


 ,
, 2. 总后第一门诊部
银河宇宙射线(GCR)中, 98%是质子和更重的离子, 它们较太阳粒子具有更高的穿透能力。在无屏蔽层条件下, 这些粒子产生的当量剂量占总当量剂量74%, 而有屏蔽的条件下, 是各种离子的相对当量剂量和由这些初级GC R粒子碎片产生的次级中子贡献的剂量。初级GCR离子的平均射程为10g /cm2, 而当一个初级离子破碎时, 会产生核电荷较小的粒子, 特别是质子和中子。随着屏蔽层厚度的增加, 总当量剂量中质子和中子所占比例增大, 没有屏蔽时占10%, 防护层厚度到30g /cm2时, 中子和质子的比例增加到61%, 其生物学效应有时能等于或甚至大于初级粒子, 所以研究中子损伤和中子相对生物效应具有重要意义[1]。染色体畸变和微核是检测辐射损伤的敏感指标, 其发生率和辐射剂量间存在密切的关系, 常用作生物剂量计估算人体吸收剂量。本研究选用35MeV质子轰击铍靶产生快中子(中子能量为18MeV), 观察染色体畸变率和微核率, 研究中子照射的剂量效应关系及其相对生物效应。
1 实验材料 1.1 标本采集血样采自2名健康男性自愿献血者, 平均年龄(25 ±1.5)岁, 无烟酒嗜好, 近期内未进行过任何医学诊断或治疗, 也未接触过有毒有害化学物质和射线照射。无菌条件下采集外周静脉血20mL, 肝素抗凝, 混匀, 分装于照射管内, 备用。
1.2 照射条件照射源:中国航天部高能物理所的快中子治疗装置, 照射位置距准直器出口处21.4cm, 束流为21-23μA, 中子束流由35 MeVp-Be质子束轰击厚铍靶产生, 固定水平束, 快中子平均能量约18MeV, 总剂量中约混有2%的γ射线, 剂量率约为18 ~ 20cGy / min。中子剂量用自行研制的医用剂量系统监测, 照射剂量由计算机控制。
分5个剂量点进行照射, 照射剂量分别为0.5、1、1.5、2和3Gy, 由高到低剂量逐一照射, 照射结束后, 将血样置37℃培养箱中静置1.5h。
60Co-γ射线照射:实验在军事医学科学院二所钴源室进行, 距源4m, 照射剂量与剂量率与中子相同。
2 实验方法 2.1 标本培养 2.1.1 染色体采用提前加秋水仙素法, 分别吸取各剂量点血样0.5 mL, 加入到5 mL混合培养液(由RPMI 1640培养基、植物血凝素、小牛血清等配制)中, 同时加入秋水仙素, 终浓度为0.015 μg /mL, 37℃恒温培养48h。
2.1.2 微核采用松胞素法, 将0.3 ml肝素抗凝血, 加入到3 mL混合培养液中, 置37℃温箱培养44h, 加入松胞素B, 使最终浓度为6mg /ml, 继续培养至72h收获细胞。
2.2 标本制备[2] 2.2.1 染色体标本制备将培养物混匀移入5ml离心管, 离心10 min, 吸弃上清液, 加预热37℃的0.075 mol /L KCl 8ml, 轻轻混匀, 置37℃温箱低渗30 min, 加固定液(3: 1甲醇和冰醋酸)0.5 ml, 轻轻混匀, 离心10 min, 吸尽上清液, 重新加固定液5ml, 第二次固定20 min。离心去上清, 加少量固定液, 调节细胞至适当浓度, 滴片在预冷湿玻片上, 空气干燥。10%Giemsa染色15 min, 空气干燥。
2.2.2 微核标本制备取出培养液1ml。加入低渗液3ml, 混匀, 立即加0.5ml固定液, 混匀。离心10 min (1 500r / min)。吸弃上清液, 加固定液5ml, 固定30 min。离心弃上清, 加适当固定液混匀, 滴片。Giemsa染色。
2.3 观察指标 2.3.1 观察染色体数目为46±1、分散良好、长度适中的中期分裂细胞, 主要记录染色体非稳定性畸变, 包括无着丝粒体、双着丝粒体和着丝粒环。一名观察者所见到的畸变必须由另一观察者进行审核, 并记录显微镜坐标。按照1个三着丝点(tri)=2个双着丝点(dic), 1个四着丝点(tet)=3个双着丝点类推, 将全部畸变换算为双着丝点+环数目(dic+r), 结果以“双+环”/细胞表示。
2.3.2 双核淋巴细胞是经过一次核分裂而胞浆未分裂的淋巴细胞。微核率是1 000个双核细胞中的微核数。微核辨认的标准:主核及微核均位于胞浆内; 微核的直径小于主核的1/3;呈圆形或近圆形; 无折光性; 着核与主核一致, 稍浅或稍深; 在较大的微核中可见到染色质结构; 主核与微核互不相连, 如有重叠或切迹, 需清晰地看到各自完整的核膜; 只分析胞浆完整的细胞。
2.4 数据处理每个剂量点分析细胞数必须满足统计学要求, 由公式n=[(1-p)×96.04] /p (p为畸变细胞率, n为分析细胞数)求得。P可以从预实验中得到, 也可以在分析过程中求出。标准误计算采用S D=√X/n。其中X为染色体畸变数(微核数), n为分析细胞数。将结果输入涂开成等[3]研制的统计分析软件, 从20个数学方程式中挑选最佳模型, 拟合标准曲线。
RBE的确定通常是以60Co-γ射线为基准。计算中子的相对生物效应(RBE)。
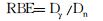
|
式中, RBE为相对生物效应; Dγ为诱发一定数量染色体畸变(或微核)所需γ射线剂量, Gy; Dn为诱发相同数量染色体畸变(或微核)所需中子剂量, Gy。
3 结果 3.1 中子和60Coγ射线诱发染色体畸变的剂量-效应关系中子和60Coγ射线照射诱发人血淋巴细胞染色体畸变结果分别见表 1和表 2。
|
|
表 1 快中子照射诱发人血淋巴细胞染色体畸变结果 |
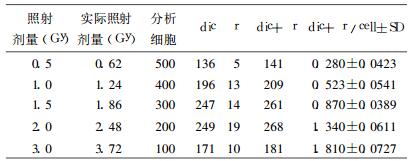
|
|
表 2 60Co-γ射线照射人淋巴细胞染色体畸变结果 |
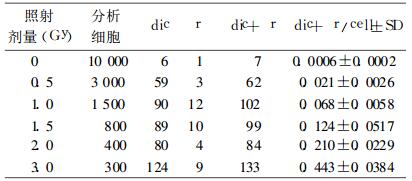
中子和60Coγ射线照射诱发人外周血淋巴细胞微核的统计结果分别见表 3和表 4。
|
|
表 3 中子照射人外周血淋巴细胞微核分析结果 |

|
|
表 5 60Co-γ射线照射人淋巴细胞微核分析结果 |
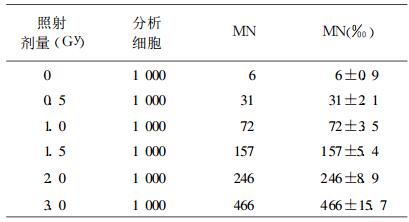
根据表 1、表 2、表 3和表 4的统计结果, 用涂开成软件拟合回归方程, 计算方程拟合度(r2), 检验回归系数显著性。用涂开成等软件进行剂量效应曲线回归方程式拟合。结果表明, 18MeV中子和60Co-γ射线在0.5 ~ 3.0Gy范围内, 染色体和微核拟合的最佳数学模式, 均为二次多项式y=c+aD+bD2, (其中y为畸变类型, D为照射剂量)。中子诱发染色体dic+r畸变, 最佳回归方程为Y=0.407D+0.0313D2、诱发淋巴细胞微核, 最佳回归方程为Y=0.060 +0.136D+0.032D2; 60Coγ射线胞诱发染色体dic+r畸变, 最佳回归方程为Y= 0.035D+0.039D2、诱发淋巴细胞微核, 最佳回归方程为Y= 0.071D+0.033D2, 各回归方程的回归系数检验, P皆小于0.01, 表明回归方程成立。结果见表 5。
|
|
表 5 中子和60Coγ诱发人外周血淋巴细胞染色体畸变和微核拟合结果 |

中子与60Coγ射线诱发人染色体畸变和微核的相对生物效应结果列于表 6和表 7。
|
|
表 6 人淋巴细胞染色体畸变(双+环)的中子RBE值 |

|
|
表 7 人淋巴细胞微核的中子RBE值 |

由表 6和表 7可以看出, 本试验中18MeV中子与60Coγ射线诱发染色体畸变dic+r的RBE值是1.77 ~ 3.59;诱发微核的RBE值是1.24 ~ 2.91。
4 讨论(1) 本研究结果表明, 在相同剂量水平, 中子诱发双加环畸变的效能大于60Coγ射线, 尤其在小剂量时效能更明显, 这主要表现在快中子诱发染色体畸变可以在同一个细胞内见到多个畸变。根据每细胞双加环的分布, 当剂量在0.5Gy, 一个细胞中可以见到3个双加环畸变, 剂量在2Gy以上时, 见到有3 ~ 4个双加环畸变的细胞很多见, 最多的一个细胞甚至见到8个双加环畸变。这种现象在60Coγ射线情况下并不多见, 除非在剂量大于4Gy时才有可能见到多畸变细胞。
(2) 关于快中子诱发染色体畸变的RBE值, 文献上多数报导的是裂变中子和单能中子[4]。类似能量的中子生物效应研究, 1977年日本Muramatsu等[5]用Be (d、n)快中子照射人血剂量为0.48 ~ 3.84Gy, 得出双着丝粒畸变和剂量为直线-平方关系, 与200kVpX射线相比较, RBE值为1.2 ~ 2.3, 随中子剂量增加而减少, 和本文中子与60Coγ射线诱发染色体畸变的RBE值1.77 ~ 3.59的结果很近似。
(3) 以微核率作观察的生物终点, 本文18MeV中子对60Coγ射线的RBE值是1.24 ~ 2.91。白玉书等[6]用2.14MeV中子对60Coγ射线分别照射离体人血, 建立微核的剂量效应曲线。中子剂量在0.1 ~ 3.0Gy之间, RBE从11.4到1.69。Vral [7]等用5.5MeV中子和60Coγ射线分别照射离体人血, 比较微核率的RBE值是从5.31到1.64。说明中子生物效应的大小, 首先决定于中子本身的能量。由于中子能量不同, 它们彼此之间的RBE值亦有差异。在不同辐射中, 都是一个相等的剂量单位时, 它们所引起的机体损害却有很大的差异[8]。
(4) 中子与60Coγ射线诱发染色体畸变拟合的方程, 都符合线性平方模型, Y=bD+cD2, 该模型的含义是双着丝粒的畸变由2个分别的剂量-反应函数引起的畸变率的总和, bD项由一个射线径迹诱发的双着丝粒; 而cD2项表示两个或更多的独立射线径迹诱发的双着丝粒。60Coγ射线拟合的染色体畸变方程中, 系数b和c两者差别不大, 说明60Coγ射线属于稀疏电离辐射, 仅仅诱发少量的双着丝粒, 而且基本上是单个径迹事件的结果。在高剂量时, 大部分双着丝粒起因于2个径迹事件, 而且它们的产额将主要随辐射剂量的平方函数变化; 中子拟合的染色体畸变方程中, 系数b是c的10几倍, 说明染色体的畸变系数b起主要作用, 也就是说中子在整个径迹中沉积大量的能量, 绝大部分染色体的畸变由单个的独立径迹产生, 也就是说高LET辐射的中子射线沿径迹沉积的能量诱发了多个染色体断裂, 剂量的平方项没有明显的贡献。
(5) 中子的RBE值随着剂量的增大而降低, 不存在恒定的RBE值。中子照射产生的双着丝粒和环状染色体与60Coγ射线比较, 当低剂量诱发的绝大部分双着丝粒起因于单个径迹事件时, 低剂量辐射最大地显示的RBE差别。Matsubara认为低剂量部分的高生物效应是由于低剂量比高剂量照射时有丝分裂延缓较少之故。低剂量时, 有丝分裂抑制现象较少, 而且微核也比高剂量时出现的早。与较低剂量比较, 较高剂量照射则有丝分裂延缓, 并导致反应的不同速率。当中子剂量大于1Gy和60Coγ射线剂量大于2Gy时, 无疑, 此效应又结合有细胞杀伤效应在内。
(6) 较敏感的效应终点所得的RBE比其他效应终点所得的RBE要大。笔者以染色体畸变率确定的中子的RBE大于以微核率来确定的RBE, 说明染色体畸变对中子比微核更为敏感, 这和已有文献[9]的结论是一致的。
(7) 微核产生于染色体畸变, 微核率在一定程度上反应了染色体畸变率。两者都和剂量相关, 微核和染色体畸变确系同类损伤所致, 只是表现形式不同。关于微核形成机制有很多学说[10], 目前的看法是染色体畸变产生的断片在细胞分裂时滞留在细胞浆中而形成微核, 本研究结果在一定程度上支持这个学说。
| [1] |
杨垂绪, 梅曼彤主编.太空放射生物学[M].广州: 中山大学出版社, 1995.
|
| [2] |
金璀珍主编.放射生物剂量估计[M].北京: 军事医学科学出版社, 2002.
|
| [3] |
涂开成, 曹珍山, 叶常青, 等. 辐射剂量-效应研究中的数学模型及应用[J]. 中国辐射卫生, 1993, 2(3): 107-110. |
| [4] |
Bauchinger M, Koestert L, Schmid E, et al. Chromosomeab-errations in hunman lymphocytes induced by fission neutrone[J]. Int.J.Radiat.Biol, 1984, 45(5): 449. |
| [5] |
Muramatsu S., Maruyama T. Chromosome aberrations in hu-man lymphocyte after irradiation with NIRS-cyclotron fast neutron in vitro[J]. A preli minary report Nipp Act Radiol,, 1977, 37(10): 995. |
| [6] |
白玉书, 黄绮龙, 关树荣, 等. 快中子对X和γ射线诱发淋巴细胞微核效应的比较研究[J]. 中华放射医学与防护杂志, 1998, 18(4): 259-262. DOI:10.3760/cma.j.issn.0254-5098.1998.04.015 |
| [7] |
Vral A, Verhaegea F, Thieretts H, et al. MicTonudd induced by fast neutrons versus 6oCoγ rays in human peripheral blond lymphocytes[J]. Int J Radiat Biol, 1994, 65: 321. DOI:10.1080/09553009414550381 |
| [8] |
王玉民. 中子辐射生物效应[J]. 辐射防护通讯, 1982, 2(11): 9-21. |
| [9] |
杜泽吉, 李延义, 刘德贵. 中子诱发小鼠的细胞遗传学效应及其相对生物效应[J]. 苏州医学院学报, 1994, 14(4): 266-270. |
| [10] |
杜峰涛, 李林. 细胞微核形成机理探讨[J]. 现代检验医学杂志, 2007, 4(22): 19-22. |