子基础〔6〕。而铅在无Ca2+存在下也可作用于钙调素〔7〕, 进而影响CaMKⅡ的活性, 抑制LTP的产生〔8〕。因此, 认为该酶的正常变化被扰乱可能是铅导致其学习记忆损害的原因之一〔9〕。本研究结果显示, 铅可能通过影响CaMKⅡ的活性影响其下游信号分子的活性状态, 使其在铅中毒导致学习记忆损伤中占有重要位置, 可能成为防治铅中毒新的靶点, 为铅中毒的防治提供新的理论依据。
| [1] | Bay KU, De Koninck P, Leonard AS, et al. Interaction with the NMDA receptor locks CaMKⅡ in an active conformation[J]. Nature, 2001, 411(6839) : 801–805. DOI:10.1038/35081080 |
| [2] | 侯伟健, 彭博, 吴哲, 等. 铅对小鼠脑细胞外信号调节激酶活力的影响[J]. 卫生毒理学, 2003, 17(2) : 219–221. |
| [3] | Blum S, Moore AN, Adams F, et al. A Mitogen-activated protein kinase cascade in the CA1/CA2 subfield of the dorsal hippocampus is essential for long-term spatial memory[J]. J Neurosci, 1999, 19(9) : 3535–3544. |
| [4] | 杨红卫, 胡晓东, 张红梅, 等. KN-93抑制脊髓背角C-纤维诱发电位LTP的诱导和早期维持[J]. 中山大学学报:医学科学版, 2003, 24(4) : 321–328. |
| [5] | Holscher C. Synaptic plasticity and learning and memory: LTP and beyond,[J]. J Neurosci Res, 1999, 58(1) : 62–75. DOI:10.1002/(SICI)1097-4547(19991001)58:1<>1.0.CO;2-I |
| [6] | Lisman J, Schulman H, Cline H. The molecular basis of CaMKⅡ function in synaptic and behavioural memory[J]. J Nat Rev Neurosci, 2002, 3(3) : 175–190. DOI:10.1038/nrn753 |
| [7] | Wilson MA, Brunger AT. Domain flexibility in the 1.75 A resolution structure of Pb2+-calmodulin[J]. Acta Crystallogr D Biol CrystaUogr, 2003, 59(Pt 10) : 1782–1992. |
| [8] | Nihei MK, Guilarte TR. Molecular changes in glutamatergic synapses induced by Pb2+: association with deficits of LTP and spatial leaming[J]. Neurotoxicology, 2001, 22(5) : 635–643. DOI:10.1016/S0161-813X(01)00035-3 |
| [9] | 文涛, 孙黎光, 刘阳, 等. 慢性铅暴露对小鼠CaMKⅡ蛋白表达的影响[J]. 中国公共卫生, 2006, 22(10) : 1211–1212. |