


肿瘤的诱导分化疗法和凋亡疗法被誉为近二十年来肿瘤疗法上的两次较大的突破, 它突破了传统的杀伤疗法(手术、放疗和化疗等)的思想, 诱导分化疗法使肿瘤细胞"改邪归正"为正常细胞, 凋亡疗法则通过加快肿瘤细胞的凋亡, 从而调整肿瘤细胞所特有的分化受阻和增殖失控, 达到治疗肿瘤的目的。在上述两种疗法的分子水平和临床研究上, 我国学者陈竺院士和王振义院士的工作居世界领先水平, 引起国际生命科学界的重大关注。
细胞凋亡(凋零)是生命科学研究的热点问题, 作为一种不同于细胞坏死的细胞死亡方式, 它具有特征的形态和生化改变, 是细胞自身主动死亡过程, 又称为程序化细胞死亡(programmed cell death, PCD), 已有研究表明, 细胞凋亡参与正常的生理过程和病理过程。利用细胞凋亡的生化和细胞调节机制探索AIDS、恶性肿瘤和血液病治疗的新途径引起了广泛关注。其中利用中国传统医学中的砒霜(As2O3)等剧毒中药成功地治疗人早幼粒细胞白血病的临床和分子生物研究, 是该领域研究的一项突破性成果, 给出了传统医学中"以毒攻毒疗法"的分子机理, 表明了选择性诱导肿瘤细胞凋亡是肿瘤治疗上一条新的途径。
诱导分化疗法系指肿瘤细胞在诱导分化剂作用下, 使肿瘤细胞自发地分化为良性或正常细胞。二甲基亚砜(DMSO)是发现最早的肿瘤细胞诱导分化剂(1971年), 作为不同于传统肿瘤杀伤治疗的思路, 国内外学者已对肿瘤细胞诱导分化剂进行了广泛而深入的研究, 其中全反式维甲酸类化合物成功用于人早幼粒细胞白血病治疗的临床实践, 宣告了诱导分化疗法作为一种新的肿瘤治疗策略和方法的诞生。
尽管分化剂和凋亡剂临床和分子机理已开展大量研究, 但有关肿瘤细胞诱导分化和凋亡的量子机理方面的研究至今未见报道。
1 肿瘤细胞诱导分化和凋亡的适度氧化——抗氧化失衡机理大量研究表明, 自由基引起的氧化应激(Oxidation stress)与细胞分化和凋亡有较好的相关性。氧化应激能介导细胞分化和凋亡的主要证据有:①加入活性氧中间体(ROS)或耗竭抗氧化剂能导致细胞的分化或凋亡; ②细胞的分化或凋亡与ROS的生成有关; ③细胞分化或细胞凋亡能被抗氧化剂所阻断。即诱导分化剂和凋亡剂的氧化——抗氧化的适度失衡, 引起的氧化应激可介导细胞的分化或凋亡。系统地分析已有细胞诱导分化剂和凋亡剂, 发现凋亡剂的自由基氧化性与细胞凋亡呈正相关, 分化剂的自由基抗氧化性与细胞分化呈正相关。基于大量文献研究结果和作者提出的自由基和抗氧化系统理论, 本研究提出了细胞诱导分化剂和凋亡剂的氧化——抗氧化失衡的量子机理假说:
(1) 诱导分化剂或凋亡剂为得失单电子均衡性较好的物质(自由基或抗氧化剂)。
(2) 具有适中氧化性和抗氧化性物质的双向适度失衡可诱导细胞分化或凋亡。细胞分化与物质抗氧化性呈正相关, 细胞凋亡与物质氧化性呈正相关。分化剂的抗氧化性略强于氧化性, 凋亡剂的氧化性略强于抗氧化性。
(3) 细胞诱导分化机理可能与细胞得单电子过程相关, 细胞凋亡机理可能与细胞失单电子过程相关。
为验证上述细胞分化或凋亡的量子机理假说, 同时为设计分化剂和凋亡剂提供理论依据和判据。本研究用非经验的量子生物学方法, 研究了分化剂和凋亡剂的代表物DMSO、As2O3及其形成的正、负离子型自由基的结构和性能, 研究结果支持肿瘤细胞诱导分化或凋亡的量子机理。
2 实验方法本研究采用与理论和实验相平行的第三种研究方法数值计算技术。采用理论上严格的量子生物学从头算方法(abinitio)和密度泛函方法(DFT)。全部研究工作采用国际上著名的Gaussian-94软件在PC机上完成。
闭壳层分子和开壳层分子(自由基)分别采用RHF和UHF方法, 为便于统一比较, 计算统一采用ST0 -3G*基组, 全优化分子的几何构型, 并对优化分子结构做振动分析, 确定优化构型的合理性(不能有虚红外振动频率), 并获得可与实验值相对照的IR和Raman光谱值。为简化计算, 晶体As2O3结构用其溶于水中形成的AsO3模型代替。
3 实验结果和讨论DMSO、As2O3分别计算其中性分子和失、得一个电子变成正、负离子型自由基的结构和性质。
3.1 诱导分化剂DMSO的结构、性质和诱导分化机理计算结果表明:诱导分化剂DMSO的骨架基团(O, S, C, C)呈三角锥构形。失去一个电子变成的阳离子自由基· DMSO+骨架分子也呈三角锥形, 其能量、零点振动能和偶极矩变化不是很大, 但得到一个电子变成阴离子型自由基· DMSO-的骨架分子构型发生巨大变化, 由原来的三角锥构型变成了平面构型, 键长有较大变化, S-O键伸长1.0nm, S-C键伸长2.0nm, 同时引起分子零点振动能较大降低46.28 kJ/mol, 分子能量略有升高, 且上升幅度大于阳离子自由基。一个特别值得注意的现象是阴离子型自由基的偶极矩由中性分子的强极性(μ=2.90D)变为非极性(μ=0.03D)。从热力学角度看, DMSO易失去一个电子变成有较高稳定性的阳离子型自由基, 这与DMSO曾是著名的自由基捕捉剂和辐射防护剂是一致的。DMSO阴离子型自由基与阳离子自由基相比, 虽热力学稳定性不占优势, 但从运输动力学角度看, 由中性分子变成阴离子型自由基分子的极性实现双向转化, 使DMSO在生物体系中易于穿透象细胞膜这种由极性(亲水)和非极性(亲脂)组成的两性体系, 这与DMSO是优良的溶剂和膜助渗剂是一致的。同时也给出了DMSO用于研究蛋白质折叠、解链性质和溶解胆结石的溶剂性能的结构基础。
自旋密度分布是自由基化学和生物学普遍关注的问题, 实验数据可由ESR实验的超精细分裂常数间接提供。从自由基自旋密度分布看, DMSO正、负离子型自由基的分布大致相似, 均以O原子上的自旋密度最大。这与传统观念中DMSO的辐射防护性能是含硫自由基的观点是矛盾的。本研究结果确定DMSO得失电子形成的正、负离子自由基为含氧自由基, 硫元素只起增加氧自由基稳定性的作用。这一结果, 修正辐射防护剂DMSO作为硫自由基的传统观点。
从前线轨道看, DMSO的HOMO和LUMO轨道能量之比, 可定义为前线轨道能级指数Z:

|
Z值在最优化均衡的黄金分割率Z0=0.618附近, Z>0.618抗氧性略高于氧化性, 支持本研究提出分化剂的双向均衡性要求。
综合分析上述结果, 可以认为DMSO具有典型的双向自由基功能, 其失电子变成阳离子型自由基热力学占优势; 得电子形成阴离子型自由基的运输动力学性能占优势。阴离子型自由基分子构型(构象)和极性的双向转化, 可能与其诱导分化作用有一定联系, 这与文献中提出的DMSO诱导分化可能与改变细胞膜的迁移性能和稳定性有关是相符的, 本研究结果给出了其量子水平微观结构基础。同时也说明细胞诱导分化过程可能与细胞失去单电子相关, 实验结果支持本研究中提出的诱导分化剂的量子机理。
3.2 诱导凋亡剂As2O3的结构、性质和诱导凋亡机理As2O3的全部研究结果表明:As2O3及失去电子形成的阳离子自由基均有C3v对称性, 构型变化不大:阳离子型自由基的As-O键长缩短1.0nm, 键角减小3.3nm, 阳离子型自由基稳定性升高, 零点振动动能增大。由于尚未优化到得电子的阴离子型自由基的构型, 可能与其有较强氧化性相关。从热力学角度看, 可能阴离子型自由基占优势。
自由基的自旋密度分布表明, 阴离子型自由基主要分布O和As原子上但作进一步Fermi分析确定, 自旋电子主要在As原子上。即As2O33-的自由基为砷自由基。
As2O3的前线轨道能级指数计算表明:
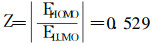
|
Z值在黄金分割率Z0附近, 支持本研究中提出的双向性物质的要求, Z0值略小于0.618, 故As2O3的氧化性能要略强于其抗氧化性。这点也支持本研究中提出的细胞凋亡的量子机理。双向均衡性Z值可能成为筛选细胞凋亡剂简单的初级判据。
总之, DMSO和As2O3电子水平的研究结果, 初步支持了本研究中提出的具有双向自由基功能物质的适度氧化应激可选择性诱导细胞分化和凋亡的量子机理。
4 结论本研究通过自由基生物学理论、量子生物学从头算和密度泛函理论, 获得下述几点创新性的结果:
(1) 提出具有双向自由基功能物质的适度氧化——抗氧化失衡诱导细胞分化和凋亡的量子机理。
(2) 提出的前线轨道能级指数Z可能是具有普遍意义双向性黄金分割判据, Z>0.618为细胞诱导分化剂; Z<0.618为细胞凋亡剂。可望成为筛选细胞诱导分化剂和凋亡剂生物活性的简单判据。还有待进一步研究证实。
(3) 首次从电子水平上给出了DMSO作为优良溶剂、助渗剂、辐射防护剂和细胞诱导分化剂的结构基础, 修正DMSO作为辐射防护剂的含硫自由基学说的传统观点。
(4) 发现DMSO阴离子自由基构型较中性分子有较大变化, DMSO具有双向的亲油亲水功能, 本质上起到分子的电子开关的作用, 提示在诱导分化机理上起独特的作用。
(5) 给出了DMSO、As2O3和及其正、负离子型自由基的几何构型、电子能谱、光谱、布局分析、自旋分析等微观结构判据。
| [1] |
陈竺, 陈赛娟. 维甲酸诱导早幼粒白血病细胞分化的分子机制研究[J]. 中国科学基金, 1994, 106. |
| [2] |
尉志武, 奎因. 生命科学研究中的二甲亚砜[J]. 科学, 1996, 48(1): 34-37. |
| [3] |
田晓华, 顾景范, 孙存普, 等. 氧化应激与细胞凋亡[J]. 军事医学科学院院刊, 1996, 20(4): 302-305. |
| [4] |
孙华斌, 邢桂平.肿瘤细胞诱导分化剂As2O3和凋亡剂DMSO的电子结构、光谱和作用机理研究[A].1997全国自由基与营养和医学学术研讨会论文集[C].1997.10: 7
|
| [5] |
Itoh N, Tsujimoto Y, Nagata S, et al. Effect of bcl-2 on Fas antigen mediated cell death[J]. J Immunol, 1993, 151: 621-627. |
| [6] |
Forrest V J. Oxidative stress-induced apoptos is prevented by Trolox[J]. Free Radical Biology and Medicine, 1994, 16(5): 675. |
| [7] |
Catchpoole D R. Etoposide-induced cytotoxicity in two human T-cell leukemic lines:delayed loss of membrane perme ability rather than DNA fragmentation as an indicator of programmed cell death[J]. cancer research, 1993, 53(18): 4287-4296. |