


DNA是生物体携带遗传信息的重要载体, 也是辐射效应中最重要的靶标。在电离辐射的作用下, DNA分子将产生多种类型的损伤[1], 包括碱基损伤, DNA单链断裂(single-stand break, SSB)和双链断裂(double-stands break, DSB), DNA- DNA交联以及DNA-蛋白质交联等。其中DNA链断裂是辐射引起的主要形式, 而DNA双链断裂的程度被认为与细胞的存活密切相关[2]。对于DNA的损伤, 一方面体内的修复系统积极参与消除或修复损伤, 以使遗传物质恢复到原有的状态。另一方面如果受损DNA未得到及时、有效的修复或发生错误修复, 则可能导致遗传突变和肿瘤发生。IRM-2小鼠是我所培育的近交系小鼠品系, 该小鼠突出的生物学特性是具有较强的辐射抗性[3]及较低的染色体畸变率[4]。我们通过对IRM-2小鼠及其亲本ICR/JCL和615小鼠进行对比研究, 观察电离辐射后DNA链断裂及DNA链断裂修复, 分析DNA修复系统在IRM-2小鼠辐射抗性中发挥的作用, 来探讨IRM-2小鼠辐射抗性产生的可能机理, 这将对辐射损伤及其修复机制研究提供新的理论依据。
1 材料和方法 1.1 材料 1.1.1 动物IRM-2小鼠、ICR/JCL及615小鼠, 6~8周龄, 体质量(20±2)g, 雄性, 随机分组。常规喂养, 由本所国家二级动物室培育[3], 级别为清洁实验动物。
1.1.2 主要试剂低熔点琼脂糖凝胶及脉冲场电泳级琼脂糖凝胶均为美国Bio-Rad公司产品。蛋白酶K为德国Merk公司产品。三羟甲基氨基甲烷(Tris)和乙二胺四乙酸(EDTA)为Babico公司产品。
1.1.3 主要仪器137Csγ射线照射源为加拿大产(型号USD), CHEF MAPER脉冲场电泳仪和Doc1000凝胶成像系统均为美国Bio-Rad公司产品。
1.2 方法 1.2.1 动物照射DNA链断裂测定:IRM-2、ICR/JCL及615小鼠分别经1、2、4、8Gyγ射线照射, 立即断颈椎处死小鼠, 取脾脏制备DNA样品, 对照组小鼠不照射。每个剂量点6只小鼠, 剂量率0.99Gy/min。DNA链断裂修复测定:小鼠分别接受2、4、8Gyγ射线照射, 分别于照射后0、0.25、0.5、1、2、4h断颈椎处死小鼠, 取脾脏制备DNA样品。每个时间点6只小鼠, 剂量率0.99Gy/min。
1.2.2 脾细胞DNA样品制备小鼠断颈椎处死, 迅速取出脾脏, 浸泡于冰浴的氯化钠-Tris-EDTA(STE)缓冲液(0.1mol/L NaCl, 10mmol/L Tris, 1 mmol/L EDTA, pH 7.4)中, 将降温处理的脾脏转入预冷的玻璃匀浆器内, 加入适量STE, 于冰上匀浆。取适量的匀浆液(含1×105)与等量37℃1%低熔点琼脂糖凝胶混匀, 迅速加入预冷的制胶模具中, 在4℃凝固10min。中性裂解(测定DNA双链断裂):胶块转入中性裂解液(1%月桂酰肌氨酸钠, 1mg/mL蛋白酶K, 0.25mol/L EDTA, pH7.4)中, 50℃裂解14h后Tris-EDTA(TE)缓冲液清洗胶块3次, 上样电泳。碱性裂解(测定DNA单链断裂):胶块转入碱性裂解液(0.5%月桂酰肌氨酸钠, 1.2mol/L NaCl, 30mmol/L NaOH, 2mmol/L EDTA)中, 室温裂解1h, 用碱性漂洗液(30mmol/L NaOH, 2mmol/L EDTA)浸泡1h, 中间换液1次, 清洗完毕, 上样电泳。
1.2.3 脉冲场电泳用1×TAE(40mmol/L Tris, 20mmol/L NaAc, 10mmol/L EDTA)制备0.75%琼脂糖凝胶, 将制备好的含DNA的胶块填入加样孔中, 用少量低熔点琼脂糖凝胶封固。中性电泳条件:电泳槽中加入1×TAE缓冲液, 电压梯度1.0 V/cm, 角度120°, 转换时间60s, 14℃恒温电泳20h。碱性电泳条件:电泳槽中加入碱性漂洗液, 电压梯度0.6V/cm, 角度120°, 转换时间10s, 14℃恒温电泳20h。电泳结束后, 将凝胶转入含0.5mg/L的溴化乙锭(EB)染液中染色12h, 然后转入蒸馏水中脱色4h。
1.2.4 电泳图像分析将染色后的凝胶转入Doc1000凝胶成像系统采集图像, 用Molecular AnalysisTM软件分析DNA条带分布及灰度关系。
1.2.4.1DNA链断裂量以孔外进入凝胶(样品胶块释放)的DNA占孔外和孔内总DNA比例(FAR)表示, 具体按以下公式计算[5]:
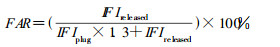
|
式中:IFI(Integrated fluorescent intensities)代表DNA条带的荧光强度积分值, IFIreleased为进入凝胶的DNA条带的荧光强度积分值, IFIplug为加样孔内DNA的荧光强度积分值。
1.2.4.2DNA链断裂修复以剩余损伤占初始损伤的百分比表示, 即Percent of Breaks Remaining, 按下述公式计算:
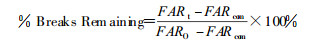
|
式中, FARt为不同修复时间后DNA断裂量, FAR0为照射后0时间DNA初始损伤量, FARCon为对照组(未照射组)DNA断裂量。
以照射后0时间脾细胞DNA断裂(FAR)为100%, 经不同时间修复后DNA断裂占0时间DNA断裂百分比表示损伤修复程度。
1.3 统计学处理实验数据以x±s表示, 计量资料采用t-检验。以P < 0.05为有统计学意义。
2 结果 2.1 小鼠脾细胞DNA链的断裂 2.1.1 DNA双链断裂和单链断裂的脉冲场电泳图像不同剂量的γ射线作用于小鼠后, 照射组加样孔外可见清晰的DNA拖尾, 说明辐射造成了脾细胞DNA链断裂损伤效应。泳出的DNA片断(即dsb、ssb数量)随照射剂量的增加而增加, 而对照组加样孔外基本无DNA拖尾, 见图 1, 2。

|
图 1 小鼠脾细胞双链断裂PFGE电泳图谱 |

|
图 2 小鼠脾细胞单链断裂PFGE电泳图谱 |
对照组IRM -2小鼠本底DNA损伤较低, 即ssb数目低于未照射的亲本ICR和615小鼠, dsb的数量明显低于ICR和615小鼠(P < 0.01)。
不同剂量(1、2、4和8Gy)照射后, IRM-2小鼠ssb和dsb的数量均低于相同剂量照射的亲本ICR和615小鼠:在dsb各剂量组中, IRM-2小鼠dsb数量明显低于ICR和615小鼠(P < 0.05和P < 0.01), 见表 1; 在1和8Gyssb剂量组中, IRM- 2小鼠ssb数量明显低于615小鼠(P < 0.01和P < 0.05), 见表 2。
|
|
表 1 不同剂量照射后小鼠脾细胞DNA双链断裂 |
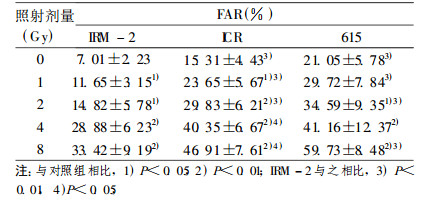
|
|
表 2 不同剂量照射后小鼠脾细胞DNA单链断裂 |
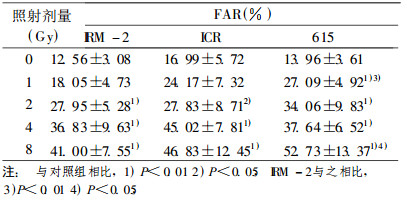
不同剂量照射后, 随修复时间的延长, 小鼠脾细胞dsb和ssb数量均呈下降趋势, 体现了小鼠抵抗外界损伤的自我保护能力。起初ssb的修复为快速修复, 0.5h后损伤基本得到修复; 而dsb的修复慢于ssb, 1h后大部分损伤才得到修复。照射后1hssb和dsb修复速度都降低, 当修复时间超过2h后, 断裂又呈现上升趋势, 见图 3, 4。

|
图 3 小鼠脾细胞双链断裂修复PFGE电泳图谱 |

|
图 4 小鼠脾细胞单链断裂修复PFGE电泳图谱 |
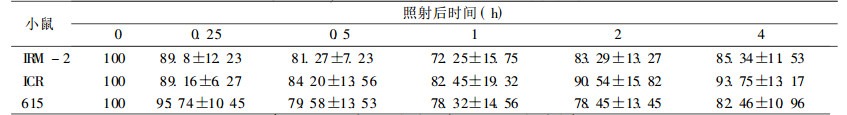
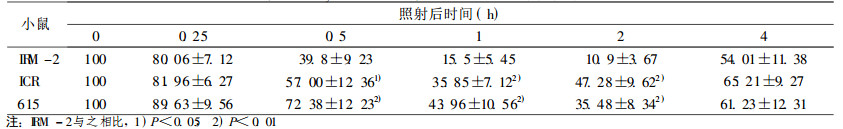
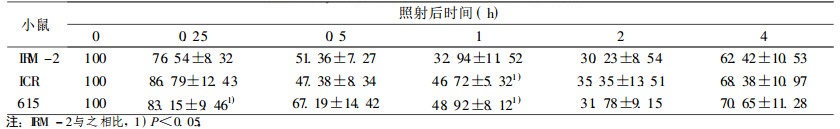
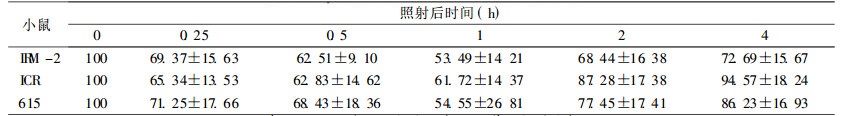
在较低剂量2 Gy时, IRM-2小鼠约20% dsb和50% ssb得到修复, 与亲本615及ICR/JCL小鼠相比, 无统计学差异。当接受4、8Gy剂量照射后, IRM-2小鼠表现出较高的修复效率, 表现在:4Gy 0.5h后约60% dsb损伤得到修复, 2h后基本修复完全达90%, ssb修复较快于dsb, 1h后剩余的损伤已接近本底水平; 8Gy大剂量照射后1h约70% dsb损伤修复, ssb修复较快, 1h后修复90%。而亲本小鼠修复后剩余的损伤远高于IRM-2小鼠, dsb和ssb修复速率也比IRM-2小鼠慢(P < 0.05和P < 0.01), 见表 3~8。
|
|
表 3 2Gy照射后小鼠脾细胞DNA双链断裂修复 |
|
|
表 4 4Gy照射后小鼠脾细胞DNA双链断裂修复 |
|
|
表 5 8Gy照射后小鼠脾细胞DNA双链断裂修复 |
|
|
表 6 2Gy照射后小鼠脾细胞DNA单链断裂修复 |
|
|
表 7 4Gy照射后小鼠脾细胞DNA单链断裂修复 |
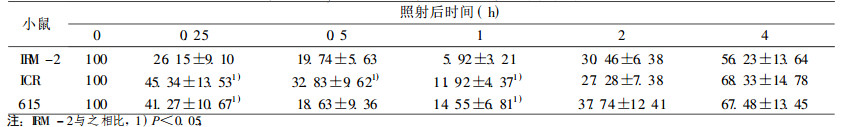
|
|
表 8 8Gy照射后小鼠脾细胞DNA单链断裂修复 |
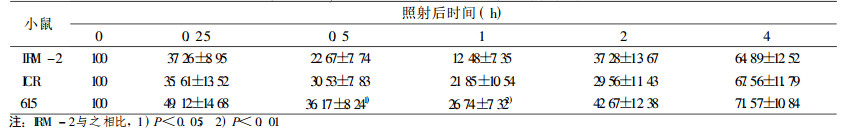
早在1968年, Alexander提出了细胞辐射敏感性取决于其DNA链断裂修复能力的概念。近年来, 越来越多的研究证明, DNA链断裂和修复与辐射敏感性密切相关, DNA单链断裂和断裂后的修复与细胞辐射敏感性有一定的相关性, 更多的研究认为DNA双链断裂重接能力是细胞辐射敏感性的一个决定因素[6]。
在离体细胞研究中已有大量资料说明DNA双链断裂修复水平低下与高敏感性的一致性。Zhou等用脉冲场电泳法对18例脑肿瘤病人的肿瘤细胞测定其离体20Gy照射后的DNA双链断裂修复, 按修复保温2h末的残留DNA双链断裂量判断, 观察到来自不同病人的肿瘤细胞的DNA修复水平确有不同。发现修复水平低的放疗效果较好, 存活时间长, 而修复水平高的对放疗均表现抗性。
我们用脉冲场电泳法研究IRM-2小鼠辐射抗性的机理时, 发现辐射诱发的DNA链断裂中IRM-2小鼠DNAdsb和ssb数量明显低于ICR/JCL及615非抗性小鼠。在DNA链断裂修复中, IRM-2小鼠不仅修复后剩余的损伤远低于非抗性亲本小鼠, 而且dsb和ssb修复速率比亲本小鼠快, 表现出更高的修复效率, 体现了IRM-2小鼠辐射抗性的特性。本研究也印证了DNA链断裂和修复与辐射敏感性密切相关。
大部分的体外实验显示, 并非所有的dsb都能得到修复, 本实验也发现如此。目前人们还不知道为什么有些断裂能够修复, 而有些断裂则不能。另外由于dsb造成DNA双链分子的完全离断, 所测到的修复并不表示重接是否正确, 这也使问题分析更加复杂。
令我们感到意外的是, 小鼠修复后2h DNA链断裂又呈现上升趋势, 可能是由于电离辐射诱发小鼠脾细胞凋亡, 细胞DNA降解从而产生新的片断, 这与先前的研究结果相符[7]。
| [1] |
Jenner TJ, Fulford J, O, Neill P. Contribution of base lesions to radiation - induced clustered DNA damage:implication for models of radiation response[J]. Radiat Res, 2001, 156(5Pt2): 590-593. |
| [2] |
Prise KM, Pinto M, Newman HC, et al. A review of studies of ionizing radiation - induced double - strand break clustering[J]. Radiat Res, 2001, 156: 572-576. DOI:10.1667/0033-7587(2001)156[0572:AROSOI]2.0.CO;2 |
| [3] |
周继文, 吴红英, 王月英, 等. IRM-2近交系小鼠对电离辐射抗性的研究[J]. 中国实验动物学报, 2001, 9(2): 73-77. DOI:10.3969/j.issn.1005-4847.2001.02.002 |
| [4] |
李进, 周继文, 吴红英, 等. IRM-2近交系小鼠的G-显带核型和自发畸变率的研究[J]. 中国实验动物学报, 2001, 9(3): 178-181. DOI:10.3969/j.issn.1005-4847.2001.03.013 |
| [5] |
Gulston M, de Lara C, Jenner T, et al. Processing of clustered DNA damage generates additional double - strand breaks in mammalian cells post - irradiation[J]. Nucleic Acids Res, 2004, 32(4): 1602-1609. DOI:10.1093/nar/gkh306 |
| [6] |
Maity A, Kao GD, Muschel RJ, et al. Potential molecular targets for manipulating the radiation response[J]. Int J Radiation Oncol Biol Phys, 1997, 37: 639-653. DOI:10.1016/S0360-3016(96)00598-6 |
| [7] |
李雨, 郑秀龙, 罗成基, 等. 恒场电泳检测DNA双链断裂及其应用[J]. 辐射研究与辐射工艺学报, 1998, 16(4): 234-237. |