


Chemistry and biology of marine-derived Trichoderma metabolites

Abstract
Marine-derived fungi of the genus Trichoderma have been surveyed for pharmaceuticals and agrochemicals since 1993, with various new secondary metabolites being characterized from the strains of marine animal, plant, sediment, and water origin. Chemical structures and biological activities of these metabolites are comprehensively reviewed herein up to the end of 2022 (covering 30 years). More than 70 strains that belong to at least 18 known Trichoderma species have been chemically investigated during this period. As a result, 445 new metabolites, including terpenes, steroids, polyketides, peptides, alkaloids, and others, have been identified, with over a half possessing antimicroalgal, zooplankton-toxic, antibacterial, antifungal, cytotoxic, anti-inflammatory, and other activities. The research is highlighted by the molecular diversity and antimicroalgal potency of terpenes and steroids. In addition, metabolic relevance along with co-culture induction in the production of new compounds is also concluded. Trichoderma strains of marine origin can transform and degrade heterogeneous molecules, but these functions need further exploration.Graphical Abstract
Keywords
Trichoderma Metabolite Terpene Polyketide Peptide Bioactivity1 Introduction
Trichoderma was first described as a genus of filamentous fungi by Persoon in 1794 [1]. It belongs to the family Hypocreaceae of the order Hypocreales in the class Sordariomycetes of the phylum Ascomycota, rather than the previous deuteromycetous taxa, according to the Dictionary of the Fungi (10th edition) [2]. Fungal species of this genus feature numerous conidia in varying shades of green, rapid growth rate, and high adaptability to various terrestrial and aquatic ecosystems. They are free-living or occur as mycoparasites and opportunistic, avirulent plant symbionts [3]. Their identification is difficult due to the few morphological characteristics available and the slight variations among different species [4]. Most of them can be linked to the teleomorphic Hypocrea species (Ascomycota), but this generic name has not been suggested to be further used by the proposal from Rossman et al. [5]. Several species were once assigned to other genera, such as Aphysiostroma, Eidamia, Cordyceps, Gliocladium, Podostroma, Sarawakus, Sphaeria, Sporotrichum, Stilbella, Stilbum, and Thuemenella, and 65 names inclusive of the basionyms of accepted species in Trichoderma and other genera are not currently adopted [6]. Totally, 254 species and two varieties with at least 1100 strains were characterized until 2015 [6, 7], and new species have been continually recognized thereafter [8–11].
Since the initial work by Weindling regarding the parasitism of T. lignorum on other fungi in the early 1930s [12, 13], Trichoderma species have achieved great successes to suppress plant diseases and promote plant growth in agriculture [7], and more than 250 Trichoderma-based products have been commercialized so far [14]. Their interactions with pathogens and plants appear complicated, but the production of bioactive secondary metabolites is regarded as one of the key mechanisms [15–17]. Investigations toward the antagonism of Trichoderma species against phytopathogenic fungi have resulted in the isolation and identification of a number of antifungal antibiotics, including terpenes, polyketides, peptides, and alkaloids [18]. A series of metabolites have been reported as plant growth promoters or inhibitors [19], and a plethora of metabolites with other activities, such as antibacterial, cytotoxic, and enzyme-inhibitory properties, have also been discovered [19–23]. Over the past eight decades, some a thousand new polar and nonpolar metabolites have been isolated and identified from Trichoderma species of various origin including terrestrial and marine environments.
Since 1993, secondary metabolites from marine-derived Trichoderma have gradually been surveyed [24]. Until the end of 2022, 445 new compounds have been identified from 77 strains of Trichoderma fungi, 60 of which belong to 18 known species involving T. asperelloides (1 strain), T. asperellum (5), T. atroviride (9), T. aureoviride (1), T. brevicompactum (2), T. citrinoviride (4), T. erinaceum (1), T. hamatum (1), T. harzianum (12), T. koningii (3), T. longibrachiatum (8), T. orientale (1), T. reesei (4), T. saturnisporum (1), T. virens (4), T. viride (1), H. lixii (1), and H. vinosa (1) (Figs. 1, 2). These 77 Trichoderma strains have been isolated from animal (sponge (19 strains), mussel (4), fish (1), tunicate (2), sea star (2), sea fan (1), and soft coral (2)), plant (alga (18), mangrove (4), and halophyte (1)), sediment (20), and seawater (3) samples (Fig. 3). The new compounds (Tables 1, 2, 3) can be divided into terpenes, steroids, polyketides, peptides, alkaloids, and others (Fig. 4), of which 235 members possess antimicroalgal, zooplankton-toxic, antibacterial, antifungal, cytotoxic, anti-inflammatory, antiviral, phytotoxic, insect-toxic, zebrafish-toxic, antifouling, antioxidant, enzyme-inhibitory, NF-κB-inhibitory, anti-pulmonary fibrosis, anti-Aβ fibrillization, and neuroprotective activities (Fig. 5) [24–130]. The present review attempts to give a comprehensive compilation of these molecules and highlights their chemical diversity and biological activity.
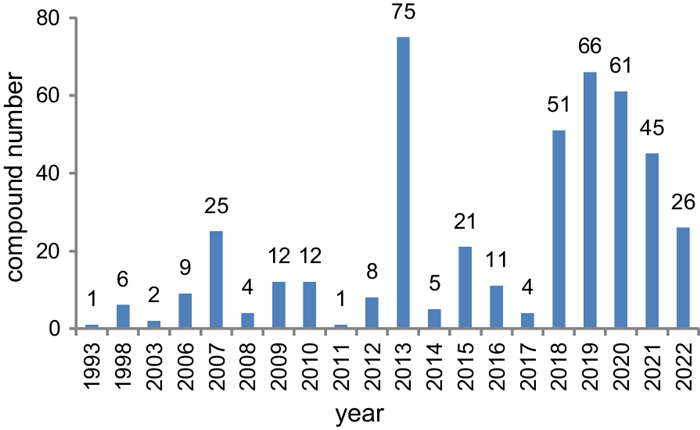
Numbers of new compounds from marine-derived Trichoderma during 1993–2022
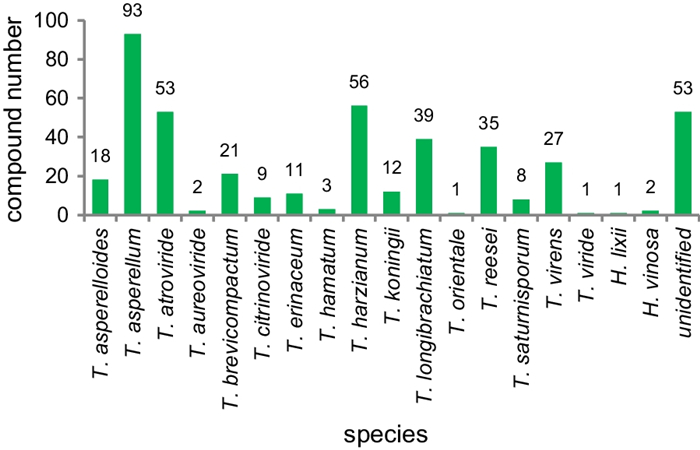
Numbers of new compounds from different marine-derived Trichoderma species
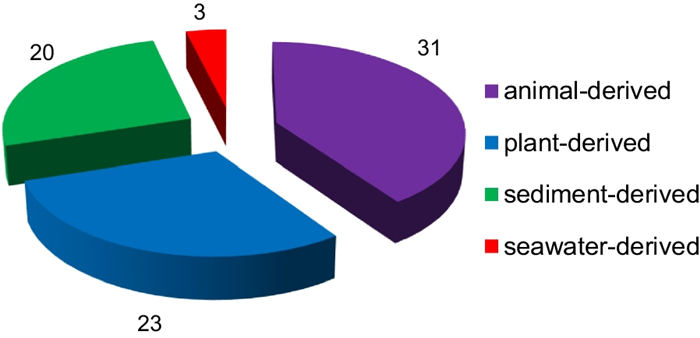
Sources and numbers of marine-derived Trichoderma strains possessing new compounds
Terpenes (1–165) and steroids (166–168) from the marine-derived Trichoderma
Polyketides (169–295) from the marine-derived Trichoderma
Peptides (296–426), alkaloids (427–440), and others (441–445) from the marine-derived Trichoderma
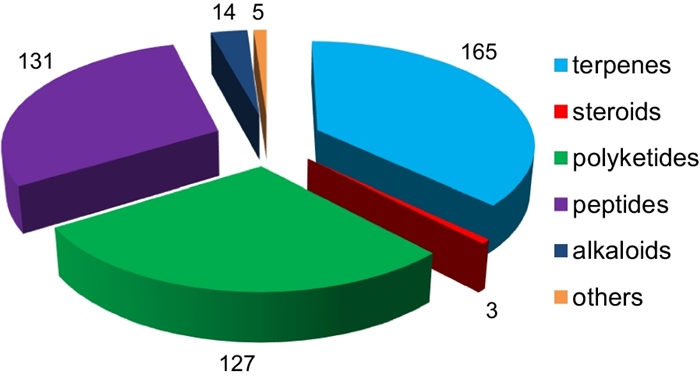
Types and numbers of new compounds from marine-derived Trichoderma
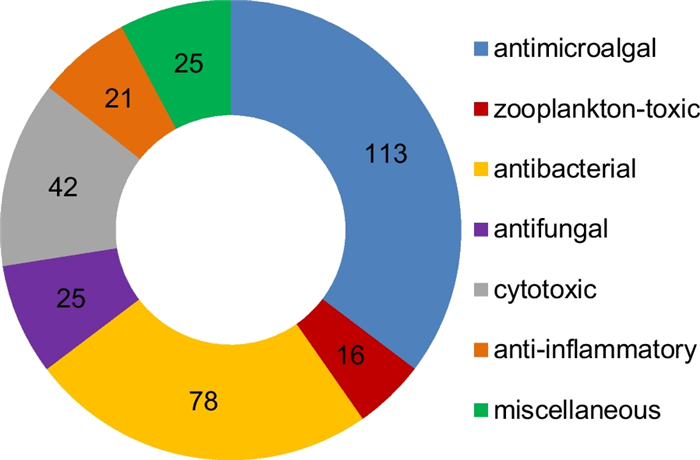
Numbers of bioactive new compounds from marine-derived Trichoderma
2 Structure and occurrence
2.1 Terpenes
A total of 165 new terpenes (1–165, Table 1) were isolated and identified from 10 marine-derived Trichoderma species, including T. asperelloides (18 compounds), T. asperellum (47), T. atroviride (2), T. brevicompactum (20), T. citrinoviride (4), T. erinaceum (5), T. hamatum (3), T. harzianum (27), T. longibrachiatum (4), and T. virens (19), and five unidentified strains (16) [25–65]. These terpenes can be classified to monoterpenes with a menthane skeleton, sesquiterpenes with cyclonerane, bisabolane, trichothecane, carotane, cadinane, acorane, cuparane, farnesane, synderane, pupukeanane, and harzianoic acid skeletons, and diterpenes with harziane, proharziane, wickerane, citrinovirin, fusicoccane, cleistanthane, and sordaricin skeletons. The above 19 basic scaffolds along with the degraded and substituted ones demonstrate the high structural diversity of terpenes from marine-derived Trichoderma.
Monoterpenes have seldom been discovered from Trichoderma species, including marine-derived ones. Monoterpenes 1 and 2 (Fig. 6) were obtained from the alga-endophytic T. asperellum and represent the only two menthane derivatives from this genus [25]. These two compounds with only two chiral centers were identified as mutual epimers by the similar but different NMR data, because those of enantiomers are the same as each other. Their absolute configurations at C-7 were assigned to be S and R, respectively, by quantum chemical calculations of electronic circular dichroism (ECD) spectra. Based on the epimeric relationship between 1 and 2, these two metabolites were proposed to have the same absolute configuration at C-1, but it was not determined spectroscopically. As a chlorinated analog of them, 3-chloromenthan-1,2,7-triol had also been isolated from a fungus, Tryblidiopycnis sp. of mangrove origin [131]. Based on previous reports, monoterpenes were rarely encountered not only in Trichoderma species but also in other marine and even terrestrial-derived fungi [132].
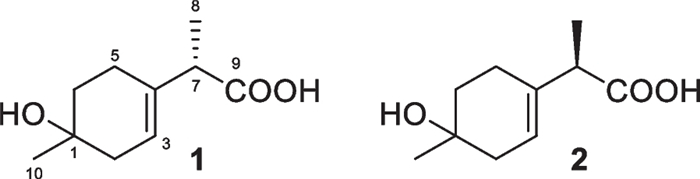
Monoterpenes from marine-derived Trichoderma
Sesquiterpenes with 126 new members (3–128, Table 1) make up the largest group of terpenes from marine-derived Trichoderma, and they comprise 11 basic carbon skeletons [26–52]. A common cyclonerane skeleton is present in 23 isolates (3–25, Fig. 7), obtained from T. asperellum [26–28], T. harzianum [29, 30], T. hamatum [31], T. erinaceum [32], T. citrinoviride [33], and T. asperelloides [34]. This scaffold is characterized by the dimethylated cyclopentane ring attached by a 1,6-dimthylhexanyl side chain. Oxidation, reduction, cyclization, and substitution construct their diverse structures. All of them feature an oxygen atom bonded to C-7. The hydroxy group at C-3 of 22–24 possesses an opposite orientation, and the double bond in the five-membered ring of 25 also renders this molecule unique [33, 34]. Two isolates (26 and 27, Fig. 7) from T. asperellum and T. asperelloides, respectively, have degraded cyclonerane frameworks, with the degradation happening at the side chain or the ring unit [28, 34]. In addition, 10 nitrogenous cyclonerane derivatives (28–37, Fig. 8) were obtained from T. asperellum [27, 35]. Each of them contains a nitrogen-bearing substitute, and the highlight is the presence of an isoxazole ring in 35 [27]. Cyclonerins A (28) and B (29) harbor a hydroxamic acid unit, that can chelate ferric ion, and represent the first two fungal hydroxamic acids with a terpene-derived scaffold [27]. The identification of these compounds was performed through various spectroscopic methods. Quantum chemical calculations were used to aid the assignments of relative configurations for 22 and 26 and absolute configurations for 22, 28–32, and 34, and X-ray diffraction was employed to determine the absolute configuration of 6 [27, 28, 33]. Although the relative configuration at C-7 of 7–9 and 12 was not given in literature [28], it should be the same as that 26 based on biogenetic considerations. Cyclonerane sesquiterpenes, especially the known cyclonerodiol, can be produced by many species of Trichoderma and other fungal genera, such as Trichothesium, Fusarium, and Epichloe [33], but they have rarely been detected in plants and animals. It is worth mentioning that marine-derived Trichoderma strains have contributed more diverse cycloneranes than terrestrial-derived ones so far [20, 22].
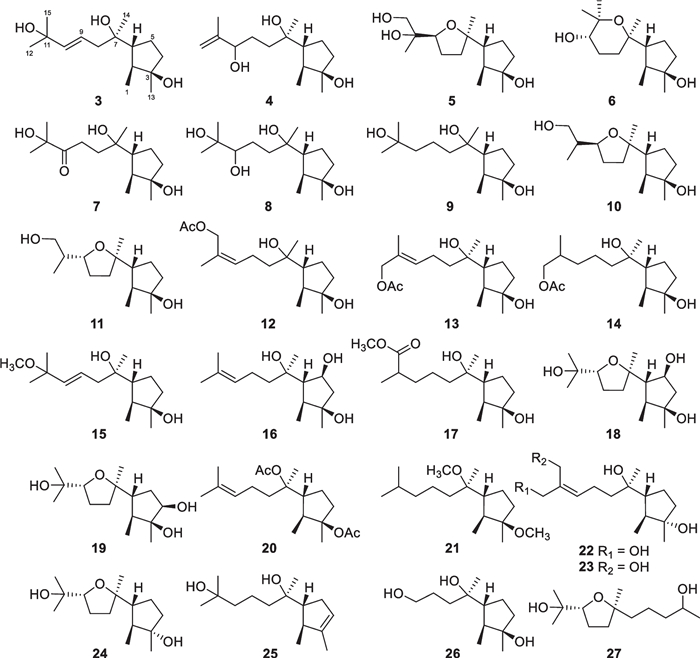
Cyclonerane sesquiterpenes and catabolites from marine-derived Trichoderma
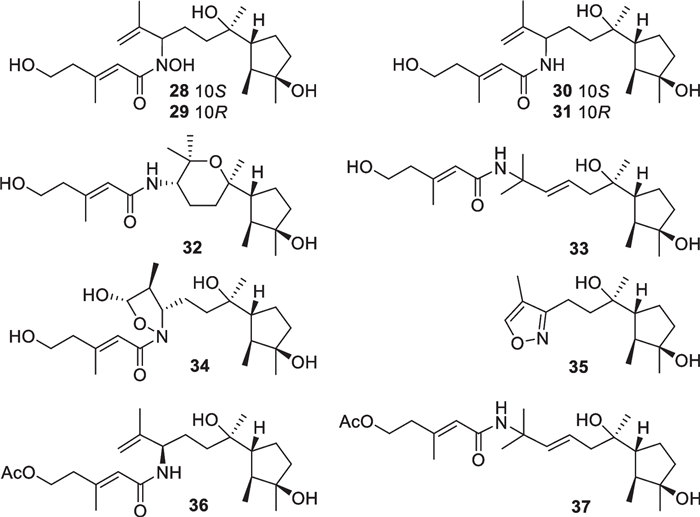
Nitrogenous cyclonerane sesquiterpenes from marine-derived Trichoderm
Bisabolane sesquiterpenes have been known as metabolites of plants, animals, and microbes for a long time, but the first discovery from Trichoderma just happened in 2011 [133, 134]. Bisabolane derivatives from marine-derived Trichoderma also exhibit high structural diversity. 14 members (38–51, Fig. 9) with an untouched bisabolane skeleton were isolated from four Trichoderma species, including T. asperellum [25, 26, 36], T. brevicompactum [37], T. asperelloides [34], and T. erinaceum [32]. Among them, trichaspside A (38) represents the first natural bisabolane aminoglycoside, and all the others possess oxygenated side chain termini. Meanwhile, 21 norbisabolane sesquiterpenes (52–72, Fig. 10), with four (53–56) containing an aminoglycoside moiety, were discovered from T. asperellum [25, 26, 36, 38], T. atroviride [39], and T. asperelloides [34]. All the norbisabolanes are possibly produced by elimination of a terminal methyl group from the side chain moieties of their precursors, and the majority of them bear an oxygen atom at C-11. It is regretted that absolute configurations for most of the bisabolane and norbisabolane derivatives remain unsolved because of lacking ECD signals and perfect crystals. However, trichobisabolin Z (72) has a Cotton effect at 328 nm due to the presence of an α, β-unsaturated carbonyl group, which enabled the assignment of the absolute configuration at C-6 by quantum chemical calculations [34]. An acidic hydrolysis reaction was performed during the absolute configuration establishment of 38 [26]. Trichodermaerin A (51) with no optical activity was deduced to be a racemic mixture, but the separation failed via various chiral HPLC columns [32]. In general, the high populations of norbisabolane sesquiterpenes and aminoglycosides may be characteristic of marine-derived Trichoderma.
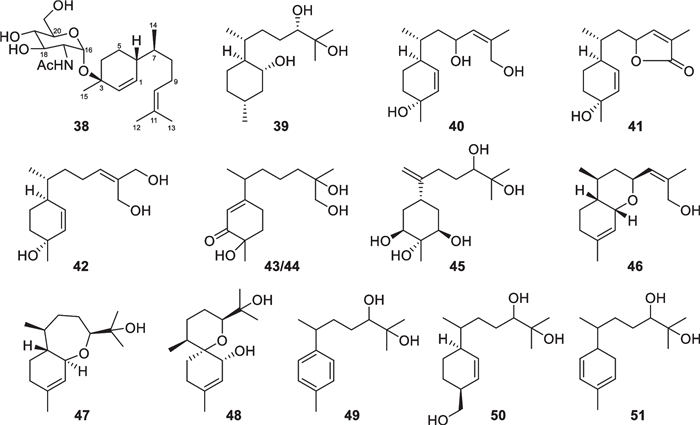
Bisabolane sesquiterpenes from marine-derived Trichoderma
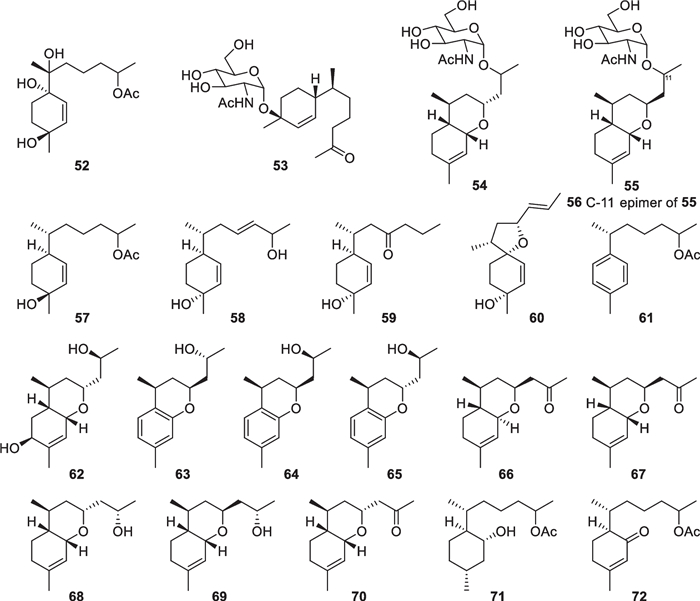
Norbisabolane sesquiterpenes from marine-derived Trichoderma
As secondary metabolites of some Trichoderma and Fusarium species, trichothecane sesquiterpenes have been regarded as a class of mycotoxins for animals and humans [20]. Besides the known trichodermin, 18 trichothecane derivatives (73–90, Fig. 11) were isolated from marine-derived Trichoderma species, including T. brevicompactum and T. hamatum as well as an unidentified strain [31, 40–43]. Similar to the known trichothecane sesquiterpenes [135], the majority of these new isolates possess a 2,11-epoxy unit. However, trichodermol chlorohydrin (82) and trichodermarin N (90) are exceptions, with the former being the first natural halogenated trichothecane derivative. Both 2,11-epoxy and 11,12-epoxy units are present in trichodermarin H (84), of which the absolute configuration along with that of trichodermarin G (83) was assigned by X-ray crystallography. The absolute configurations of 77–80 were ascertained by analysis of their ECD data aided by quantum chemical calculations, while that of 81 was determined by agreement of its specific optical rotation with the hydrolysate of 77. It is interesting that glycosides are also not absent in this class of metabolites. Three members (88–90) were identified to possess sugar moieties, with 90 being the first glucosamine-coupled trichothecane. It is worth noting that the production of trichothecane toxins may decrease the application of their producers, though some strains display excellent antifungal effects [136].
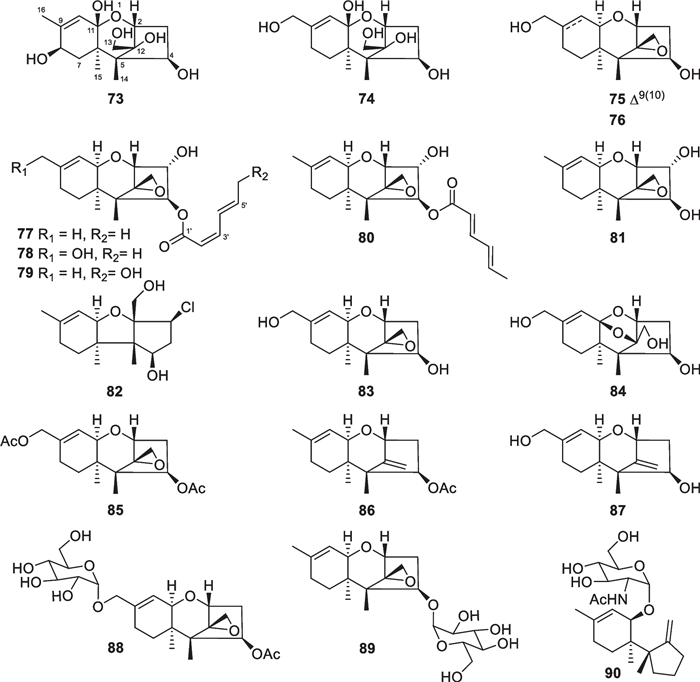
Trichothecane sesquiterpenes from marine-derived Trichoderma
Carotane, also designated daucane, sesquiterpenes with 10 new members (91–100, Fig. 12) were purified from two strains of the marine algicolous T. virens [44, 45]. Among Trichoderma species, T. virens is the main producer of carotane sesquiterpenes, but their origin is not confined to only this species. A soil-borne T. crassum strain was also reported to yield carotanes [137]. All these 10 isolates are oxygenated at C-3 and C-4, and the peculiarity is a carbonyl group, rather than a hydroxy group, at C-3 of trichocarotin C (93). The absolute configuration of trichocarotin A (91) was determined by analysis of X-ray crystallographic data, while those of trichocarotin B (92) and 14-O-methyltrichocarotin G (99) were established by ECD spectra. Although other filamentous fungi, such as Byssochlamys, Penicillium, and Aspergillus species, also produce carotane sesquiterpenes, the oxidation at C-3 and C-4 are uncommon [138]. Thus, the oxygenated carotanes at both C-3 and C-4 have chemotaxonomic significance for their Trichoderma producers.
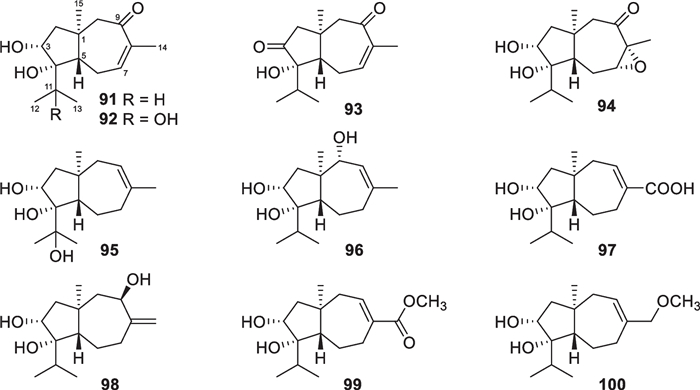
Carotane sesquiterpenes from marine-derived Trichoderma
Besides carotanes, cadinane sesquiterpenes also exist in T. virens [44, 45]. However, this class of sesquiterpenes (101–116, Fig. 13) are distributed in a broader spectrum of other Trichoderma species, such as T. asperelloides [34], T. asperellum [46], T. harzianum [48], and an unidentified strain [47]. The first untouched cadinane sesquiterpene trichocadinin A (101) from Trichoderma was reported in 2018, even if a 2,3-seco derivative, named heptelidic acid, was found from T. viride in 1980 [20]. The 2,3-seco cadinane sesquiterpenes (114–116) were also obtained from two other Trichoderma species of marine origin, with halogenation being present in 114. Moreover, trichocadinins Ⅰ (112) and J (113) possess a 2-norcadinane framework. Except for 102–104, these cadinane derivatives feature a carboxyl group at C-11. Relying on the presence of a conjugated carboxyl group, the absolute configurations of 101 and 105–115 were established by ECD spectra. Additionally, the absolute configuration of 104 was determined by X-ray diffraction. Although cadinane sesquiterpenes have occurred frequently in plants and fungi, the ring-opening derivatives are distributed narrowly in nature [139].
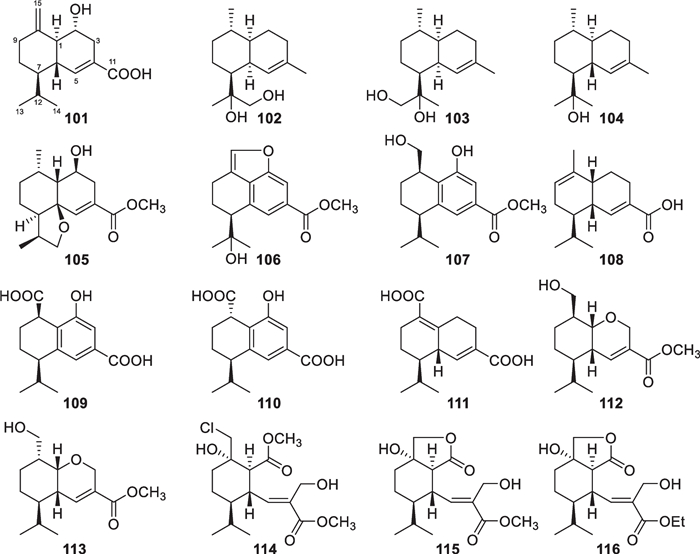
Cadinane sesquiterpenes and catabolites from marine-derived Trichoderma
Other 12 sesquiterpenes (Fig. 14) with acorane (117–119) [30, 37, 49], cuparane (120 and 121) [43], farnesane (122 and 123) [37], synderane (124) [50], norpupukeanane (125) [51], harzianoic acid (126 and 127) [52], and ethylated bisabolane (128) [26] frameworks were discovered from T. harzianum, T. brevicompactum, T. asperellum, T. longibrachiatum, and an unidentified strain. 8-Acoren-3,11-diol (117), trichoacorin A (118), and trichoacorside A (119) are the only three spiro-fused sesquiterpenes characterized from marine-derived Trichoderma, with 119 being the first acorane aminoglycoside. Trichocuparins A (120) and B (121) represent the first two Trichoderma-derived cuparane derivatives, and the absolute configuration of the former was determined by analysis of X-ray crystallographic data. This carbon skeleton harbors the same ring system as trichothecane, but one of the four methyl groups resides at a different position. Trichonerolins A/B (122/123), trichodermoside (124), and trichodermene A (125) are also firstly occurring structural types in Trichoderma metabolites, especially an aminosugar unit in 124 and a complicated ring system in 125. There are two chiral centers in 122 and 123, of which the chemical shift deviations undoubtedly arise from their epimeric relationship. Harzianoic acid B (127) features a new natural scaffold with a cyclobutane nucleus, and harzianoic acid A (126) is its 15-nor derivative. In addition, the ethylated bisabolane skeleton of trichaspin (128) is unprecedented, too. Most of these skeletons have been discovered in only one or two species, their universality in marine-derived Trichoderma needs to be further explored.
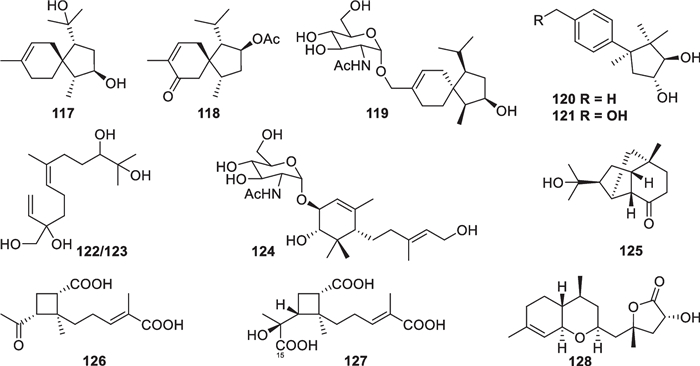
Other sesquiterpenes from marine-derived Trichoderma
The number of new diterpenes with seven basic carbon skeletons from marine-derived Trichoderma amounts to 37 (129–165, Table 1) [26, 30, 32, 46, 53–65]. Of those, 25 members (129–153, Fig. 15) are harziane derivatives, that were isolated from T. asperellum [26, 46], T. longibrachiatum [53, 54], T. harzianum [30, 56, 57], T. asperelloides [59], T. erinaceum [32, 60], and two unidentified strains [55, 58]. The common harziane skeleton harbors a fused cyclobutane, cyclopentane, cyclohexane, and cycloheptane ring system, that is unique in nature. Harzianone (129) was reported as the second harziane diterpene in 2012, 20 years later than the discovery of harziandione with a second carbonyl group at C-3. The absolute configuration of 129 was ascertained by quantum chemical calculations of ECD data, and that of harziandione was confirmed by simulation of the specific optical rotation at the same time [53]. Other isolates with this rigid scaffold have different oxidation degrees and positions, and the double bond at C-9 is hydrogenated in 144–146 in particular. The first total synthesis of a harziane diterpene was reported in 2020, with highly diastereocontrolled construction of the cyclobutane ring via enyne cycloisomerization being a key step [140]. This work also resulted in configurational revision at C-9 of the target harzian-9-ol, isolated from the tree-associated T. atroviridae. Harzianelactone (147) and trichodermaerin (150) represent the first 11,12-lactone and 10,11-lactone, respectively, that are possibly produced by catalysis of the Baeyer–Villiger monooxygenase [58, 60]. X-ray diffraction with Mo Kα radiation secured the relative configuration of 150, that was isolated from T. asperellum later [141]. Lactonation seemingly happens only in the four-membered ring, and a subsequent reaction may yield ring-opening products, such as 153 [59]. In addition, two proharziane diterpenes (154 and 155) with a 14-membered macroring being inlaid with a cyclohexane unit were identified from T. asperelloides and T. harzianum [30, 59], and they seem intermediates during the biosynthesis of corresponding harzianes [142]. The absolute configurations of all these 27 isolates were confirmed by chiral techniques. As the assignment of 129, ECD analysis played an important role in determining the absolute configurations of most isolates. X-ray diffraction was also used to determine the absolute configurations of several molecules, including 131, 132, 140, and 142. The distribution of harziane, secoharziane, and proharziane diterpenes is not confined to only the above Trichoderma species, and they rarely occur in other fungal genera. Thus, these diterpenes with tetracyclic and bicyclic scaffolds are promising to be regarded as biomarkers for Trichoderma fungi.
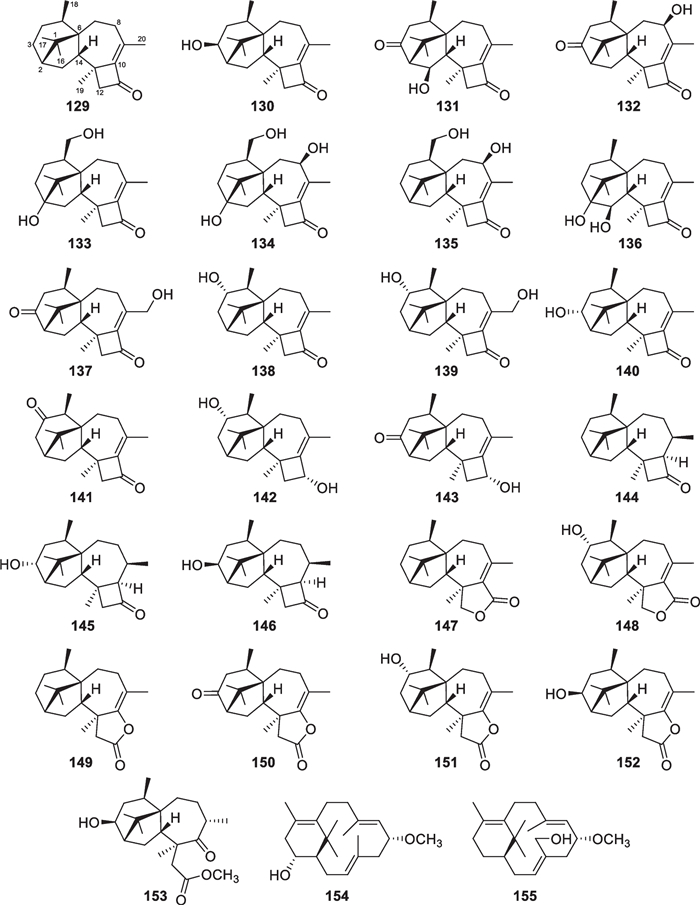
Harziane, secoharziane, and proharziane diterpenes from marine-derived Trichoderma
Wickerane diterpenes look like exclusive metabolites of Trichoderma, six members (156–161, Fig. 16) were discovered from sponge-derived T. harzianum [61, 62]. Their distribution is not as broad as that of harziane derivatives. T. atroviride of soil or plant origin contributed the first two wickerane diterpenes, wickerols A and B (also named trichodermanin A), and their biogenetic route was predicted with 13C-labeled acetate [143, 144]. In the pathway, the verticillyl cation is the same as the intermediate in the biosynthesis of harziane diterpenes [142, 143]. The complicated tetracyclic scaffold and its relative configuration were guaranteed by X-ray crystallographic analysis [144]. A stereocontrolled synthesis from commercial sitolactone led to the assignment of absolute configurations and the revision of the original specific optical rotation sign of wickerol A [145]. Compared to wickerols A and B, trichodermanins C–H (156–161) feature high degrees of oxidation. The lack of any olefinic and aromatic unit is also characteristic of these diterpenes. The absolute configurations of 156, 157, and 159 were determined by application of the modified Mosher’s method, while that of 158 was established by the negative Cotton effect at 258 nm in the ECD spectrum of its dibenzoate. In view of the narrow distribution spectrum, this class of diterpenes may also be potential biomarkers for several Trichoderma species.
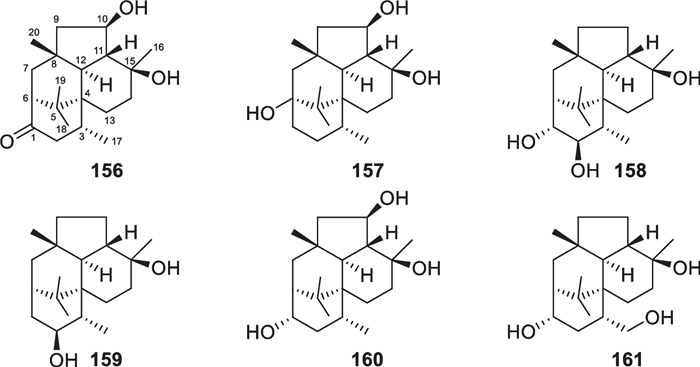
Wickerane diterpenes from marine-derived Trichoderma
Besides the above harziane and wickerane, there are also four diterpene skeletons (Fig. 17) occurring in marine-derived Trichoderma. Citrinovirin (162) with a new norditerpene skeleton and trichocitrin (163) with a fusicoccane skeleton were isolated from T. citrinoviride [63, 64]. Harzianolic acid A (164) with a cleistanthane skeleton and trichosordarin A (165) with a 15-nor transformed sordaricin skeleton were obtained from T. harzianum [56, 65]. The latter three metabolites (163–165) are firstly occurring structural types in this genus, and the furan ring in 163, the chlorine atom in 164, and the deoxymonosaccharide in 165 greatly enhance their novelty.
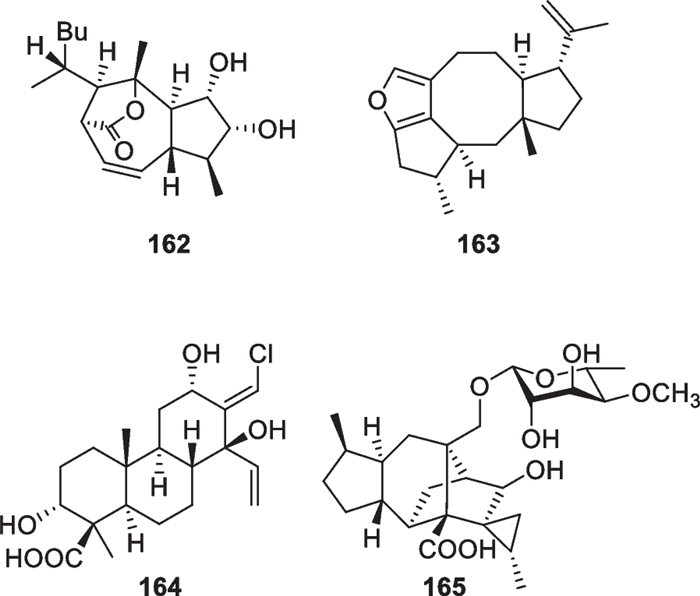
Other diterpenes from marine-derived Trichoderma
2.2 Steroids
Compared to the high number of terpenes, only three new steroidal metabolites (166–168, Fig. 18) with ergostane and its transformed and degraded skeletons were discovered from marine-derived Trichoderma species, including T. brevicompactum [37], T. asperellum [66], and T. atroviride [39]. It is worth mentioning that all these isolates feature high degrees of oxidation, that even results in the broken ergostane scaffolds in 167 and 168. Considering the structural complexity, the assignments of absolute configurations of 167 and 168 were completed by both ECD and X-ray crystallographic analyses. Unfortunately, no steroidal metabolites of the viridin series have been found in marine-derived Trichoderma [20].
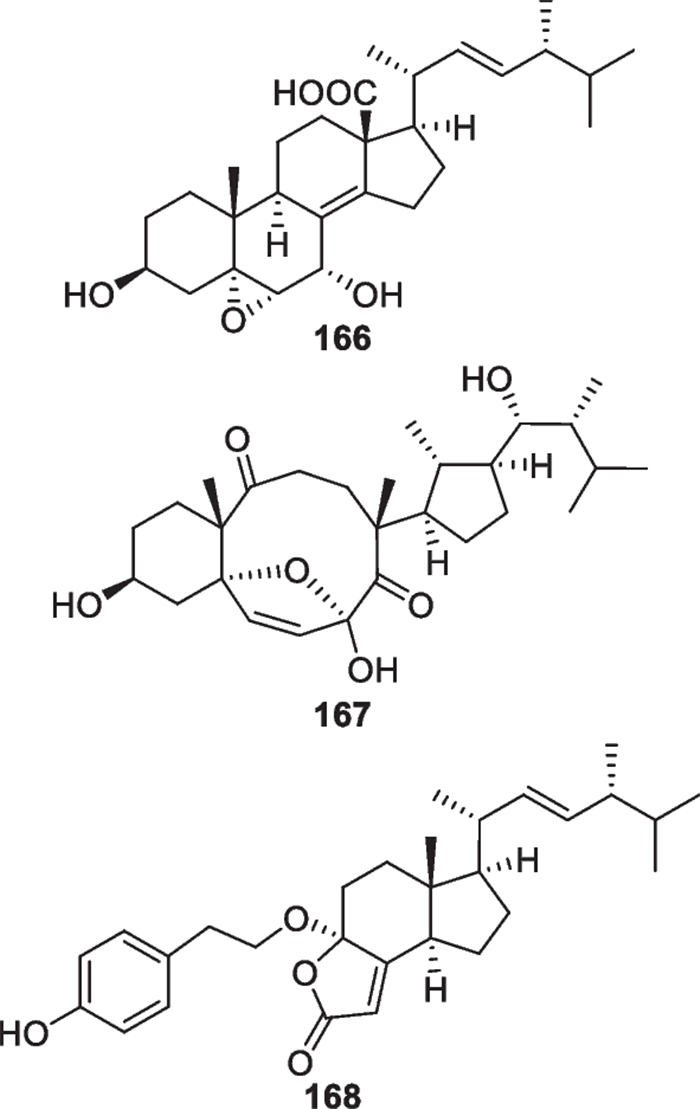
Ergosterol derivatives from marine-derived Trichoderma
2.3 Polyketides
Despite the lower number of new polyketides (169–295) than that of terpenes, they have more prolific sources. Totally, 14 marine-derived Trichoderma species, including T. asperellum (2 compounds), T. atroviride (6), T. aureoviride (2), T. citrinoviride (3), T. erinaceum (6), T. harzianum (27), T. koningii (9), T. longibrachiatum (3), T. reesei (32), T. saturnisporum (8), T. virens (3), T. viride (1), H. lixii (1), and H. vinosa (2), and nine unidentified strains (22) have the ability to produce polyketides [24, 25, 32, 33, 39, 49, 67–106]. These metabolites can be classified into cyclopentenone, sorbicillinoid, koninginin, decalin, xanthone, anthraquinone, naphthopyrone, and other acyclic and cyclic categories, with several novel skeletons being present.
As shown in Fig. 19, four pairs of C13 lipids named harzianumols A-H (169–176) were isolated from the sponge-associated T. harzianum [67]. Each pair of them occurred as an inseparable enantiomeric mixture, and their absolute configurations were identified through the modified Mosher’s method. As a possible derivative of lauric acid, oxylipin 177 was discovered from the alga-derived T. atroviride [39]. Additionally, two methyl-branched lipids (178 and 179) with the same backbone were identified from T. citrinoviride, and the latter contains a unique 1,3-dioxolane nucleus [68, 69]. Nafuredin C (180) with a δ-lactone unit was obtained from the mangrove-endophytic T. harzianum, occurring as the third member of this structural family [70]. An algicolous T. citrinoviride strain can also yield a member of this type [64], but the producers of nafuredins are not only Trichoderma but also Aspergillus, Penicillium, and Talaromyces [146, 147]. T. harzianum of sponge origin gave harzialactone B (181), of which the δ-lactone moiety seems formed through the Baeyer–Villiger oxidation of a cyclopentenone precursor [71].
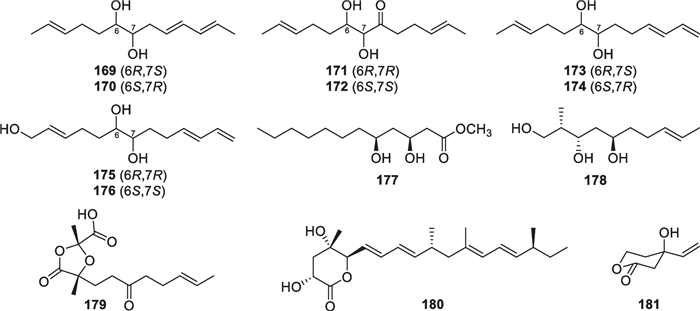
Acyclic polyketides and lactones from marine-derived Trichoderma
Cyclopentenone derivatives with 15 members (182–196, Fig. 20) were discovered from T. harzianum [71, 72], T. asperellum [25], T. koningii [74, 77], T. atroviride [75, 76], and two unidentified strains [73, 78]. This class of metabolites are structurally simple, and all of them contain an α, β-unsaturated carbonyl group in the five-membered ring. Seven (182–188) of the isolates possess only seven carbon atoms, with one olefinic carbon adjacent to the carbonyl group tending to be chlorinated. One more carbon atom is present the nucleic moieties of 189–195, which exist as esters or acids without a chlorine atom at α position of the carbonyl group. The structure of 196 differs from those of the others by the presence of a long side chain and two methyl groups on the cyclopentenone nucleus. The configurational assignments of 182–184 were achieved by total syntheses, with the former one being speculated as scalemic enantiomers by consideration of its smaller specific rotation than that of the synthesized one [72]. On the other hand, the conjugated carbonyl group facilitates the stereochemical determination of some chiral members, such as 186, 187, and 196, by ECD data [25, 78]. In spite of the similarity between cyclopentenones and isonitriles of Trichoderma origin, the former should be synthesized through polyketide synthases, but the latter were deduced to be derived from tyrosine [148].
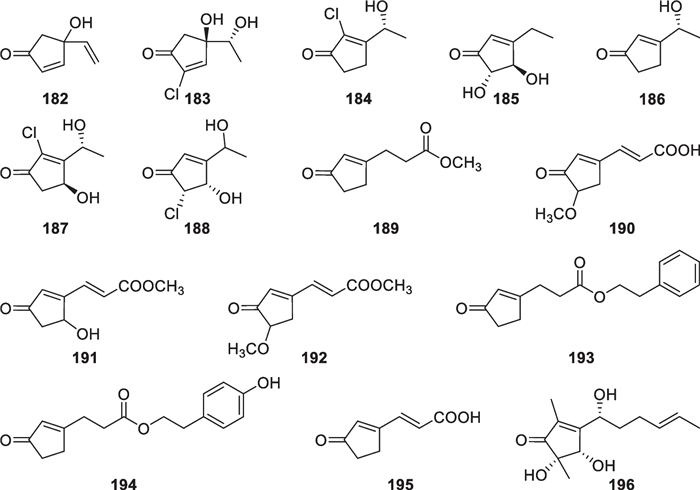
Cyclopentenone derivatives from marine-derived Trichoderma
As the largest group of polyketides from marine-derived Trichoderma, sorbicillinoids and their derivatives consist of 47 new members. Monomeric sorbicillinoids (197–217, Fig. 21) from T. reesei [81, 82], T. saturnisporum [84], T. longibrachiatum [49, 85], and three unidentified strains [79, 80, 83] are the most primitive representatives, and the majority of them are constructed with a phenyl core substituted by a sorbyl-derived acyclic or epoxy side chain and one or two methyl groups. Methylation happens at C-3 and/or C-5 before cyclization of the hexaketide chain by the Claisen reaction [149], and hydroxylation commonly occurs at C-2 and/or C-4. However, the C-5 position of 206 is also bonded to a hydroxy group, rather than a methyl group. 202, 209, 210, and 214 harbor nonstandard sorbyl chains, possibly arising from the length change in original polyketide chains or oxidation scission of C6 units. Based on biogenetic considerations, saturnispol E (216) and epoxysorbicillinol (217) with a cyclohexene core are possible deoxy and epoxy derivatives of the tautomeric intermediates of sorbicillin, while (-)-trichodermatone (215) is a regioisomer of sorbicillinol [149]. In addition, 12 dimeric sorbicillinoids (218–229, Fig. 22), also called bisorbicillinoids, were obtained from T. reesei [81, 82, 86], T. saturnisporum [84], and one unidentified strain [79]. All the isolates are heterogeneous dimers, and their skeletal dimerization happens between C-3 of one monomer and C-6 of the other. Their structures appear more complicated, especially the unique cage-like core in 218 and 219. These dimers were proposed to be constructed through intermolecular single or double Michael reaction/ketalization, with sorbicillinol or its analogs serving as key intermediates [150]. Apart from the above monomers and dimers, there are also 14 other sorbicillinoid derivatives (230–243, Fig. 23) from marine-derived T. saturnisporum [84], T. reesei [82, 86], and two unidentified strains [83, 87]. 2,3-Dihydro 2-hydroxy vertinolide (230) and saturnispol F (231) belong to the vertinolide subfamily, with the lactone unit being probably formed via intramolecular esterification and ring cleavage sequences. Saturnispols C (232) and D (233) feature a bicyclo[2.2.2]octanedione motif that arises from a Diels–Alder [4 + 2] cycloaddition between the corresponding sorbicillinoid monomer and phenylethylene. Based on the same mechanism, the formation of each trichodermanones A-D (234–237) appears rationalized by reaction between sorbicillinol and a dienophile. The last six members (238–243) differ greatly from the others, due to the presence of a methylene group between the sorbyl chains and the six-membered ring. Moreover, a naphthalene ring is present in 238 and 239, and a bicycle [3.2.1] lactone unit appears in 240–243. During the identification of chiral isolates, ECD spectra were widely used to confirm their absolute configurations, and X-ray diffraction was also used for several members, such as 200, 202, 206, and 214. A total synthesis of (±)-epoxysorbicillinol was fulfilled from commercial diethyl methylmalonate in 13 steps, simultaneously permitting epoxidation and avoiding aromatization [151]. A conversion from sorbicillin to (±)-epoxysorbicillinol was also achieved via the formation of a p-quinol intermediate by an oxidative dearomatization [152]. To date, the number of sorbicillinoid members exceeds 130, and they have been discovered from no less than 10 genera of ascomycetes [86, 153]. It is interesting that all the known marine-derived Trichoderma species that produce sorbicillinoids belong to the Longibrachiatum clade [154].
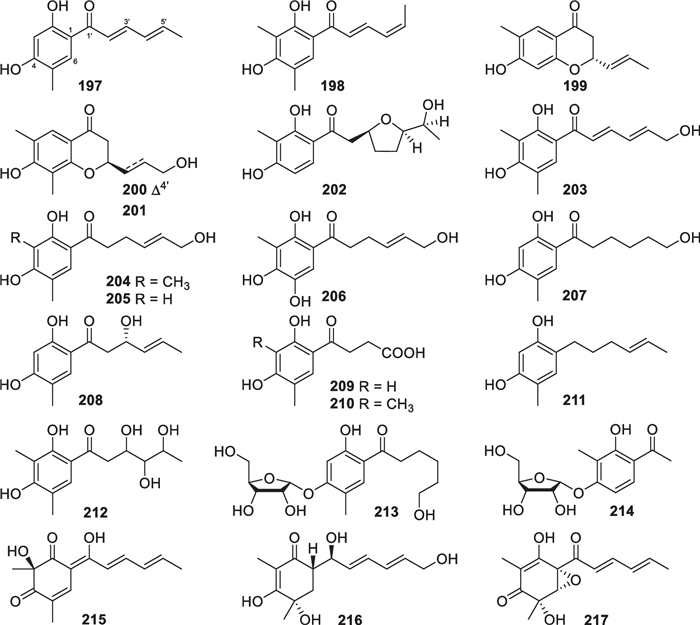
Monomeric sorbicillinoids from marine-derived Trichoderma
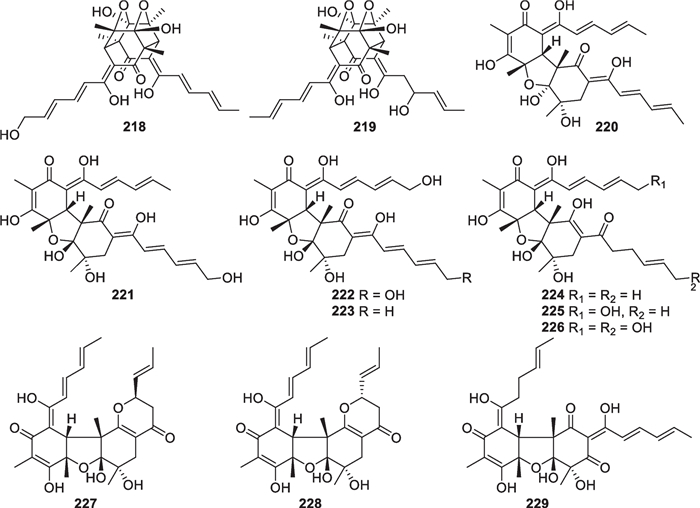
Dimeric sorbicillinoids from marine-derived Trichoderma
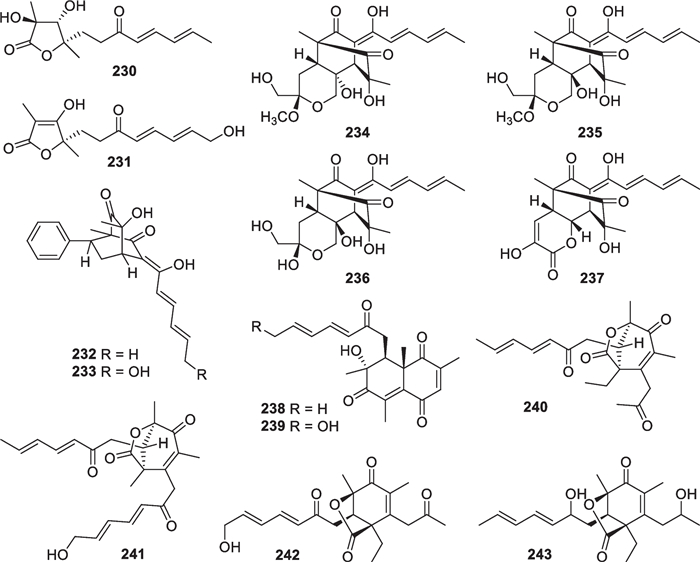
Other sorbicillinoid derivatives from marine-derived Trichoderma
As a family of octaketides, koninginins with a narrow distribution are also the representative metabolites from Trichoderma. To date, 12 new members (244–255, Fig. 24) have been isolated from marine-derived T. reesei [89], T. koningii [91], H. lixii [90], and one unidentified strain [92]. The first koninginin, named koninginin A, was obtained from T. koningii in 1989 [20], and it defined the typical koninginin skeleton that contains a benzopyran nucleus and a heptyl side chain. This skeleton is present in 245–249, especially a highly unsaturated chromone motif in 248. In 244, a second cyclohexene ring is incorporated into this basic structure, forming an unprecedented pentacyclic framework with a ketalic and a hemiketalic carbon. Starting from L-tartaric acid, a synthetic route for this complicated molecule, that harbors eight chiral carbons, was developed in a stereocontrolled manner [155]. Instead of the pyran ring, one or two furan rings exist in 250–255. The former two are unprecedented due to the presence of a ditetrahydrofuran-bearing tricyclic skeleton. Although ECD spectra were applied to confirming the absolute configurations for most of the isolates [89, 91, 92], the relative configurations between the chiral carbon in cyclohexene ring and that in pyran or furan ring failed to be given in some structures due to the lack of any NOE correlation. Chemical syntheses suggested that epimeric pairs possess very similar spectroscopic data, which also make it difficult to distinguish their relative configurations by comparison of spectroscopic data [156].
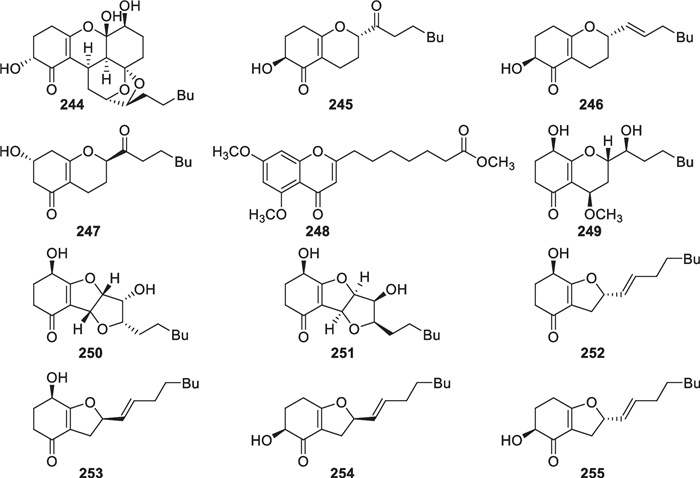
Koninginins and their derivatives from marine-derived Trichoderma
Decalin derivatives, such as trichosetin and compactin, were previously detected in several Trichoderma species [20]. Marine-derived T. harzianum along with an unidentified strain gave another type of decalin derivatives (256–268, Fig. 25), with a C3 and a C4 side chain as well as two methyl groups [24, 93–98]. The bicyclic nucleus is a trans-fused ring system, with a double bond at C-6. Acylation of hydroxy groups on the decalin core and the C3 side chain contributes to the structural diversity of this octaketide type, especially the formation of the dimer 266. Except for trichoharzianin (256), the acyl groups are confined to (Z)- and (E)-3-methylpent-2-enedioic acid (3-methylglutaconic acid) and their monoesters, which are presumed to arise from mevalonic acid [24]. A tyrosol residue that can be converted from tyrosine exists in 265 [157], similar to 168 and 194. The absolute configurations of most members were assigned by chemical conversion, including alkali-hydrolysis to yield triols or further acylation to furnish tribenzoates. Dimolybdenum tetraacetate [Mo2(OAc)4] was used to form complexes with the cis vic-diol group in hydrolysates of 257–259, and analyses of their ECD spectra resulted in determination of the absolute configurations. Analogs with the same or similar alkylated decalin skeletons were also identified from other fungal species of the genera Eupenicillium, Fusarium, Geomyces, Phoma, Spopormiella, and Stemphylium [95, 96], but the 3-methylglutaconate units were rarely encountered therein.
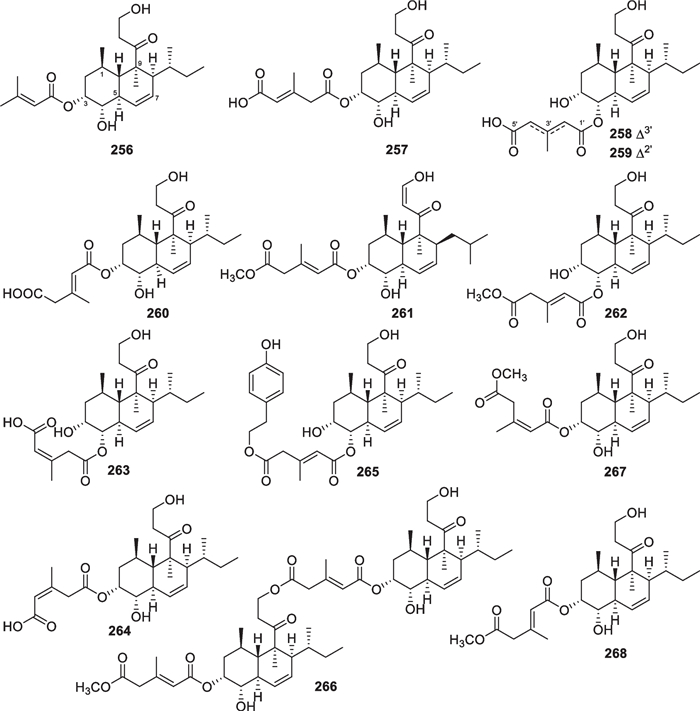
Decalin derivatives from marine-derived Trichoderma
Xanthone, anthraquinone, and naphthopyrone derivatives have been discovered from multifarious plants and/or fungi [158, 159], but their distribution in Trichoderma is not much attractive. Only nine new members (269–277, Fig. 26) were obtained from marine-derived Trichoderma, including T. aureoviride [99], T. harzianum [100], H. vinosa [102], and one unidentified strain [101]. Trichodermaxanthone (269) contains a typical xanthone skeleton, which rarely occurs in the metabolites of this genus. Octaketides 270–273 possess a basic anthraquinone scaffold, but one of the two carbonyl functionalities is reduced to a hydroxy group in epimers 271 and 272. Considering the similarity between 274 and 275, they possibly arise from the same heptaketide intermediate. Hypochromins A (276) and B (277) have a dimeric naphtho-γ-pyrone framework, which are present in a broad spectrum of ascomycetous producers [160]. There is one or more chiral centers in 271, 272, and 275, of which the configurations were determined by interpretation of ECD spectra. A crystallographic analysis further confirmed the absolute configuration of 271. In addition, 276 and 277 with atropisomerism are axially chiral molecules, and the single bond between the two monomers was assigned as S-configuration by the exciton chirality method. These highly unsaturated isolates exhibit yellow or red colors, which may be taken as pigments by their producers.
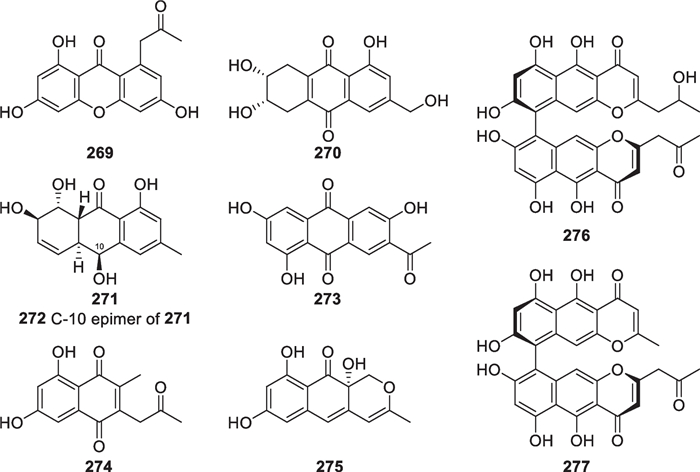
Xanthone, anthraquinone, and naphthopyrone derivatives from marine-derived Trichoderma
In addition to the above sorbicillinoid, xanthone, anthraquinone, and naphthopyrone derivatives, a few other aromatic polyketides (278–295, Fig. 27) with one or two benzene, furan, and/or pyran rings have been isolated from marine-derived Trochoderma. Trichorenins A-C (278–280) from T. virens harbor a 5/5/6/5-fused ring system, probably arising from the cyclization of an octaketide intermediate [103]. This tetracyclic scaffold had been reported previously, but it is contained in a diterpene with five methyl groups formed through a mevalonate pathway [161]. These two skeletons with the same ring system were guaranteed by single-crystal X-ray diffraction analysis, respectively [103, 161]. The structure of harzialactone A (281) with a benzylfuran motif from T. harzianum is shared in 282–286 from T. atroviride and T. erinaceum [32, 71, 104, 105]. Except for 282, these analogous metabolites adopt the same 2R and 4R configurations. As established by the modified Mosher’s method, the original 2S and 4S configurations of 281 were corrected by synthesis from monoacetone-D-glucose via a regioselective oxidation [104]. Later on, this compound and its stereo isomers were further synthesized from simple benzene derivatives through several other stereoselectively chemical and enzymatic routes [162–164]. The absolute configurations of 283–286 were assigned by comparison of experimental and calculated specific optical rotation data [32]. It is worth mentioning that polyketides of this series have been found not only in Trichoderma but also in Aspergillus [71]. Two diphenyl ethers, including a symmetrical (287) and an asymmetrical derivative (288), were identified from T. erinaceum [32]. This class of metabolites have been detected in various fungal genera, especially in Aspergillus and Penicillium [165, 166], and they were proposed to be formed through polyketide pathways by analysis of the biosynthetic gene cluster [167]. As a metabolite of T. citrinoviride, trichophenol A (289) is the only 3-phenylisocoumarin member from this genus [33]. Its structure appears similar to flavonoids, and the isocoumarin nucleus is also present in azaphilones of Trichoderma origin [20]. A chromone unit exists in 290 and 291, which were isolated from an unidentified Trichoderma strain of hydrothermal vent sediment origin [94]. They are structurally similar to 248, but their substitution patterns and side chain lengths appear different. Four simple α-pyrone derivatives, named trichoharzianone (292), trichopyrone (293), and saturnispols G (294) and H (295), were obtained from T. harzianum, T. viride, and T. saturnisporum [84, 93, 106]. The latter two were postulated to be yielded by elimination of a C3 fragment from the precursor sorbicillinol, followed by a lactonization reaction [84].
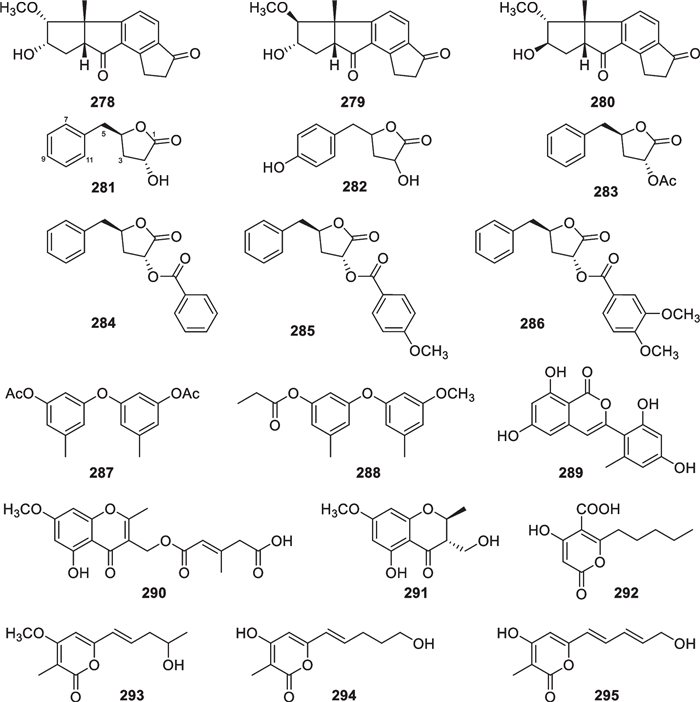
Other aromatic polyketides from marine-derived Trichoderma
2.4 Peptides
Comprising 131 new members (296–426), peptides are the second largest family of metabolites from marine-derived Trichoderma. They were identified from seven Trichoderma species, including T. asperellum (42 compounds), T. atroviride (41), T. harzianum (1), T. longibrachiatum (32), T. orientale (1), T. reesei (1), T. virens (4), and two unidentified strains (9) [25, 35, 41, 70, 107–124]. Their structural types involve cyclic and linear peptides, with the latter being composed of dipeptides, aminolipopeptides, peptaibols, and other peptaibiotics.
Cyclopeptides contain 10 diketopiperazines and one cyclotetrapeptide (296–306, Fig. 28), discovered from T. asperellum [25, 35], T. virens [109], T. atroviride [110], T. reesei [111], and one unidentified strain [41, 107, 108]. Among them, seven diketopiperazines (296–302) contain sulfur atoms, especially a disulfide bridge in 296–298 and a trisulfide bridge in 299 and 300. One of the two modified amino acid residues is substituted by a bromine, an iodine, and a chlorine atom in 296, 297, and 300, respectively, and these halogenated moieties also feature a rarely occurring 1,2-oxazine ring. During the structure elucidation of 301 and 302, the 1H and 13C NMR signals of thiomethyl groups were found to resonate at δH 1.67–2.45 and δC 12.7–18.0, differing greatly from those of oxygen- and nitrogen-bearing methyl groups. ECD spectra played an important role in determining absolute configurations for these sulfides. Three other diketopiperazines (303–305) possess at least one modified amino acid residue, and the symmetrical member (304) was assigned the absolute configuration by X-ray diffraction. Trichoderide A (306) represents the only cyclotetrapeptide from marine-derived Trichoderma, and all the amino acid residues were determined to feature R-configuration on the basis of acid hydrolysis followed by chiral HPLC analysis. In spite of the difference in ring sizes, both 305 and 306 have ornithine and succinic acid residues.
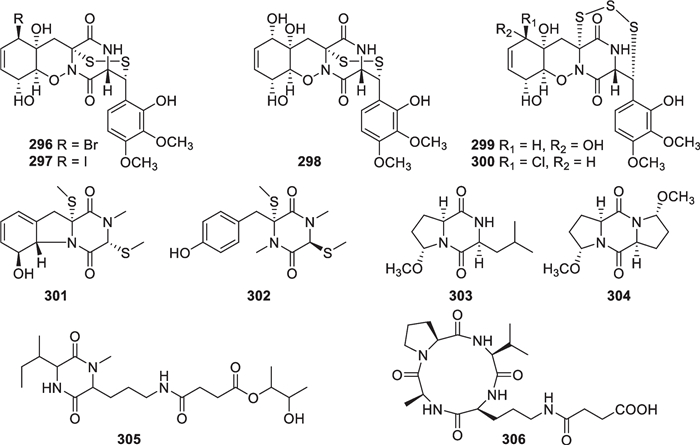
Cyclopeptides including diketopiperazines from marine-derived Trichoderma
Four linear dipeptides (307–310, Fig. 29) were obtained from T. virens and T. harzianum as well as an unidentified strain [70, 112, 113]. Despite the different sources, these isolates exhibit high structural similarities. Besides the 1,2-oxazine ring that exists in diketopiperazines 296–300, a coumarin motif substituted by two methoxy groups is also present in these four metabolites. The connectivity of oxazine and α-pyrone functionalities in 307 was confirmed by X-ray diffraction analysis. The difference between 307 and 308 is the replacement of a hydroxy group in the former by a chlorine atom in the latter. The chemical total syntheses of 307 and 308 as well as their analogs were achieved by at least three groups, and their focus was the construction of the oxazadecalin core [168–172]. Trichodermamide G (309) and dithioaspergillazine A (310) also feature a sulfide bridge with one or two sulfur atoms, respectively, located in only one amino acid residue. These two sulfides were assigned absolute configurations by analysis of their ECD spectra aided by quantum chemical calculations. A series of dipeptides with the same or similar structures have also been discovered from other fungal genera, such as Aspergillus, Penicillium, and Spicaria [173–175].
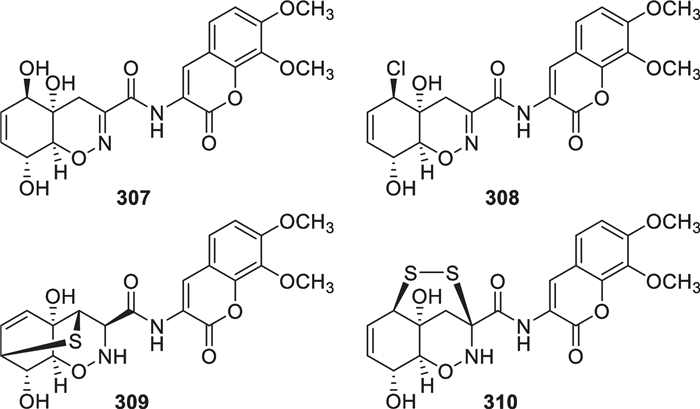
Dipeptides from marine-derived Trichoderma
Marine-derived Trichoderma species have contributed a large number of linear polypeptides (311–426, Figs. 30, 31, 32, 33, 34) with 9 to 20 amino acid residues, accounting for ca. 10% of all the Trichoderma-derived peptaibiotics (over 1000) [176, 177]. In view of the definitions given in literature [178], all of them can be sorted into aminolipopeptides, peptaibols, and other peptaibiotics, characterized by high contents of α-aminoisobutyric acid (Aib) and acylated N-termini. In the aminolipopeptide subfamily, there are only three members (311–313), arising from an unidentified Trichoderma strain [114]. Besides four Aibs, the non-proteinogenic 2-methyl decanoic acid, 2-amino-6-hydroxy-4-methyl-8-oxodecanoic acid (AHMOD), and 2-[(2′-aminopropyl) methylamino] ethanol residues are present in all the three isolates. These chiral units in 311 were assigned the absolute configurations by total synthesis [115]. Besides an α-methyl-branched fatty acid, the linkage between proline and a lipoamino acid residue at N-terminus conforms to the standard of this type. In the peptaibol subfamily, a total of 83 members (314–350 and 352–397) with 9, 10, 11, 19, and 20 residues were identified from four marine-derived Trichoderma species, including T. asperellum [116–118], T. longibrachiatum [119, 120, 123], T. atroviride [121, 122], and T. orientale [124]. Each member contains a β-amino alcohol or its acetate at C-terminus, derived from the reduction of the corresponding amino acid precursor [121]. The identification of 314–321 and 386–393 was performed by analysis of the NMR and mass spectroscopic data of pure compounds, and X-ray diffraction was used to determine the structure and relative configuration of 317. Their absolute configurations were confirmed by acid hydrolysis followed by derivation with Ru(D4-Por*)CO or Marfey’s reagent [116, 117, 122]. Other peptaibol isolates were mainly identified by interpretation of mass spectra given by various techniques, such as ultrahigh pressure liquid chromatography/electrospray ionization tandem mass spectrometry (UHPLC/ESI–MS/MS) [118], electrospray ionization ion-trap mass spectrometry (ESI-IT-MS) [119–121, 123, 124], collision-induced dissociation mass spectrometry (CID-MS) [120], and gas chromatography/electron impact mass spectrometry (GC/EI-MS) [119, 120, 123]. Although 351 from T. asperellum was claimed as a peptaibol in literature, it should belong to the peptaibiotic subfamily due to the lack of an amino alcohol at C-terminus [118]. Additionally, 29 unprecedented peptaibiotics (398–426) with 17 amino acid residues were identified from T. atroviride [121]. In addition to nine constant amino acid residues, a peculiar residue with a mass of 129 Da appears at C-terminus of these peptaibiotics. Unfortunately, the differentiation between some amino acid residues and their isomers, such as leucine/isoleucine, valine/isovaline, leucinol/isoleucinol, and valinol/isovalinol, remains not completed in many peptaibols and other peptaibiotics. Natural peptaibiotics have been found in some 30 known genera of fungi [121], especially in mycoparasitic ones of the Hypocreales [177].
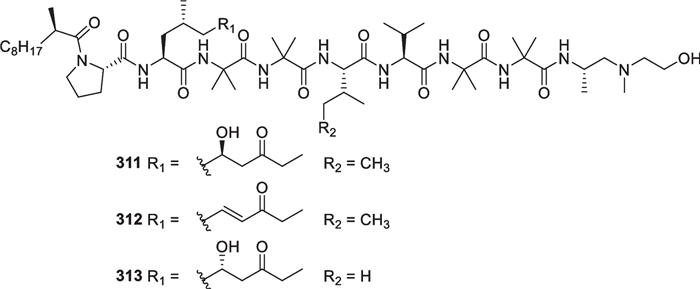
Aminolipopeptides from marine-derived Trichoderma
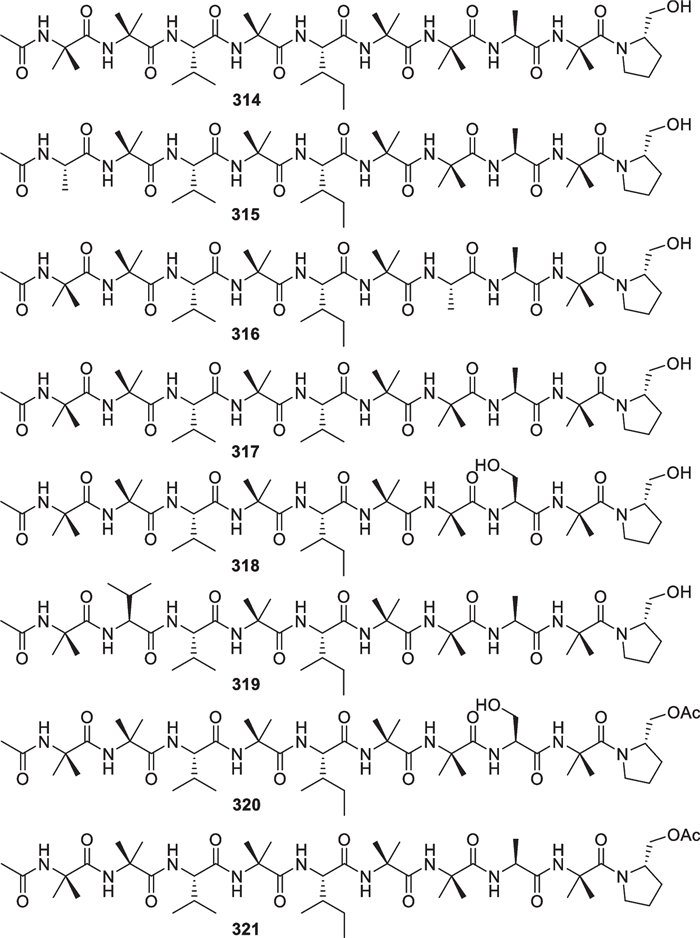
10-Residue peptaibols from marine-derived Trichoderma

9/10-Residue peptaibols and a 10-residue peptaibiotic from marine-derived Trichoderma (Lxx: Leu/Ile)

11-Residue peptaibols from marine-derived Trichoderma (xxx: Val/Iva/Leu/Ile, xxol: Valol/Ivaol/Leuol/Ileol)

19/20-Residue peptaibols and 17-residue peptaibiotics from marine-derived Trichoderma (Lxx: Leu/Ile, Vxx: Val/Iva, xxx: Val/Iva/Leu/Ile, yyy: Aib/Ala/Val/Iva)
2.5 Alkaloids
Except for 17 nitrogen-bearing terpenes and 131 peptides, there are also 14 other nitrogenous metabolites (427–440, Fig. 35) from T. asperellum (1 compound) [25], T. atroviride (2) [105, 110], T. citrinoviride (2) [128], T. harzianum (1) [98], T. reesei (1) [81], T. virens (1) [127], and five unidentified strains (6) [125, 126]. An unidentified Trichoderma strain of deep sea origin (-3300 m) afforded β-carboline alkaloids 427–430, with the latter two being synthesized previously. The absolute configurations of 427, 429, and 430 were determined by comparison of experimental and calculated specific optical rotation data. Trichodins A (431) and B (432) harbor an α-pyridone ring fused with a monoterpene unit. Regardless of the stereochemistry, these two alkaloids seem formed by reaction of deoxy-PF1140 with phenol and ribosylated phenol, respectively [179]. A quinoline motif substituted by both chlorine and bromine atoms exists in the new natural product 433, and an imidazo[1,5-b]isoquinolone tricyclic system is present in 434. As a naturally occurring compound, 4-oxazolepropanoic acid (435) features an oxazole ring, that has also been found in other metabolites of Trichoderma origin [20]. Seven members, including 436–442, contain one or more pyrrole rings. The pyrrolidin-2-one unit in 437 is possibly incorporated into a sorbicillinoid precursor by a Diels–Alder reaction, and this class of molecules are not rich in nature [81]. Tetramic acids that possess a 2,4-pyrrolidinedione ring are widespread in terrestrial and marine organisms [180], but only two members (438 and 439) with the similar scaffold have been discovered from a marine-derived Trichoderma species. One C3 and three C4 units construct the acyclic 440, which partially resembles diketopiperazine 305 of the same origin. In general, most of the heterocyclic units have been reported in other Trichoderma metabolites, but the halogenation in 433 and the ring system in 434 are peculiar to some extent.
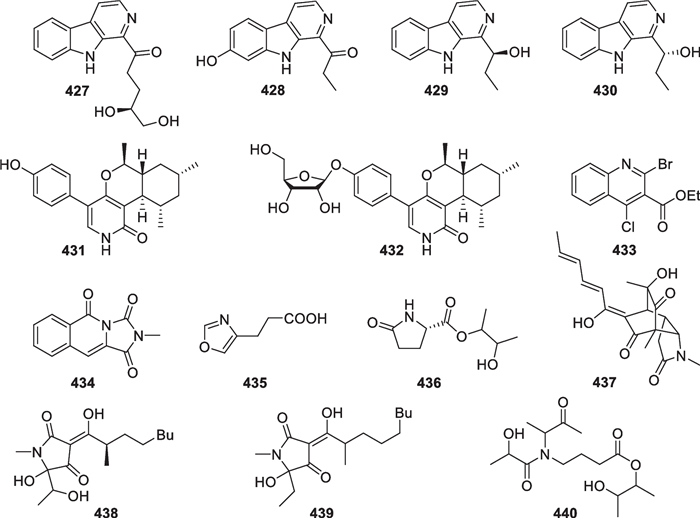
Alkaloids from marine-derived Trichoderma
2.6 Others
Five other metabolites (441–445, Fig. 36), possibly arising from amino acids or sugars, were obtained from T. atroviride (1 compound) [110], T. koningii (3) [77, 129], and T. reesei (1) [130]. Among them, both 441 and 442 possess an acetal linkage due to the reaction of 2,3-butandiol with 4-hydroxyphenylacetaldehyde (4-HPAA) or 5-hydroxymethylfurfural (5-HMF), while 443 seems formed through aldol condensation between 4-HPAA and 5-HMF. 4-HPAA is an intermediate during the conversion of tyrosine to tyrosol via tyramine or 4-hydroxyphenylpyruvate [157], and 5-HMF arises from hexoses, such as glucose and fructose, via dehydration [181]. A tyrosol residue exists in 444, of which the 3-hydroxy-3-(p-hydroxyphenyl)propanoate moiety also looks like a tyrosine derivative. Gliocladinin D (445) is a terphenyl glycoside, and its core is initially condensed between two molecules of 4-hydroxyphenylpyruvate under the catalysis of a tridomain nonribosomal peptide synthetase [182]. Terphenyls have been detected in fungi for a long time [183], but their occurrence in Trichoderma is really poor.
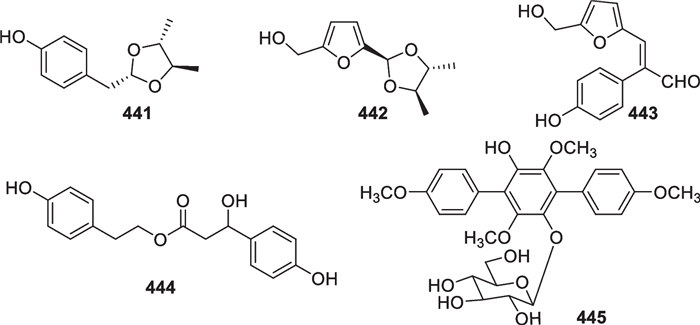
Other metabolites from marine-derived Trichoderma
3 Biological activity
3.1 Antimicroalgal
Eight marine microalgae (phytoplankton), including Amphidinium carterae, Chattonella marina, Heterocapsa circularisquama, Heterosigma akashiwo, Karlodinium veneficum, Prorocentrum donghaiense, Phaeocystis globosa, and Scrippsiella trochoidea, that can give rise to harmful algal blooms, have been employed to evaluate the antimicroalgal (algicidal) activity of metabolites from marine-derived Trichoderma. As a result, 113 isolates, including 89 sesquiterpenes, 12 diterpenes, three steroids, eight polyketides, and one peptide, exhibit more or less inhibition of the phytoplankton tested (Tables 1, 2, 3). The half maximal inhibitory concentrations (IC50) of 109 metabolites and two ferric complexes are shown in Table 4, and the remaining four ones (162, 163, 165, and 178) display no more than 85% inhibitory rates against A. carterae, C. marina, H. akashiwo, P. donghaiense, P. globosa, and/or S. trochoidea at a concentration of 80 or 100 μg/mL [63–65, 68]. It is worth mentioning that 19 molecules, including 15, 32, 65, 68, 69, 93–95, 104, 106, 107, 114, 115, 129, 149, 167, and 278–280, that account for 16.8% of all the active ones, possess excellent inhibition effects on one or more microalga species, with IC50 values being ≤ 1.0 μg/mL. The former 15 members are involved in five types of terpenes, inclusive of cyclonerane, bisabolane, carotane, cadinane, and harziane derivatives.
Antimicroalgal activity of 109 metabolites from the marine-derived Trichoderma
Structure–activity relationship analyses of cyclonerane sesquiterpenes reveal that the side chain and cyclopentane ring of 15 (IC50 0.66 μg/mL) play important roles in antagonism against C. marina. Although the activity of 33 (IC50 1.1 μg/mL) against C. marina also guarantees this deduction, the nitrogenous unit at the side chain terminus slightly reduces the inhibition effect. In contrast with 6 (IC50 13 μg/mL), the high inhibition ability of 32 (IC50 0.63 μg/mL) against H. akashiwo undoubtedly arises from the nitrogenous side chain at C-10. The enhanced inhibition against H. akashiwo due to the nitrogenous unit is also reflected by comparison of activities between 15 (IC50 23 μg/mL) and 33 (IC50 10 μg/mL). On the other hand, hydroxamic acids 28 and 29 do not feature attractive antimicroalgal activity, but their ferric complexes, Fe(28)3 and Fe(29)3, possess really more inhibition than themselves [27]. It is interesting that these two ferric complexes have no lethality to the marine zooplankton Artemia salina that has often been employed for screeing toxins. In view of the high and broad-spectrum activity and low toxicity of Fe(28)3, it is promising to be applied as a potent inhibitor to control harmful algal blooms in mariculture.
Among bisabolanes and norbisabolanes, only 65, 68, and 69 exhibit IC50 values being ≤ 1.0 μg/mL in the antagonistic assay against microalgae. Their high effects may correlate with the presence of a pyran unit, but it is not the only reason by consideration of the activities of its analogs, such as 54–56 and 62–70. Stereochemistry of the pyran unit and substitution of the side chain and cyclohexane ring also influence the antagonism. Compounds 65 and 68 have the same relative configurations at C-9 and C-11, which may contribute to their high activities against C. marina. As in 69, the variation of relative configurations at C-9 and C-11 decreases the effect on C. marina, but increases the ability to inhibit H. akashiwo. Unfortunately, the aminoglycoside unit in 54–56 fails to effectively improve the antimicroalgal activity.
Carotane sesquiterpenes 93–95 have growth inhibition against C. marina, with IC50 values ranging from 0.24 to 0.33 µg/mL. The carbonyl group at C-3, hydroxy group at C-11, and epoxy group at the seven-membered ring have been indicated to be the key functional groups [44]. Of the 16 cadinanes, five members (104, 106, 107, 114, and 115) possess inhibition against C. marina, H. akashiwo, and/or P. donghaiense, with IC50 values of 0.22–0.96 μg/mL. Similar to 95, both 104 and 106 feature an isopropanol unit that likely contributes to the high activities of these two molecules. On the other hand, the methyl ester group attached to a double bond or an aromatic ring may enhance the ability to inhibit H. akashiwo and P. donghaiense by consideration of the high activities of 106, 107, 114, and 115. As for the diterpenes, harzianes 129 and 149 are more inhibitory against K. veneficum and H. akashiwo, respectively, than the others tested. The double bond at C-9 seems helpful for their inhibition, but the hydroxy group at C-3 has obvious negative effect.
Among three ergosterol derivatives, the highly transformed steroid 167 features high inhibition against C. marina, H. akashiwo, K. veneficum, and P. donghaiense, with IC50 values ranging from 0.27–0.59 μg/mL [66]. Despite the significant activities of this molecule against all the microalgae tested, its structure–activity relationship fails to be speculated due to the lack of any analog. The broad-spectrum inhibition ability of this steroid derivative suggests its application potential in suppressing algal blooms, which will depend on more research on mechanism exploration and toxicity evaluation. Relative to this broad-spectrum algicide, polyketides 278–280 have inhibition effects on only two, C. marina and K. veneficum, of the four microalgae tested [103]. Their slight differences in activity suggest the configurations of two chiral carbons that link a hydroxy group and a methoxy group, respectively, to be weak to influence the algicidal ability.
Antimicroalgal mechanisms of 68 and 104 against C. marina have been preliminarily demonstrated by detecting superoxide dismutase (SOD) and malondialdehyde (MDA) as well as soluble protein (SP) under the influence of these two metabolites, respectively [34]. Both 68 and 104 can effectively decrease the SOD activity and increase MDA and SP levels of C. marina, intimating the damage to antioxidant and membrane systems.
3.2 Zooplankton-toxic
Besides phytoplankton, zooplankton are also widespread in the marine ecosystem and play indispensable roles in the global fishery. A low number of metabolites, 3–5, 38, 53, 57, 61, 93, 98, 101, 128, 129, 143, 149, 162, and 165, have been reported to be toxic to the zooplankton Artemia salina, a brine shrimp species. All these compounds belong to the terpene family, and their toxicities seem not very strong. As metabolites of T. asperellum, 3–5, 38, 53, 128, and 143 have inhibitory rates of 52.2–78.7% at 100 μg/mL, while 57 and 61 possess half maximal lethal concentration (LC50) values of 48 and 62 μg/mL, respectively [26, 36]. As metabolites of T. virens, sesquiterpenes 93, 98, and 101 are toxic to this zooplankton with LC50 values of 66, 56, and 21 μg/mL, repectively [44]. As metabolites of T. longibrachiatum, diterpenes 129 and 149 have LC50 values of 23.1 and 19 μg/mL, respectively [53, 54]. In addition, 162 from T. citrinoviride and 165 from T. harzianum also possess lethal effects on A. salina, with LC50 values of 65.6 and 112 μg/mL, respectively [63, 65]. Apart from these hypotoxic molecules, there are also many nontoxic metabolites in the above or other Trichoderma species of marine origin [27, 44, 103]. In a whole, toxicities to zooplankton are not comparable to antimicroalgal activities for the metabolites from marine-derived Trichoderma.
3.3 Antibacterial
Regardless of unidentified strains, more than 20 Gram-positive and Gram-negative bacterial species, such as Acinetobacter baumannii, Aeromonas hydrophilia, Bacillus subtilis, Enterococcus faecalis, E. faecium, Escherichia coli, Helicobacter pylori, Klebsiella pneumonia, Mycobacterium smegmatis, M. bovis, M. tuberculosis, Photobacterium angustum, Pseudoalteromonas citrea, Pseudomonas aeruginosa, Staphylococcus aureus, S. epidermidis, Vibrio anguillarum, V. harveyi, V. parahaemolyticus, V. splendidus, methicillin-resistant S. aureus (MRSA), vancomycin-resistant E. faecalis (VRE), and vancomycin-resistant E. faecium, were used for antibacterial evaluation. Most of them are pathogens for human beings and aquacultural organisms. A total of 78 metabolites, comprising 47 terpenes, three steroids, 11 polyketides, 14 peptides, one alkaloid, and two others, feature bacteriostatic activities (Tables 1, 2, 3). However, most of these compounds have only moderate to weak inhibition of one or more bacterial species.
If a 10 μg/mL minimum inhibitory concentration (MIC) is taken as the threshold [184], only nine metabolites are regarded as active agents. Sesquiterpene 119 suppresses the aquatic pathogen V. harveyi with an MIC value of 4 μg/mL, while sorbicillinoids 213 and 214 inhibit A. hydrophilia at the same level [49]. Sorbicillinoid derivative 231 has been detected to inhibit S. aureus, VRE, P. aeruginosa, and K. pneumonia, with MIC values of 3.32, 1.63, 6.65, and 6.65 μg/mL, respectively [84]. During the same detection, 295 possesses an MIC value of 12.9 μg/mL to suppress B. subtilis and VRE, suggesting the γ-lactone moiety of 231 to be the key pharmacophore. Chromones 290 and 291 inhibit four H. pylori strains with MIC values of 2–8 μg/mL, and the former also suppresses four S. aureus strains, including MRSA, and one E. faecalis strain as well as one E. faecium strain within the same MIC range [94]. However, these two chromone derivatives have no activity against the Gram-negative E. coli and P. aeruginosa at 64 μg/mL. Aminolipopeptides 311–313 have MIC values of 0.02–2.0 μg/mL against M. smegmatis, M. bovis, and M. tuberculosis under both aerobic and hypoxic conditions, and the AHMOD moiety in 311 and 313 has been deduced to play a key role in anti-mycobacterial activity [114].
Peptaibols 386–393 have stronger inhibition of environmental bacteria than of laboratory ones, though the effects are not much attractive [122]. As a gram-negative bacterium, E. coli seems more resistant to these peptaibols than the gram-positive ones [122]. Compounds 162 and 442 also comply with this regular pattern in antagonism to E. coli and S. aureus, but 163 inhibits E. coli rather than S. aureus [63, 64, 77]. During the disk diffusion assays, 43 compounds, including 5, 15–19, 22–24, 27, 38, 39, 49, 52–56, 62–70, 72, 82, 102, 103, 117, 129, 143, 145, 154, 167, 168, 177, 178, 186, 187, and 289, can inhibit the gram-negative Vibrio species with inhibitory zone diameters of 6–11 mm at a concentration of 20, 40, 50, or 100 μg/disk. As an outstanding representative, 177 has also been assayed for antagonistic activity using the double dilution method, but it features only MIC values of 13–50 μg/mL against the four Vibrio species tested [39]. Less than the number of Vibrio antagonists, 23 metabolites, inclusive of 24, 27, 46–49, 63–66, 68–71, 102–104, 129, 166–168, 177, and 289, can suppress the gram-negative P. citrea. Of these, 289 is the most effective one as seen from the disk diffusion test, but it feature only an MIC value of 16 μg/mL in the microdilution detection [33].
3.4 Antifungal
The production of fungicidal antibiotics is one of the key mechanisms for Trichoderma to control phytopathogenic fungi [18]. However, only 25 new metabolites, 77–81, 83–85, 88, 89, 116, 119, 125, 167, 180, 191, 213, 214, 220, 250, 290, 308, 395, 397, and 431, from marine-derived Trichoderma have antifungal activity against plant or human pathogens (Tables 1, 2, 3). Trichothecane sesquiterpenes, with active members accounting for over a half, are protruded in antagonism to yeast-like and filamentous fungi. Of them, 77–81 possess MIC values ranging from 1.6 to 50 μg/mL against Candida albicans and Cryptococcus neoformans, with 77 being the most potent (MIC values 3.1 and 1.6 μg/mL, respectively) [41, 42]. Structure–activity relationship analyses reveal that substituents at C-4 and C-16 greatly influence the fungistatic potency. Additionally, 83–85, 88, and 89 have MIC values of 32 or 64 μg/mL to inhibit Botrytis cinerea, Cochliobolus miyabeanus, Fusarium oxysporum f. sp. cucumerium, F. oxysporum f. sp. niveum, or Phomopsis asparagi [43]. The low MIC values (4.0–16 μg/mL) of trichodermin suggest that deacetylation and glycosylation at C-4 and hydroxylation and acetoxylation at C-16 are detrimental to the antifungal ability.
If a threshold as in antibacterial evaluation is used, most of the other plant-pathogenic antagonists seem not much attractive. Eleven pathogens of the genera Alternaria, Ceratobasidium, Colletotrichum, Curvularia, Fusarium, Penicillium, and Physalospora have been employed to detect the effects of 119, 213, and 214, but only Curvularia spicifera appears sensitive to the former two metabolites with MIC values of 8 μg/mL [49]. 116 suppresses the pathogenic fungi Colletotrichum gloeosporioides and F. oxysporum with MIC values of 50 and 100 μg/mL, respectively, the same as those of triadimefon [48]. 125 inhibits Colletotrichum lagrnarium and C. fragariae with MIC values of 8 and 16 μg/mL, respectively, but it is relatively weak to antagonize B. cinerea from grape (64 μg/mL) and strawberry (32 μg/mL), F. oxysporum f. sp. cucumerinum (128 μg/mL), and F. oxysporum f. sp. lycopersici (256 μg/mL) [51]. 167 possesses inhibition of Glomerella cingulata with an MIC value of 12 μg/mL [66]. 180 has been assayed for inhibitory activity against B. cinerea, Pestallozzia theae, Phytophthora parasitica, Magnaporthe oryzae, and Valsa mali, with the latter two species being the most sensitive to this metabolite (MICs 8.63 and 34.5 μM) [70]. 191 and 220 have an MIC value of 125 μg/disk and a median effective dose (ED50) of 9.13 μg/mL, respectively, to inhibit Pestalotiopsis theae through the paper disk dilution method, rather than the conventional broth dilution test [76, 82].
Besides trichothecanes, several other metabolites are also antagonistic to human pathogens. Chromone 290 has an MIC value of 16 μg/mL against two C. albicans strains and an MIC value of 64 μg/mL against one Aspergillus fumigatus strain [94]. Dipeptide 308 inhibits amphoterocin-resistant C. albicans with an MIC value of ca. 15 μg/mL, with the chlorine atom being a pharmacophore [112]. Among polypeptides, peptaibol 395 possesses only 63% inhibitory rate against A. fumigatus, a human-pathogenic fungus, at 100 μg/mL [123], while 397 inhibits C. albicans ATCC 10231 from bronchomycosis, 247 FN from finger nails, 311 FN from finger nails, and 098 VC from vaginal cavity with MIC values of 2.49, 4.92, 19.66, and 2.49 μM [124]. Alkaloid 431 suppresses human-pathogenic C. albicans with an IC50 value of 25.38 μM, and ribosylation of the phenolic hydroxy group dramatically reduces the effect as indicated by the low activity of 432. Although 250 possesses no antagonistic effect on C. albicans, it features synergistic antifungal activity with ketoconazole [91]. It must be said that these tests are valuable tries to use Trichoderma metabolites to antagonize human-pathogenic fungi.
3.5 Cytotoxic
A series of human and murine cancer cell lines, such as A-375, A375-S2, A549, A2780, Bel-7402, CNE1, CNE2, DU-145, HCT-8, HCT-15, HCT-116, Hela, HepG2, HL-60, Huh-7, Jurkat, KB, Lovo, MCF-7, MDA-MB-435 s, NCIH-460, NCI-H929, SUNE1, SW620, P388, and L1210 cells, have been used for in vitro cytostatic evaluations, and most of them have been performed through the 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) method. 42 metabolites, including 10 terpenes, 26 polyketides, four peptides, and two alkaloids, have been determined to possess more or less cytotoxicities (Tables 1, 2, 3). If an IC50 value of 10 μM is taken as the potency threshold [184], only 12 metabolites have potent activities.
Among these potent molecules, cadinane sesquiterpene 109 possesses cytotoxicities against NCI-H929 myeloma and SW620 colorectal cancer cells with IC50 values of ca. 6.8 and 9.3 μM, respectively, while 110 inhibits NCIH-460 lung and SW620 colorectal cancer cell lines with IC50 values of ca. 7.8 and 8.6 μM, respectively. The presence of two carboxylic acid groups in 109 and 110 has been deduced to be helpful to their activities [47]. Wickerane diterpenes 156–161 have been assayed for cytotoxicities against HL-60, P388, and L1210 leukemia cells, but only the carbonyl-bearing 156 with IC50 values of 6.8–7.9 μM is potent [61, 62]. Cyclopentenones 182–184 significantly inhibit P388 leukemia cells with IC50 values of 1.69, 6.85, and 9.03 μM, respectively [71]. Bisorbicillinoid 218 features inhibition of human A549 lung and MCF-7 breast cancer cells with IC50 values of 5.1 and 9.5 μM, respectively [86]. Decalin derivatives 265 and 266 suppress HL-60, P388, and L1210 leukemia cells with IC50 values ranging from 3.8–6.7 μM [97], four orders of magnitude lower than that of 261 [95]. Dipeptides 308 and 310 are potential inhibitors for HCT-116 and Jurkat cells, with IC50 values of 0.71 and 1.3 μM, respectively. The chlorine atom in 308 and the disulfide bridge in 310 play key roles in their cytotoxicities [112, 113]. Dipeptide 308 can also inhibit the proliferation of Hela cells, with an IC50 value of 3.1 μM. Besides differences in potency, 308 also has distinct action mechanism, accumulating cells in the S phase, from several synthetic analogs. Its mechanistic specificity is probably attributed to the chlorohydrin moiety, which may contribute to developing DNA-damaging agents. However, numerous in vivo antitumor experiments are required before evaluating the therapeutic potential of this molecule [172]. In addition, peptaibol 395 has an IC50 value of 2.5 μM in inhibition of KB cells [123]. These positive results further strengthen the clinical potential of Trichoderma metabolites.
3.6 Anti-inflammatory
The anti-inflammatory property of some metabolites has been assayed for inhibiting the production of nitric oxide (NO) in different cell lines induced by lipopolysaccharide [40, 55, 81, 88]. The number of active metabolites amounts to 21, and all of them belong to terpene and polyketide families. Trichothecane sesquiterpenes 73–76 reduce the lipopolysaccharide-induced NO production in microglial BV-2 cells at 10 μM, with 75 being the most potent [40]. Harziane diterpenes 132 and 133 weakly suppress the production of NO in RAW264.7 macrophages at 100 μM [55]. Sorbicillinoids 200, 201, 203–206, 208, 211, and 223–228 feature higher anti-inflammatory effects than indomethacin, a positive control with an IC50 value of 41 μM. In particular, the IC50 values of 200, 203–206, 223, and 224 are lower than 10 μM. Monomeric sorbicillinoid 205 with an IC50 value of 0.94 μM is the most potent one, and substitution modes at the phenyl core and the side chain contribute to its high activity [81]. This molecule is promising to be developed as a therapeutic agent after a series of in vitro and in vivo determination in the future. It can be concluded that substitution and oxidation of the benzene ring and side chain have important impacts on anti-inflammatory activities of this type of molecules. In a dose-dependent manner across 3–60 μM, 236 can reduce the nitrite production stimulated by lipopolysaccharide in J774A.1 macrophages [88].
3.7 Miscellaneous
There are 25 metabolites that possess other activities, such as antiviral, phytotoxic, insect-toxic, zebrafish-toxic, antifouling, antioxidant, enzyme-inhibitory, NF-κB-inhibitory, anti-pulmonary fibrosis, anti-Aβ fibrillization, and neuroprotective properties. Sesquiterpenes 126 and 127 have moderate activities against the hepatitis C virus, with half maximal effective concentration (EC50) values being ca. 24.5 and 20.4 μM, respectively [52]. Harziane diterpenes 138–140, 142, 148, and 151 exert growth inhibition of amaranth and lettuce seedlings at 200 ppm. Although they also have an inhibitory effect on amaranth roots, all of them are avirulent to the elongation of lettuce roots [57]. Besides antimicrobial and cytotoxic activities, the acute toxicity to the larvae of Calliphora vomitoria has also been detected for 395, with the minimal effective dose (MED) being 250 μg/mg [123]. Despite the potent inhibition of P. theae by 220, its high effects on mortality and malformation of zebrafish at a low concentration (0.625 μM) exclude the antifungal application [82]. Decalin derivative 268 prevents the settlement of Bugula neritina, a widespread fouling organism in marine environments, larvae with an EC50 value of 29.8 μg/mL [98]. Six metabolites, 196, 234–236, 442, and 444, have been evaluated to be moderate antioxidants through the 1,1-diphenyl-2-picrylhydrazyl (DPPH) radical scavenging method [77, 78, 87, 129]. Of them, 196 is also an effective inhibitor of the Aβ fibril formation and a moderate protectant against the hydrogen peroxide-induced neuronal death [78]. Koninginins 254 and 255 have moderate inhibition of protein tyrosine phosphatase 1B (PTP1B), with IC50 values of 53.1 and 65.1 μM, respectively [92]. In addition, naphthopyrones 276 and 277 possess IC50 values of 58.7 and 18.0 μΜ, respectively, against tyrosine kinase, with the bisnaphtho-γ-pyrone nucleus being responsible for the inhibition [102]. Vertinolide 230 has an IC50 value of 13.83 μM to inhibit TNF-α-induced NF-κB, and no cytotoxicity against the human embryonic kidney cell line 293 has been detected at 50 μM [83]. Alkaloids 427, 429, and 430 have been screened for inhibition of collagen accumulation, and only 427 with a ca. 85.21% inhibitory rate at 10 μM features high potency to reduce the pulmonary fibrosis [125].
4 Metabolic relevance
Trichoderma species have their own metabolic characteristics, reflected by the diverse cycloneranes, bisabolanes, harzianes, sorbicillinoids, and peptaibols. These metabolites may exist in a broad spectrum of species, as shown in Tables 1, 2, 3. On the other hand, it is easy to be understood that different Trichoderma species of marine origin also produce varied metabolites. For examples, carotanes, trichothecanes, wickeranes, and decalins have been encountered in one or a small number of species, demonstrating the interspecific differences of metabolites. Moreover, Trichoderma strains of the same species tend to feature discrepancies in metabolic profiles, evidenced by the discovery of new compounds from each isolate. This phenomenon can be attributed to the variation of strain sources, though technical divergence and personal tendency during isolation and identification processes also make differences to some extent.
The findings of more than 400 new compounds from marine-derived Trichoderma commendably explain the shaping effect of marine environments on biosynthetic routes. The impacts may be caused by genic or transcriptional variations, especially for the known species that have also been reported as terrestrial-derived ones. To date, several new or known skeletons, such as cuparane, synderane, pupukeanane, harzianoic acid, citrinovirin, fusicoccane, cleistanthane, and trichorenin, have been discovered from only marine-derived strains, but their uniqueness still requires more supports from both natural product chemistry and molecular biology. As for substitution, the aminosugar unit has exclusively been found in the metabolites, such as 38, 53–56, 90, 119, and 124, of marine-derived Trichoderma so far. Moreover, marine-derived Trichoderma strains have contributed a series of halogenated metabolites, including 82, 114, 164, 183, 184, 187, 188, 296, 297, 300, 308, and 433, which exhibit higher structural diversity than those from terrestrial-derived strains [20]. Halogenation is characteristic of marine natural products, and the chlorine, bromine, and iodine atoms in these metabolites come from the seawater constituents or the added sodium halides [107, 112].
Culture substrates and conditions play important roles in the secondary metabolite production of marine-derived Trichoderma [185]. Algicolous Trichoderma sp. TPU199 was chemically investigated on the basis of several different cultivations, leading to the discovery of 77–81, 296–300, and 310 as well as several known terpenes, peptides, and polyketides [41, 42, 107, 108, 113]. Fermented in the freshwater medium under agitation for 7 days, this strain produced trichothecane sesquiterpenes 77, 80, and 81 [42]. Supplemented with 3.0% NaI, the culture broth further afforded trichothecane sesquiterpenes 78 and 79 and diketopiperazines 297–299, besides 77 [41]. If only NaI was replaced by NaBr, a brominated diketopiperazine (296) was then yielded [107]. In the same freshwater medium, a long-term (5 weeks) static fermentation induced the production of dipeptide 310 [113]. Agitated for 7 days in the seawater medium, the culture broth gave dipeptide 308 and diketopiperazine DC1149B, a chlorinated analog of 296 and 297 [107]. When 1% dimethyl sulfoxide (DMSO) was added into the seawater, rather than freshwater, medium, the production of diketopiperazine 300 was further induced under agitation for 7 days [108].
"One strain many compounds" (OSMAC) and chemical epigenetic manipulation strategies were also used to the coral-derived T. harzianum XS-20090075 [186], which resulted in the discovery of 12 new molecules [56, 57, 98, 100]. A culture for four weeks in the rice medium led to the production of harziane diterpenes 138–142, 148, and 151 [57]. When the culture period was elongated to 45 days, decalin derivatives 267 and 268 were produced in the same medium [98]. Treated with a histone deacetylase inhibitor (sodium butyrate), two new diterpenes (137 and 164) including a chlorinated one (164) were obtained from the culture in the rice medium [56]. Changed to the Czapek’s medium, the culture afforded a alkaloid (433) that contains both bromine and chlorine atoms [98]. In the PDA (possibly an error of PDB) medium, a static culture for 45 days yielded hydroxyanthraquinones 271 and 272 [100].
Besides the whole substrate variation, a single nutritional factor may also alter the metabolic profile of a Trichoderma strain. Addition of a single amino acid, L-phenylalanine, into the culture medium of T. erinaceum F1-1 elicited the production of 18 aromatic metabolites, with 283–288 being new ones. Simultaneously, an excess (2 g/L) of L-phenylalanine also resulted in the morphological alteration of this fungal strain. However, it was strange that only one of the divergent molecules harbors a nitrogen atom, differing from the privileged production of alkaloids in other marine fungi. The result indicated the incapability of some Trichoderma strains to incorporate a single amino acid into metabolic pathways [32].
5 Co-culture induction
Trichoderma species are always present in extremely complex microbial communities. Apart from abiotic conditions, other microbes also interfere their growth and metabolism in nature. Undoubtedly, co-culture experiments are effective approaches to simulate the actual ecological situation and awake some sleeping genes. Mixed fermentation of marine-derived Trichoderma strains with other fungal and bacterial strains led to the production of nine new metabolites, 446–454 (Fig. 37) [187–190]. Chaunopyran A (446) was produced at an extremely low level by monoculture of Chaunopycnis sp. CMB-MF028, but its productivity was greatly improved by co-cultivation with T. hamatum CMB-MF030. Both the fungal isolates were obtained from the inner tissue of the mollusc Siphonaria sp [187]. (Z)-2-Ethylhex-2-enedioic acid (447) and (E)-4-oxo-2-propylideneoct-7-enoic acid (448) were yielded by co-culture of Trichoderma sp. Gc(M2) and Penicillium sp. Ma(M3)V, isolated from different marine sponge species [188]. Two new sesquiterpenes (449 and 450) and four new polyketides (451–454) were identified from co-culture extracts of fungus Trichoderma sp. 307 and bacterium Acinetobacter johnsonii B2, obtained from a mangrove plant and an aquaculture pond, respectively [189, 190]. It can be concluded that Trichoderma species or other microbes might produce extra products due to the activation of silent metabolic routes during co-culture processes [191]. At least 75 Trichoderma species have the mycoparasitic ability [7], and their fermentation mixed with other fungi means more intensive competition or antagonism that may involve the large-scale production of antibiotics.
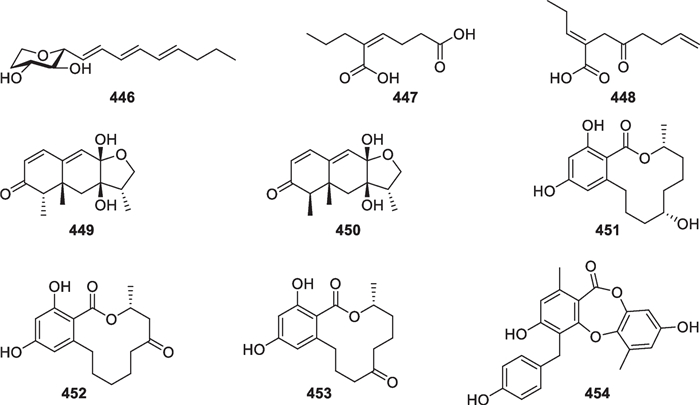
Metabolites from co-cultures of marine-derived Trichoderma and other microbial strains
6 Biotransformation and biodegradation
Trichoderma species exhibit biotransformation functions towards heterogeneous organic matters due to the production of enriched enzymes [192, 193], but marine-derived strains have less been examined. As the metabolites of Chaunopycnis sp., pyridoxatin atropisomers were transformed to a pair of methylated derivatives by the co-cultivated T. hamatum [187]. As a biogenetic precursor of many monoterpenes, geraniol was transformed to 1,7-dihydroxy-3,7-dimethyl-(E)-oct-2-ene by a marine-derived Hypocrea strain. During the process, a double bond was oxygenated to an alcohol [194]. On the other hand, three marine-derived Trichoderma strains effectively degraded malachite green, pentachlorophenol, and chlorpyrifos, respectively, which are industrial and agricultural agents causing environmental pollution [195–197]. These strains were obtained from mangrove, ascidian, and sponge organisms, respectively, and their degradation ability may rely on the production of exoenzymes, such as laccase and peroxidase [195, 198]. Considering the diverse substitution patterns in metabolites from marine-derived Trichoderma, their biotransformation and biodegradation potentials will not disappoint the relevant researchers in future.
7 Conclusions
Over a duration of 30 years, marine-sourced 77 Trichoderma strains have contributed 445 new metabolites, including many new structural types from this genus and new carbon skeletons in nature. Their metabolic characteristics are also reflected by the high populations of irregular carbon numbers and cyclization and cleavage patterns in some scaffolds. These phenomena might arise from the oxidation by reactive oxygen species produced under saline and alkaline stresses. Halogen and sulfur atoms as well as sugar units are incorporated into 12, nine, and 15 members, respectively, especially eight aminosugar-bearing ones. The production of halides probably profits from enriched halogen anions and related enzymes. The discovery greatly adds to the molecular diversity of Trichoderma metabolites, especially those with cyclonerane, bisabolane, harziane, sorbicillinoid, and peptaibol frameworks. Among the metabolites, 235 members feature antimicroalgal, zooplankton-toxic, antibacterial, antifungal, cytotoxic, anti-inflammatory, and other activities, demonstrating the application potentials of marine-derived Trichoderma in aquaculture, agriculture, and healthcare. Marine environments probably shape the metabolic profiles of the involved Trichoderma strains, and culture methods seem also responsible for the metabolic diversity. Considering the mycoparasitic instinct of many Trichoderma species, co-culture with other fungi seems a prospective strategy to mine their valuable metabolites. On the other hand, marine-derived Trichoderma strains may methylate and oxidize heterogeneous molecules, and their biodegradation ability is also attractive. In view of the high structural diversity and various bioactivities, the molecules produced or transformed by marine-derived Trichoderma are promising candidates to develop potent drugs for human diseases and antagonistic agents in agriculture and aquaculture.
Notes
Acknowledgements
We acknowledge the authors cited in references for their contributions to the research on metabolites from marine-derived Trichoderma.
Author contributions
YPS peformed data collection and analysis and wrote the draft. NYJ designed and revised the manuscript. All authors have read and approved the manuscript.
Funding
The preparation of this review and our related research were financially supported by the National Natural Science Foundation of China (42076096 and 31670355), the Taishan Scholar Project Special Funding (tsqn201909164), and the Youth Innovation Promotion Association of the CAS (2023222).
Availability of data and materials
All the data and materials provided in the manuscript are obtained from included references and available upon request.
Declarations
Ethics approval and consent to participate
Ethical declaration is not applicable for this review.
Competing interests
The authors declare that there are no competing interests associated with this work.
References
-
1.Persoon CH. Disposita methodica fungorum. Römer’s Neues Mag Bot. 1794;1: 81-128. PubMed Google Scholar
-
2.Kirk PM, Cannon PF, Minter DW, Stalpers JA. Anisworth & Bisby’s Dictionary of the Fungi (10th edition). Wallingford: CAB International Publishing; 2008. p. 1–771. PubMed Google Scholar
-
3.Harman GE, Howell CR, Viterbo A, Chet I, Lorito M. Trichoderma species—opportunistic, avirulent plant symbionts. Nat Rev Microbiol. 2004;2: 43-56. CrossRef PubMed Google Scholar
-
4.Samuels GJ. Trichoderma: systematics, the sexual state, and ecology. Phytopathology. 2006;96: 195-206. CrossRef PubMed Google Scholar
-
5.Rossman AY, Seifert KA, Samuels GJ, Minnis AM, Schroers H-J, Lombard L, Crous PW, Põldmaa K, Cannon PF, Summerbell RC, Geiser DM, Zhuang W-Y, Hirooka Y, Herrera C, Salgado-Salazar C, Chaverri P. Genera in Bionectriaceae, Hypocreaceae, and Nectriaceae (Hypocreales) proposed for acceptance or rejection. IMA Fungus. 2013;4: 41-51. CrossRef PubMed Google Scholar
-
6.Bissett J, Gams W, Jaklitsch W, Samuels GJ. Accepted Trichoderma names in the year 2015. IMA Fungus. 2015;6: 263-95. CrossRef PubMed Google Scholar
-
7.Contreras-Cornejo HA, Macías-Rodríguez L, del-Val E, Larsen J. Ecological functions of Trichoderma spp. and their secondary metabolites in the rhizosphere: interactions with plants. FEMS Microbiol Ecol. 2016;92: 036. CrossRef PubMed Google Scholar
-
8.Chen K, Zhuang W-Y. Trichoderma shennongjianum and Trichoderma tibetense, two new soil-inhabiting species in the Strictipile clade. Mycoscience. 2016;57: 311-9. CrossRef PubMed Google Scholar
-
9.Qin W-T, Zhuang W-Y. Seven wood-inhabiting new species of the genus Trichoderma (Fungi, Ascomycota) in Viride clade. Sci Rep. 2016;6: 27074. CrossRef PubMed Google Scholar
-
10.Chen K, Zhuang W-Y. Discovery from a large-scaled survey of Trichoderma in soil of China. Sci Rep. 2017;7: 9090. CrossRef PubMed Google Scholar
-
11.Zhu Z-X, Xu H-X, Zhuang W-Y, Li Y. Two new green-spored species of Trichoderma (Sordariomycetes, Ascomycota) and their phylogenetic positions. MycoKeys. 2017;26: 61-75. CrossRef PubMed Google Scholar
-
12.Weindling R. Trichoderma lignorum as a parasite of other soil fungi. Phytopathology. 1932;22: 837-45. PubMed Google Scholar
-
13.Weindling R. Studies on a lethal principle effective in the parasitic action of Trichoderma lignorum on Rhizoctonia solani and other soil fungi. Phytopathology. 1934;24: 1153-79. PubMed Google Scholar
-
14.Woo SL, Ruocco M, Vinale F, Nigro M, Marra R, Lombardi N, Pascale A, Lanzuise S, Manganiello G, Lorito M. Trichoderma-based products and their widespread use in agriculture. Open Mycol J. 2014;8: 71-126. CrossRef PubMed Google Scholar
-
15.Singh A, Shukla N, Kabadwal BC, Tewari AK, Kumar J. Review on plant-Trichoderma-pathogen interaction. Int J Curr Microbiol App Sci. 2018;7: 2382-97. CrossRef PubMed Google Scholar
-
16.Vinale F, Sivasithamparam K, Ghisalberti EL, Marra R, Woo SL, Lorito M. Trichoderma–plant–pathogen interactions. Soil Biol Biochem. 2008;40: 1-10. CrossRef PubMed Google Scholar
-
17.Howell CR. Mechanisms employed by Trichoderma species in the biological control of plant diseases: the history and evolution of current concepts. Plant Dis. 2003;87: 4-10. CrossRef PubMed Google Scholar
-
18.Ghisalberti EL, Sivasithamparam K. Antifungal antibiotics produced by Trichoderma spp. Soil Biol Biochem. 1991;23: 1011-20. CrossRef PubMed Google Scholar
-
19.Keswani C, Mishra S, Sarma BK, Singh SP, Singh HB. Unraveling the efficient applications of secondary metabolites of various Trichoderma spp. Appl Microbiol Biotechnol. 2014;98: 533-44. CrossRef PubMed Google Scholar
-
20.Reino JL, Guerrero RF, Hernández-Galán R, Collado IG. Secondary metabolites from species of the biocontrol agent Trichoderma. Phytochem Rev. 2008;7: 89-123. CrossRef PubMed Google Scholar
-
21.Li M-F, Li G-H, Zhang K-Q. Non-volatile metabolites from Trichoderma spp. Metabolites. 2019;9: 58. CrossRef PubMed Google Scholar
-
22.Bai B, Liu C, Zhang C, He X, Wang H, Peng W, Zheng C. Trichoderma species from plant and soil: an excellent resource for biosynthesis of terpenoids with versatile bioactivities. J Adv Res. 2023;49: 81-102. CrossRef PubMed Google Scholar
-
23.Su D, Ding L, He S. Marine-derived Trichoderma species as a promising source of bioactive secondary metabolites. Mini-Rev Med Chem. 2018;18: 1702-13. CrossRef PubMed Google Scholar
-
24.Kobayashi M, Uehara H, Matsunami K, Aoki S, Kitagawa I. Trichoharzin, a new polyketide produced by the imperfect fungus Trichoderma harzianum separated from the marine sponge Micale cecilia. Tetrahedron Lett. 1993;34: 7925-8. CrossRef PubMed Google Scholar
-
25.Song Y-P, Miao F-P, Fang S-T, Yin X-L, Ji N-Y. Halogenated and nonhalogenated metabolites from the marine-alga-endophytic fungus Trichoderma asperellum cf44-2. Mar Drugs. 2018;16: 266. CrossRef PubMed Google Scholar
-
26.Song Y-P, Liu X-H, Shi Z-Z, Miao F-P, Fang S-T, Ji N-Y. Bisabolane, cyclonerane, and harziane derivatives from the marine-alga-endophytic fungus Trichoderma asperellum cf44-2. Phytochemistry. 2018;152: 45-52. CrossRef PubMed Google Scholar
-
27.Song Y-P, Miao F-P, Yin X-L, Ji N-Y. Nitrogenous cyclonerane sesquiterpenes from an algicolous strain of Trichoderma asperellum. Org Chem Front. 2019;6: 3698-704. CrossRef PubMed Google Scholar
-
28.Song Y-P, Miao F-P, Liu X-H, Yin X-L, Ji N-Y. Cyclonerane derivatives from the algicolous endophytic fungus Trichoderma asperellum A-YMD-9-2. Mar Drugs. 2019;17: 252. CrossRef PubMed Google Scholar
-
29.Fang S-T, Wang Y-J, Ma X-Y, Yin X-L, Ji N-Y. Two new sesquiterpenoids from the marine-sediment-derived fungus Trichoderma harzianum P1–4. Nat Prod Res. 2019;33: 3127-33. CrossRef PubMed Google Scholar
-
30.Song Y-P, Fang S-T, Miao F-P, Yin X-L, Ji N-Y. Diterpenes and sesquiterpenes from the marine algicolous fungus Trichoderma harzianum X-5. J Nat Prod. 2018;81: 2553-9. CrossRef PubMed Google Scholar
-
31.Ma X-Y, Song Y-P, Shi Z-Z, Ji N-Y. Three sesquiterpenes from the marine-alga-epiphytic fungus Trichoderma hamatum Z36–7. Phytochem Lett. 2021;43: 98-102. CrossRef PubMed Google Scholar
-
32.Guo Y-W, Gong B-Q, Yuan J, Li H-J, Mahmud T, Huang Y, Li J-F, Yang D-P, Lan W-J. l-Phenylalanine alters the privileged secondary metabolite production in the marine-derived fungus Trichoderma erinaceum F1–1. J Nat Prod. 2020;83: 79-87. CrossRef PubMed Google Scholar
-
33.Liu X-H, Hou X-L, Song Y-P, Wang B-G, Ji N-Y. Cyclonerane sesquiterpenes and an isocoumarin derivative from the marine-alga-endophytic fungus Trichoderma citrinoviride A-WH-20-3. Fitoterapia. 2020;141: 104469. CrossRef PubMed Google Scholar
-
34.Zou J-X, Song Y-P, Liu X-H, Li X-N, Ji N-Y. Bisabolane, cadinane, and cyclonerane sesquiterpenes from an algicolous strain of Trichoderma asperelloides. Bioorg Chem. 2021;115: 105223. CrossRef PubMed Google Scholar
-
35.Song Y, Miao F, Yin X, Ji N. Three nitrogen-containing metabolites from an algicolous isolate of Trichoderma asperellum. Mar Life Sci Technol. 2020;2: 155-60. CrossRef PubMed Google Scholar
-
36.Shi Z-Z, Miao F-P, Fang S-T, Yin X-L, Ji N-Y. Trichobisabolins A-H, eight new bisabolane derivatives from the marine-alga-epiphytic fungus Trichoderma asperellum Y6–2. Fitoterapia. 2019;134: 372-7. CrossRef PubMed Google Scholar
-
37.Shi Z-Z, Liu X-H, Song Y-P, Yin X-L, Ji N-Y. Sesquiterpenoids and a steroid from the algicolous Trichoderma brevicompactum. Fitoterapia. 2021;153: 104983. CrossRef PubMed Google Scholar
-
38.Song Y-P, Miao F-P, Liu X-H, Yin X-L, Ji N-Y. Seven chromanoid norbisabolane derivatives from the marine-alga-endophytic fungus Trichoderma asperellum A-YMD-9-2. Fitoterapia. 2019;135: 107-13. CrossRef PubMed Google Scholar
-
39.Liu X-H, Song Y-P, Wang B-G, Ji N-Y. Sesquiterpenes and lipids from the algicolous fungus Trichoderma atroviride RR-dl-3-9. Phytochem Lett. 2021;45: 6-12. CrossRef PubMed Google Scholar
-
40.Safwan S, Wang S-W, Hsiao G, Hsiao S-W, Hsu S-J, Lee T-H, Lee C-K. New trichothecenes isolated from the marine algicolous fungus Trichoderma brevicompactum. Mar Drugs. 2022;20: 80. CrossRef PubMed Google Scholar
-
41.Yamazaki H, Takahashi O, Kirikoshi R, Yagi A, Ogasawara T, Bunya Y, Rotinsulu H, Uchida R, Namikoshi M. Epipolythiodiketopiperazine and trichothecene derivatives from the NaI-containing fermentation of marine-derived Trichoderma cf. brevicompactum. J Antibiot. 2020;73: 559-67. CrossRef PubMed Google Scholar
-
42.Yamazaki H, Yagi A, Takahashi O, Yamaguchi Y, Saito A, Namikoshi M, Uchida R. Antifungal trichothecene sesquiterpenes obtained from the culture broth of marine-derived Trichoderma cf. brevicompactum and their structure–activity relationship. Bioorg Med Chem Lett. 2020;30: 127375. CrossRef PubMed Google Scholar
-
43.Shi Z-Z, Liu X-H, Li X-N, Ji N-Y. Antifungal and antimicroalgal trichothecene sesquiterpenes from the marine algicolous fungus Trichoderma brevicompactum A-DL-9-2. J Agric Food Chem. 2020;68: 15440-8. CrossRef PubMed Google Scholar
-
44.Shi Z-Z, Fang S-T, Miao F-P, Yin X-L, Ji N-Y. Trichocarotins A-H and trichocadinin A, nine sesquiterpenes from the marine-alga-epiphytic fungus Trichoderma virens. Bioorg Chem. 2018;81: 319-25. CrossRef PubMed Google Scholar
-
45.Song Y-P, Shi X-S, Wang B-G, Ji N-Y. Cadinane and carotane derivatives from the marine algicolous fungus Trichoderma virens RR-dl-6-8. Fitoterapia. 2020;146: 104715. CrossRef PubMed Google Scholar
-
46.Song Y-P, Miao F-P, Liang X-R, Yin X-L, Ji N-Y. Harziane and cadinane terpenoids from the alga-endophytic fungus Trichoderma asperellum A-YMD-9-2. Phytochem Lett. 2019;32: 38-41. CrossRef PubMed Google Scholar
-
47.Cui J, Shang R-Y, Sun M, Li Y-X, Liu H-Y, Lin H-W, Jiao W-H. Trichodermaloids A-C, cadinane sesquiterpenes from a marine sponge symbiotic Trichoderma sp. SM16 fungus. Chem Biodivers. 2020;17: e2000036. CrossRef PubMed Google Scholar
-
48.Qin C, Hu Z, Xiong Y, Chen M, Li C, Ding W. A new sesquiterpene derivative from the mangrove endophytic fungus Trichoderma harzianum (strain No. R1). Chem Nat Compd. 2021;57: 312-4. CrossRef PubMed Google Scholar
-
49.Wang Y, Li X-M, Yang S-Q, Zhang F-Z, Wang B-G, Li H-L, Meng L-H. Sesquiterpene and sorbicillinoid glycosides from the endophytic fungus Trichoderma longibrachiatum EN-586 derived from the marine red alga Laurencia obtusa. Mar Drugs. 2022;20: 177. CrossRef PubMed Google Scholar
-
50.Du X-P, Li Y-Y, Lu C-H, Zheng Z-H, Shen Y-M. A novel sesquiterpene glucoside from Trichoderma sp. PT2. Nat Prod Res Dev. 2010;22: 544-7. PubMed Google Scholar
-
51.Du F-Y, Ju G-L, Xiao L, Zhou Y-M, Wu X. Sesquiterpenes and cyclodepsipeptides from marine-derived fungus Trichoderma longibrachiatum and their antagonistic activities against soil-borne pathogens. Mar Drugs. 2020;18: 165. CrossRef PubMed Google Scholar
-
52.Li B, Li L, Peng Z, Liu D, Si L, Wang J, Yuan B, Huang J, Proksch P, Lin W. Harzianoic acids A and B, new natural scaffolds with inhibitory effects against hepatitis C virus. Bioorg Med Chem. 2019;27: 560-7. CrossRef PubMed Google Scholar
-
53.Miao F-P, Liang X-R, Yin X-L, Wang G, Ji N-Y. Absolute configurations of unique harziane diterpenes from Trichoderma species. Org Lett. 2012;14: 3815-7. CrossRef PubMed Google Scholar
-
54.Zou J-X, Song Y-P, Ji N-Y. Deoxytrichodermaerin, a harziane lactone from the marine algicolous fungus Trichoderma longibrachiatum A-WH-20-2. Nat Prod Res. 2021;35: 216-21. CrossRef PubMed Google Scholar
-
55.Li H, Liu X, Li X, Hu Z, Wang L. Novel harziane diterpenes from deep-sea sediment fungus Trichoderma sp. SCSIOW21 and their potential anti-inflammatory effects. Mar Drugs. 2021;19: 689. CrossRef PubMed Google Scholar
-
56.Shi T, Shao C-L, Liu Y, Zhao D-L, Cao F, Fu X-M, Yu J-Y, Wu J-S, Zhang Z-K, Wang C-Y. Terpenoids from the coral-derived fungus Trichoderma harzianum (XS-20090075) induced by chemical epigenetic manipulation. Front Microbiol. 2020;11: 572. CrossRef PubMed Google Scholar
-
57.Zhao D-L, Yang L-J, Shi T, Wang C-Y, Shao C-L, Wang C-Y. Potent phytotoxic harziane diterpenes from a soft coral-derived strain of the fungus Trichoderma harzianum XS-20090075. Sci Rep. 2019;9: 13345. CrossRef PubMed Google Scholar
-
58.Zhang M, Liu J-M, Zhao J-L, Li N, Chen R-D, Xie K-B, Zhang W-J, Feng K-P, Yan Z, Wang N, Dai J-G. Two new diterpenoids from the endophytic fungus Trichoderma sp. Xy24 isolated from mangrove plant Xylocarpus granatum. Chin Chem Lett. 2016;27: 957-60. CrossRef PubMed Google Scholar
-
59.Zou J-X, Song Y-P, Zeng Z-Q, Ji N-Y. Proharziane and harziane derivatives from the marine algicolous fungus Trichoderma asperelloides RR-dl-6-11. J Nat Prod. 2021;84: 1414-9. CrossRef PubMed Google Scholar
-
60.Xie Z-L, Li H-J, Wang L-Y, Liang W-L, Liu W, Lan W-J. Trichodermaerin, a new diterpenoid lactone from the marine fungus Trichoderma erinaceum associated with the sea star Acanthaster planci. Nat Prod Commun. 2013;8: 67-8. PubMed Google Scholar
-
61.Yamada T, Suzue M, Arai T, Kikuchi T, Tanaka R. Trichodermanins C-E, new diterpenes with a fused 6-5-6-6 ring system produced by a marine sponge-derived fungus. Mar Drugs. 2017;15: 169. CrossRef PubMed Google Scholar
-
62.Yamada T, Fujii A, Kikuchi T. New diterpenes with a fused 6-5-6-6 ring system isolated from the marine sponge-derived fungus Trichoderma harzianum. Mar Drugs. 2019;17: 480. CrossRef PubMed Google Scholar
-
63.Liang X-R, Miao F-P, Song Y-P, Liu X-H, Ji N-Y. Citrinovirin with a new norditerpene skeleton from the marine algicolous fungus Trichoderma citrinoviride. Bioorg Med Chem Lett. 2016;26: 5029-31. CrossRef PubMed Google Scholar
-
64.Liang X-R, Miao F-P, Song Y-P, Guo Z-Y, Ji N-Y. Trichocitrin, a new fusicoccane diterpene from the marine brown alga-endophytic fungus Trichoderma citrinoviride cf-27. Nat Prod Res. 2016;30: 1605-10. CrossRef PubMed Google Scholar
-
65.Liang X-R, Ma X-Y, Ji N-Y. Trichosordarin A, a norditerpene glycoside from the marine-derived fungus Trichoderma harzianum R5. Nat Prod Res. 2020;34: 2037-42. CrossRef PubMed Google Scholar
-
66.Song Y-P, Shi Z-Z, Miao F-P, Fang S-T, Yin X-L, Ji N-Y. Tricholumin A, a highly transformed ergosterol derivative from the alga-endophytic fungus Trichoderma asperellum. Org Lett. 2018;20: 6306-9. CrossRef PubMed Google Scholar
-
67.Li B, Huang Q-X, Gao D, Liu D, Ji Y-B, Liu H-G, Lin W-H. New C13 lipids from the marine-derived fungus Trichoderma harzianum. J Asian Nat Prod Res. 2015;17: 468-74. CrossRef PubMed Google Scholar
-
68.Liu X-H, Ji N-Y. Isolation, identification, and bioactivity of a new triol from algicolous fungus Trichoderma citrinoviride. Chem Bioeng. 2022;39: 15-8. PubMed Google Scholar
-
69.Hu X, Gong M-W, Zhang W-W, Zheng Q-H, Liu Q-Y, Chen L, Zhang Q-Q. Novel cytotoxic metabolites from the marine-derived fungus Trichoderma citrinoviride. Heterocycles. 2014;89: 189-96. CrossRef PubMed Google Scholar
-
70.Zhao D-L, Zhang X-F, Huang R-H, Wang D, Wang X-Q, Li Y-Q, Zheng C-J, Zhang P, Zhang C-S. Antifungal nafuredin and epithiodiketopiperazine derivatives from the mangrove-derived fungus Trichoderma harzianum D13. Front Microbiol. 2020;11: 1495. CrossRef PubMed Google Scholar
-
71.Amagata T, Usami Y, Minoura K, Ito T, Numata A. Cytotoxic substances produced by a fungal strain from a sponge: physico-chemical properties and structures. J Antibiot. 1998;51: 33-40. CrossRef PubMed Google Scholar
-
72.Usami Y, Ikura T, Amagata T, Numata A. First total syntheses and configurational assignments of cytotoxic trichodenones A-C. Tetrahedron: Asymm. 2000;11: 3711-25. CrossRef PubMed Google Scholar
-
73.You J, Dai H, Chen Z, Liu G, He Z, Song F, Yang X, Fu H, Zhang L, Chen X. Trichoderone, a novel cytotoxic cyclopentenone and cholesta-7,22-diene-3β, 5α, 6β-triol, with new activities from the marine-derived fungus Trichoderma sp. J Ind Microbiol Biotechnol. 2010;37: 245-52. CrossRef PubMed Google Scholar
-
74.Qiu P, Ding L, Sun D, He S. A new cyclopentenone derivative from the sponge-associated fungus Hypocrea koningii. Chem Nat Compd. 2018;54: 631-3. CrossRef PubMed Google Scholar
-
75.Sun S, Tian L, Wang Y, Wu H, Lu X, Pei Y. A novel natural product from the fermentation liquid of marine fungus Trichoderma atroviride G20–12. Asian J Trad Med. 2009;4: 123-7. PubMed Google Scholar
-
76.Tang X-X, Liu S-Z, Sun Y-Y, He F-M, Xu G-X, Fang M-J, Zhen W, Qiu Y-K. New cyclopentenoneacrylic acid derivatives from a marine-derived fungus Trichoderma atroviride H548. Nat Prod Res. 2021;35: 3772-9. CrossRef PubMed Google Scholar
-
77.Ding L-J, Gu B-B, Jiao W-H, Yuan W, Li Y-X, Tang W-Z, Yu H-B, Liao X-J, Han B-N, Li Z-Y, Xu S-H, Lin H-W. New furan and cyclopentenone derivatives from the sponge-associated fungus Hypocrea Koningii PF04. Mar Drugs. 2015;13: 5579-92. CrossRef PubMed Google Scholar
-
78.Fang F, Zhao J, Ding L, Huang C, Naman B, He S, Wu B, Zhu P, Luo Q, Gerwick WH, Yan X, Wang Q, Zhang Z, Cui W. 5-Hydroxycyclopenicillone, a new β-amyloid fibrillization inhibitor from a sponge-derived fungus Trichoderma sp. HPQJ-34. Mar Drugs. 2017;15: 260. CrossRef PubMed Google Scholar
-
79.Du L, Zhu T, Li L, Cai S, Zhao B, Gu Q. Cytotoxic sorbicillinoids and bisorbicillinoids from a marine-derived fungus Trichoderma sp. Chem Pharm Bull. 2009;57: 220-3. CrossRef PubMed Google Scholar
-
80.Lan W-J, Zhao Y, Xie Z-L, Liang L-Z, Shao W-Y, Zhu L-P, Yang D-P, Zhu X-F, Li H-J. Novel sorbicillin analogues from the marine fungus Trichoderma sp. associated with the seastar Acanthaster planci. Nat Prod Commun. 2012;7: 1337-40. PubMed Google Scholar
-
81.Zhang P, Deng Y, Lin X, Chen B, Li J, Liu H, Chen S, Liu L. Anti-inflammatory mono- and dimeric sorbicillinoids from the marine-derived fungus Trichoderma reesei 4670. J Nat Prod. 2019;82: 947-57. CrossRef PubMed Google Scholar
-
82.Liu S-Z, Xu G-X, He F-M, Zhang W-B, Wu Z, Li M-Y, Tang X-X, Qiu Y-K. New sorbicillinoids with tea pathogenic fungus inhibitory effect from marine-derived fungus Hypocrea jecorina H8. Mar Drugs. 2022;20: 213. CrossRef PubMed Google Scholar
-
83.Zaman KAU, Wu X, Sarotti AM, Cao S. New and bioactive polyketides from Hawaiian marine-derived fungus Trichoderma sp. FM652. Nat Prod Res. 2022;36: 5984-90. CrossRef PubMed Google Scholar
-
84.Meng J, Cheng W, Heydari H, Wang B, Zhu K, Konuklugil B, Lin W. Sorbicillinoid-based metabolites from a sponge-derived fungus Trichoderma saturnisporum. Mar Drugs. 2018;16: 226. CrossRef PubMed Google Scholar
-
85.Sperry S, Samuels GJ, Crews P. Vertinoid polyketides from the saltwater culture of the fungus Trichoderma longibrachiatum separated from a Haliclona marine sponge. J Org Chem. 1998;63: 10011-4. CrossRef PubMed Google Scholar
-
86.Rehman SU, Yang L-J, Zhang Y-H, Wu J-S, Shi T, Haider W, Shao C-L, Wang C-Y. Sorbicillinoid derivatives from sponge-derived fungus Trichoderma reesei (HN-2016-018). Front Microbiol. 2020;11: 1334. CrossRef PubMed Google Scholar
-
87.Neumann K, Abdel-Lateff A, Wright AD, Kehraus S, Krick A, König GM. Novel sorbicillin derivatives with an unprecedented carbon skeleton from the sponge-derived fungus Trichoderma species. Eur J Org Chem. 2007;2007: 2268-75. CrossRef PubMed Google Scholar
-
88.Marra R, Nicoletti R, Pagano E, DellaGreca M, Salvatore MM, Borrelli F, Lombardi N, Vinale F, Woo SL, Andolfi A. Inhibitory effect of trichodermanone C, a sorbicillinoid produced by Trichoderma citrinoviride associated to the green alga Cladophora sp., on nitrite production in LPS-stimulated macrophages. Nat Prod Res. 2019;33: 3389-97. CrossRef PubMed Google Scholar
-
89.Sun Y, Tian L, Huang J, Ma H-Y, Zheng Z, Lv A-L, Yasukawa K, Pei Y-H. Trichodermatides A–D, novel polyketides from the marine-derived fungus Trichoderma reesei. Org Lett. 2008;10: 393-6. CrossRef PubMed Google Scholar
-
90.Wang X-H, Xu X-Y, Li Y, Yao F-H, Chen X, Qi S-H, Tao S-H. Secondary metabolites and antimicrobial activities of Hypocrea lixii SCSIO 41520. Nat Prod Res Dev. 2020;32: 1522-8. PubMed Google Scholar
-
91.Song F, Dai H, Tong Y, Ren B, Chen C, Sun N, Liu X, Bian J, Liu M, Gao H, Liu H, Chen X, Zhang L. Trichodermaketones A–D and 7-O-methylkoninginin D from the marine fungus Trichoderma koningii. J Nat Prod. 2010;73: 806-10. CrossRef PubMed Google Scholar
-
92.Yamazaki H, Saito R, Takahashi O, Kirikoshi R, Toraiwa K, Iwasaki K, Izumikawa Y, Nakayama W, Namikoshi M. Trichoketides A and B, two new protein tyrosine phosphatase 1B inhibitors from the marine-derived fungus Trichoderma sp. J Antibiot. 2015;68: 628-32. CrossRef PubMed Google Scholar
-
93.Nuansri S, Rukachaisirikul V, Rungwirain N, Kaewin S, Yimnual C, Phongpaichit S, Preedanon S, Sakayaroj J, Muanprasat C. α-Pyrone and decalin derivatives from the marine-derived fungus Trichoderma harzianum PSU-MF79. Nat Prod Res. 2022;36: 5462-9. CrossRef PubMed Google Scholar
-
94.Lai C, Chen J, Liu J, Tian D, Lan D, Liu T, Wu B, Bi H, Tang J. New polyketides from a hydrothermal vent sediment fungus Trichoderma sp. JWM29-10-1 and their antimicrobial effects. Mar Drugs. 2022;20: 720. CrossRef PubMed Google Scholar
-
95.Yamada T, Mizutani Y, Umebayashi Y, Inno N, Kawashima M, Kikuchi T, Tanaka R. Tandyukisin, a novel ketoaldehyde decalin derivative, produced by a marine sponge-derived Trichoderma harzianum. Tetrahedron Lett. 2014;55: 662-4. CrossRef PubMed Google Scholar
-
96.Yamada T, Umebayashi Y, Kawashima M, Sugiura Y, Kikuchi T, Tanaka R. Determination of the chemical structures of tandyukisins B–D, isolated from a marine sponge-derived fungus. Mar Drugs. 2015;13: 3231-40. CrossRef PubMed Google Scholar
-
97.Suzue M, Kikuchi T, Tanaka R, Yamada T. Tandyukisins E and F, novel cytotoxic decalin derivatives isolated from a marine sponge-derived fungus. Tetrahedron Lett. 2016;57: 5070-3. CrossRef PubMed Google Scholar
-
98.Yu J-Y, Shi T, Zhou Y, Xu Y, Zhao D-L, Wang C-Y. Naphthalene derivatives and halogenate quinoline from the coral-derived fungus Trichoderma harzianum (XS-20090075) through OSMAC approach. J Asian Nat Prod Res. 2021;23: 250-7. CrossRef PubMed Google Scholar
-
99.Khamthong N, Rukachaisirikul V, Tadpetch K, Kaewpet M, Phongpaichit S, Preedanon S, Sakayaroj J. Tetrahydroanthraquinone and xanthone derivatives from the marine-derived fungus Trichoderma aureoviride PSU-F95. Arch Pharm Res. 2012;35: 461-8. CrossRef PubMed Google Scholar
-
100.Shi T, Hou X-M, Li Z-Y, Cao F, Zhang Y-H, Yu J-Y, Zhao D-L, Shao C-L, Wang C-Y. Harzianumnones A and B: two hydroxyanthraquinones from the coral-derived fungus Trichoderma harzianum. RSC Adv. 2018;8: 27596. CrossRef PubMed Google Scholar
-
101.Pang X, Lin X, Tian Y, Liang R, Wang J, Yang B, Zhou X, Kaliyaperumal K, Luo X, Tu Z, Liu Y. Three new polyketides from the marine sponge-derived fungus Trichoderma sp. SCSIO41004. Nat Prod Res. 2018;32: 105-11. CrossRef PubMed Google Scholar
-
102.Ohkawa Y, Miki K, Suzuki T, Nishio K, Sugita T, Kinoshita K, Takahashi K, Koyama K. Antiangiogenic metabolites from a marine-derived fungus, Hypocrea vinosa. J Nat Prod. 2010;73: 579-82. CrossRef PubMed Google Scholar
-
103.Shi Z-Z, Miao F-P, Fang S-T, Yin X-L, Ji N-Y. Trichorenins A–C, algicidal tetracyclic metabolites from the marine-alga-epiphytic fungus Trichoderma virens Y13–3. J Nat Prod. 2018;81: 1121-4. CrossRef PubMed Google Scholar
-
104.Mereyala HB, Gadikota RR. A concise synthesis of harzialactone A from d-glucose and revision of absolute stereochemistry. Tetrahedron Asym. 1999;10: 2305-6. CrossRef PubMed Google Scholar
-
105.Sun S, Tian L, Wu Z-H, Chen G, Wu H-H, Wang Y-N, Pei Y-H. Two new compounds from fermentation liquid of the marine fungus Trichoderma atroviride G20–12. J Asian Nat Prod Res. 2009;11: 898-903. CrossRef PubMed Google Scholar
-
106.Abdel-Lateff A, Fisch K, Wright AD. Trichopyrone and other constituents from the marine sponge-derived fungus Trichoderma sp. Z Naturforsch. 2009;64c: 186-92. PubMed Google Scholar
-
107.Yamazaki H, Rotinsulu H, Narita R, Takahashi R, Namikoshi M. Induced production of halogenated epidithiodiketopiperazines by a marine-derived Trichoderma cf. brevicompactum with sodium halides. J Nat Prod. 2015;78: 2319-21. CrossRef PubMed Google Scholar
-
108.Yamazaki H, Takahashi O, Murakami K, Namikoshi M. Induced production of a new unprecedented epitrithiodiketopiperazine, chlorotrithiobrevamide, by a culture of the marine-derived Trichoderma cf. brevicompactum with dimethyl sulfoxide. Tetrahedron Lett. 2015;56: 6262-5. CrossRef PubMed Google Scholar
-
109.Shi Z-Z, Miao F-P, Fang S-T, Yin X-L, Ji N-Y. Sulfurated diketopiperazines from an algicolous isolate of Trichoderma virens. Phytochem Lett. 2018;27: 101-4. CrossRef PubMed Google Scholar
-
110.Lu X, Tian L, Chen G, Xu Y, Wang H-F, Li Z-Q, Pei Y-H. Three new compounds from the marine-derived fungus Trichoderma atroviride G20–12. J Asian Nat Prod Res. 2012;14: 647-51. CrossRef PubMed Google Scholar
-
111.Sun Y, Tian L, Huang Y-F, Sha Y, Pei Y-H. A new cyclotetrapeptide from marine fungus Trichoderma reesei. Pharmazie. 2006;61: 809-10. PubMed Google Scholar
-
112.Garo E, Starks CM, Jensen PR, Fenical W, Lobkovsky E, Clardy J. Trichodermamides A and B, cytotoxic modified dipeptides from the marine-derived fungus Trichoderma virens. J Nat Prod. 2003;66: 423-6. CrossRef PubMed Google Scholar
-
113.Yamazaki H, Rotinsulu H, Takahashi O, Kirikoshi R, Namikoshi M. Induced production of a new dipeptide with a disulfide bridge by long-term fermentation of marine-derived Trichoderma cf. brevicompactum. Tetrahedron Lett. 2016;57: 5764-7. CrossRef PubMed Google Scholar
-
114.Pruksakorn P, Arai M, Kotoku N, Vilchèze C, Baughn AD, Moodley P, Jacobs WR Jr, Kobayashi M. Trichoderins, novel aminolipopeptides from a marine sponge-derived Trichoderma sp., are active against dormant mycobacteria. Bioorg Med Chem Lett. 2010;20: 3658-63. CrossRef PubMed Google Scholar
-
115.Kavianinia I, Kunalingam L, Harris PWR, Cook GM, Brimble MA. Total synthesis and stereochemical revision of the anti-tuberculosis peptaibol trichoderin A. Org Lett. 2016;18: 3878-81. CrossRef PubMed Google Scholar
-
116.Ren J, Xue C, Tian L, Xu M, Chen J, Deng Z, Proksch P, Lin W. Asperelines A–F, peptaibols from the marine-derived fungus Trichoderma asperellum. J Nat Prod. 2009;72: 1036-44. CrossRef PubMed Google Scholar
-
117.Chen L, Zhong P, Pan J-R, Zhou K-J, Huang K, Fang Z-X, Zhang Q-Q. Asperelines G and H, two new peptaibols from the marine-derived fungus Trichoderma asperellum. Heterocycles. 2013;87: 645-55. CrossRef PubMed Google Scholar
-
118.Ren J, Yang Y, Liu D, Chen W, Proksch P, Shao B, Lin W. Sequential determination of new peptaibols asperelines G-Z12 produced by marine-derived fungus Trichoderma asperellum using ultrahigh pressure liquid chromatography combined with electrospray-ionization tandem mass spectrometry. J Chromatogr A. 2013;1309: 90-5. CrossRef PubMed Google Scholar
-
119.Mohamed-Benkada M, Montagu M, Biard J-F, Mondeguer F, Verite P, Dalgalarrondo M, Bissett J, Pouchus YF. New short peptaibols from a marine Trichoderma strain. Rapid Commun Mass Spectrom. 2006;20: 1176-80. CrossRef PubMed Google Scholar
-
120.Ruiz N, Wielgosz-Collin G, Poirier L, Grovel O, Petit KE, Mohamed-Benkada M, du Pont TR, Bissett J, Vérité P, Barnathan G, Pouchus YF. New trichobrachins, 11-residue peptaibols from a marine strain of Trichoderma longibrachiatum. Peptides. 2007;28: 1351-8. CrossRef PubMed Google Scholar
-
121.Carroux A, Van Bohemen A-I, Roullier C, du Pont TR, Vansteelandt M, Bondon A, Zalouk-Vergnoux A, Pouchus YF, Ruiz N. Unprecedented 17-residue peptaibiotics produced by marine-derived Trichoderma atroviride. Chem Biodivers. 2013;10: 772-86. CrossRef PubMed Google Scholar
-
122.Panizel I, Yarden O, Ilan M, Carmeli S. Eight new peptaibols from sponge-associated Trichoderma atroviride. Mar Drugs. 2013;11: 4937-60. CrossRef PubMed Google Scholar
-
123.Mohamed-Benkada M, Pouchus YF, Vérité P, Pagniez F, Caroff N, Ruiz N. Identification and biological activities of long-chain peptaibols produced by a marine-derived strain of Trichoderma longibrachiatum. Chem Biodivers. 2016;13: 521-30. CrossRef PubMed Google Scholar
-
124.Touati I, Ruiz N, Thomas O, Druzhinina IS, Atanasova L, Tabbene O, Elkahoui S, Benzekri R, Bouslama L, Pouchus YF, Limam F. Hyporientalin A, an anti-Candida peptaibol from a marine Trichoderma orientale. World J Microbiol Biotechnol. 2018;34: 98. CrossRef PubMed Google Scholar
-
125.Hao M-J, Chen P-N, Li H-J, Wu F, Zhang G-Y, Shao Z-Z, Liu X-P, Ma W-Z, Xu J, Mahmud T, Lan W-J. β-Carboline alkaloids from the deep-sea fungus Trichoderma sp. MCCC 3A01244 as a new type of anti-pulmonary fibrosis agent that inhibits TGF-b/Smad signaling pathway. Front Microbiol. 2022;13: 947226. CrossRef PubMed Google Scholar
-
126.Wu B, Oesker V, Wiese J, Schmaljohann R, Imhoff JF. Two new antibiotic pyridones produced by a marine fungus, Trichoderma sp. strain MF106. Mar Drugs. 2014;12: 1208-19. CrossRef PubMed Google Scholar
-
127.Liu T, Li Z-L, Wang Y, Tian L, Pei Y-H, Hua H-M. A new alkaloid from the marine-derived fungus Hypocrea virens. Nat Prod Res. 2011;25: 1596-9. CrossRef PubMed Google Scholar
-
128.Ni M, Lin Y-N, Deng C-L. A novel alkaloid from the marine-derived fungus Trichoderma citrinoviride. Chin J Mar Drugs. 2020;39: 71-6. PubMed Google Scholar
-
129.Ding L-J, Yuan W, Li Y-X, Liao X-J, Sun H, Peng Q, Han B-N, Lin H-W, Li Z-Y, Yang F, Xu S-H. Hypocrol A, a new tyrosol derivative from a sponge-derived strain of the fungus Hypocrea koningii. Nat Prod Res. 2016;30: 1633-8. CrossRef PubMed Google Scholar
-
130.Rehman SU, Wu J-S, Yang L-J, Ting S, Shao C-L, Wang C-Y. One new terphenyl glycoside from a sponge-derived fungus Trichoderma reesei (HN-2016-018). Nat Prod Commun. 2020;15: 1-5. PubMed Google Scholar
-
131.Huang H-R, Xia X-K, She Z-G, Lin Y-C, Vrijmoed LLP, Jones EBG. A new chloro-monoterpene from the mangrove endophytic fungus Tryblidiopycnis sp. (4275). J Asian Nat Prod Res. 2006;8: 609-12. CrossRef PubMed Google Scholar
-
132.Ebel R. Terpenes from marine-derived fungi. Mar Drugs. 2010;8: 2340-68. CrossRef PubMed Google Scholar
-
133.Li C-S, Liu L-T, Yang L, Li J, Dong X. Chemistry and bioactivity of marine-derived bisabolane sesquiterpenoids: a review. Front Chem. 2022;10: 881767. CrossRef PubMed Google Scholar
-
134.Shu H-Z, Peng C, Bu L, Guo L, Liu F, Xiong L. Bisabolane-type sesquiterpenoids: Structural diversity and biological activity. Phytochemistry. 2021;192: 112927. CrossRef PubMed Google Scholar
-
135.Grove JF. Non-macrocyclic trichothecenes. Nat Prod Rep. 1988;5: 187-209. CrossRef PubMed Google Scholar
-
136.Degenkolb T, Dieckmann R, Nielsen KF, Gräfenhan T, Theis C, Zafari D, Chaverri P, Ismaiel A, Brückner H, von Döhren H, Thrane U, Petrini O, Samuels GJ. The Trichoderma brevicompactum clade: a separate lineage with new species, new peptaibiotics, and mycotoxins. Mycol Progress. 2008;7: 177-219. CrossRef PubMed Google Scholar
-
137.Yasumura R, Ashtekar KD, Tonouchi A, Nehira T, Borhan B, Hashimoto M. 7-β- and 10-β-Hydroxylated congeners of CAF-603; elucidation of absolute configuration of CAF-603 family, and their SAF studies in the anti-fungal activity. Tetrahedron. 2013;69: 9469-74. CrossRef PubMed Google Scholar
-
138.Liu X-H, Song Y-P, Yin X-L, Ji N-Y. Antimicrobial terpenoids and polyketides from the algicolous fungus Byssochlamys spectabilis RR-dl-2-13. J Agric Food Chem. 2022;70: 4658-66. CrossRef PubMed Google Scholar
-
139.Jiang L, Wen Y, Peng Y, Chen T, Chen J, Yang J, Gong T, Zhu P. Advances in biosynthesis of cadinane sesquiterpenes. Chin J Biotech. 2021;37: 1952-67. PubMed Google Scholar
-
140.Hönig M, Carreira EM. Total synthesis and structural revision of a harziane diterpenoid. Angew Chem Int Ed. 2020;59: 1192-6. CrossRef PubMed Google Scholar
-
141.Chantrapromma S, Jeerapong C, Phupong W, Quah CK, Fun H-K. Trichodermaerin: a diterpene lactone from Trichoderma asperellum. Acta Cryst. 2014;E70: o408-9. PubMed Google Scholar
-
142.Barra L, Dickschat JS. Harzianone biosynthesis by the biocontrol fungus Trichoderma. ChemBioChem. 2017;18: 2358-65. CrossRef PubMed Google Scholar
-
143.Yamamoto T, Izumi N, Ui H, Sueki A, Masuma R, Nonaka K, Hirose T, Sunazuka T, Nagai T, Yamada H, Ōmura S, Shiomi K. Wickerols A and B: novel anti-influenza virus diterpenes produced by Trichoderma atroviride FKI-3849. Tetrahedron. 2012;68: 9267-71. CrossRef PubMed Google Scholar
-
144.Sun P-X, Zheng C-J, Li W-C, Jin G-L, Huang F, Qin L-P. Trichodermanin A, a novel diterpenoid from endophytic fungus culture. J Nat Med. 2011;65: 381-4. CrossRef PubMed Google Scholar
-
145.Deng J, Ning Y, Tian H, Gui J. Divergent synthesis of antiviral diterpenes wickerols A and B. J Am Chem Soc. 2020;142: 4690-5. CrossRef PubMed Google Scholar
-
146.Ui H, Shiomi K, Yamaguchi Y, Masuma R, Nagamitsu T, Takano D, Sunazuka T, Namikoshi M, Ōmura S. Nafuredin, a novel inhibitor of NADH-fumarate reductase, produced by Aspergillus niger FT-0554. J Antibiot. 2001;54: 234-8. CrossRef PubMed Google Scholar
-
147.Zhang Z, He X, Zhang G, Che Q, Zhu T, Gu Q, Li D. Inducing secondary metabolite production by combined culture of Talaromyces aculeatus and Penicillium variabile. J Nat Prod. 2017;80: 3167-71. CrossRef PubMed Google Scholar
-
148.Baldwin JE, Derome AE, Field LD, Gallagher PT, Taha AA, Thaller V, Brewer D, Taylor A. Biosynthesis of a cyclopentyl dienyl isonitrile acid in cultures of the fungus Trichoderma hamatum (Bon) Bain. aggr. J Chem Soc Chem Commun. 1981;1981: 1227-9. CrossRef PubMed Google Scholar
-
149.Sugaya K, Koshino H, Hongo Y, Yasunaga K, Onose J-I, Yoshikawa K, Abe N. The biosynthesis of sorbicillinoids in Trichoderma sp. USF-2690: prospect for the existence of a common precursor to sorbicillinol and 5-epihydroxyvertinolide, a new sorbicillinoid member. Tetrahedron Lett. 2008;49: 654-7. CrossRef PubMed Google Scholar
-
150.Harned AM, Volp KA. The sorbicillinoid family of natural products: isolation, biosynthesis, and synthetic studies. Nat Prod Rep. 2011;28: 1790-810. CrossRef PubMed Google Scholar
-
151.Wood JL, Thompson BD, Yusuff N, Pflum DA, Matthäus MSP. Total synthesis of (±)-epoxysorbicillinol. J Am Chem Soc. 2001;123: 2097-8. CrossRef PubMed Google Scholar
-
152.Pettus LH, Water RW, Pettus TRR. Synthesis of (±)-epoxysorbicillinol using a novel cyclohexa-2,5-dienone with synthetic applications to other sorbicillin derivatives. Org Lett. 2001;3: 905-8. CrossRef PubMed Google Scholar
-
153.Meng J, Wang X, Xu D, Fu X, Zhang X, Lai D, Zhou L, Zhang G. Sorbicillinoids from fungi and their bioactivities. Molecules. 2016;21: 715. CrossRef PubMed Google Scholar
-
154.Samuels GJ, Ismaiel A, Mulaw TB, Szakacs G, Druzhinina IS, Kubicek CP, Jaklitsch WM. The Longibrachiatum clade of Trichoderma: a revision with new species. Fungal Divers. 2012;55: 77-108. CrossRef PubMed Google Scholar
-
155.Shigehisa H, Suwa Y, Furiya N, Nakaya Y, Fukushima M, Ichihashi Y, Hiroya K. Stereocontrolled synthesis of trichodermatide A. Angew Chem Int Ed. 2013;52: 3646-9. CrossRef PubMed Google Scholar
-
156.Liu G, Wang Z. Total synthesis of koninginin D, B and E. Synthesis. 2001;2001: 119-27. CrossRef PubMed Google Scholar
-
157.Satoh Y, Tajima K, Munekata M, Keasling JD, Lee TS. Engineering of a tyrosol-producing pathway, utilizing simple sugar and the central metabolic tyrosine, in Escherichia coli. J Agric Food Chem. 2012;60: 979-84. CrossRef PubMed Google Scholar
-
158.Fan D-M, Liu Y, Tang L. Review on pharmacological activities of natural xanthone. Nat Prod Res Dev. 2017;29: 503-10. PubMed Google Scholar
-
159.Siddamurthi S, Gutti G, Jana S, Kumar A, Singh SK. Anthraquinone: a promising scaffold for the discovery and development of therapeutic agents in cancer therapy. Future Med Chem. 2020;12: 1037-69. CrossRef PubMed Google Scholar
-
160.Xu Y, Vinas M, Alsarrag A, Su L, Pfohl K, Rohlfs M, Schäfer W, Chen W, Karlovsky P. Bis-naphthopyrone pigments protect filamentous ascomycetes from a wide range of predators. Nat Commun. 2019;10: 3579. CrossRef PubMed Google Scholar
-
161.Bian G, Han Y, Hou A, Yuan Y, Liu X, Deng Z, Liu T. Releasing the potential power of terpene synthases by a robust precursor supply platform. Metab Eng. 2017;42: 1-8. CrossRef PubMed Google Scholar
-
162.Kumar DN, Reddy CR, Das B. Stereoselective synthesis of cytotoxic marine metabolite harzialactone A by three different routes. Synthesis. 2011;2011: 3190-4. CrossRef PubMed Google Scholar
-
163.Ballaschk F, Özkaya Y, Kirsch SF. Stereocontrolled synthesis of harzialactone A and its three stereoisomers by use of standardized polyketide building blocks. Eur J Org Chem. 2020;2020: 6078-80. CrossRef PubMed Google Scholar
-
164.Chen B, Yin H-F, Wang Z-S, Xu J-H. New synthesis of harzialactone A via kinetic resolution using recombinant Fusarium proliferatum lactonase. Tetrahedron Asymm. 2010;21: 237-40. CrossRef PubMed Google Scholar
-
165.Wang Y-N, Mou Y-H, Dong Y, Wu Y, Liu B-Y, Bai J, Yan D-J, Zhang L, Feng D-Q, Pei Y-H, Hu Y-C. Diphenyl ethers from a marine-derived Aspergillus sydowii. Mar Drugs. 2018;16: 451. CrossRef PubMed Google Scholar
-
166.Zhang Y, Li X-M, Shang Z, Li C-S, Ji N-Y, Wang B-G. Meroterpenoid and diphenyl ether derivatives from Penicillium sp. MA-37, a fungus isolated from marine mangrove rhizospheric soil. J Nat Prod. 2012;75: 1888-95. CrossRef PubMed Google Scholar
-
167.Xu X, Liu L, Zhang F, Wang W, Li J, Guo L, Che Y, Liu G. Identification of the first diphenyl ether gene cluster for pestheic acid biosynthesis in plant endophyte Pestalotiopsis fici. ChemBioChem. 2014;15: 284-92. CrossRef PubMed Google Scholar
-
168.Wan X, Doridot G, Joullié MM. Progress towards the total synthesis of trichodermamides A and B: construction of the oxazine ring moiety. Org Lett. 2007;9: 977-80. CrossRef PubMed Google Scholar
-
169.Wan X, Joullié MM. Enantioselective total syntheses of trichodermamides A and B. J Am Chem Soc. 2008;130: 17236-7. CrossRef PubMed Google Scholar
-
170.Lu C-D, Zakarian A. Total synthesis of (±)-trichodermamide B and of a putative biosynthetic precursor to aspergillazine A using an oxaza-cope rearrangement. Angew Chem Int Ed. 2008;47: 6829-31. CrossRef PubMed Google Scholar
-
171.Mfuh AD, Zhang Y, Stephens DE, Vo AXT, Arman HD, Larionov OV. Concise total synthesis of trichodermamides A, B, and C enabled by an efficient construction of the 1,2-oxazadecaline core. J Am Chem Soc. 2015;137: 8050-3. CrossRef PubMed Google Scholar
-
172.Jans PE, Mfuh AM, Arman HD, Shaffer CV, Larionov OV, Mooberry SL. Cytotoxicity and mechanism of action of the marine-derived fungal metabolite trichodermamide B and synthetic analogues. J Nat Prod. 2017;80: 676-83. CrossRef PubMed Google Scholar
-
173.Capon RJ, Ratnayake R, Stewart M, Lacey E, Tennant S, Gill JH. Aspergillazines A–E: novel heterocyclic dipeptides from an Australian strain of Aspergillus unilateralis. Org Biomol Chem. 2005;3: 123-9. CrossRef PubMed Google Scholar
-
174.Lin Y, Shao Z, Jiang G, Zhou S, Cai J, Vrijmoed LLP, Jones EBG. Penicillazine, a unique quinolone derivative with 4H–5,6-dihydro-1,2-oxazine ring system from the marine fungus Penicillium sp. (strain #386) from the South China Sea. Tetrahedron. 2000;56: 9607-9. CrossRef PubMed Google Scholar
-
175.Liu R, Gu Q-Q, Zhu W-M, Cui C-B, Fan G-T. Trichodermamide A and aspergillazine A, two cytotoxic modified dipeptides from a marine-derived fungus Spicaria elegans. Arch Pharm Res. 2005;28: 1042-6. CrossRef PubMed Google Scholar
-
176.Röhrich CR, Jaklitsch WM, Voglmayr H, Iversen A, Vilcinskas A, Nielsen KF, Thrane U, von Döhren H, Brückner H, Degenkolb T. Front line defenders of the ecological niche! Screening the structural diversity of peptaibiotics from saprotrophic and fungicolous Trichoderma/Hypocrea species. Fungal Divers. 2014;69: 117-46. CrossRef PubMed Google Scholar
-
177.Bills G, Li Y, Chen L, Yue Q, Niu X-M, An Z. New insights into the echinocandins and other fungal non-ribosomal peptides and peptaibiotics. Nat Prod Rep. 2014;31: 1348-75. CrossRef PubMed Google Scholar
-
178.Degenkolb T, Brückner H. Peptaibiomics: towards a myriad of bioactive peptides containing Cα-dialkylamino acids. Chem Biodivers. 2008;5: 1817-43. CrossRef PubMed Google Scholar
-
179.de Silva ED, Geiermann AS, Mitova MI, Kuegler P, Blunt JW, Cole ALJ, Munro MHG. Isolation of 2-pyridone alkaloids from a New Zealand marine-derived Penicillium species. J Nat Prod. 2009;72: 477-9. CrossRef PubMed Google Scholar
-
180.Ghisalberti EL. Bioactive tetramic acid metabolites. Stud Nat Prod Chem. 2003;28: 109-63. CrossRef PubMed Google Scholar
-
181.Pedersen AT, Ringborg R, Grotkjær T, Pedersen S, Woodley JM. Synthesis of 5-hydroxymethylfurfural (HMF) by acid catalyzed dehydration of glucose-fructose mixtures. Chem Eng J. 2015;273: 455-64. CrossRef PubMed Google Scholar
-
182.Clinger JA, Zhang Y, Liu Y, Miller MD, Hall RE, Van Lanen SG, Phillips GN Jr, Thorson JS, Elshahawi SI. Structure and function of a dual reductase-dehydratase enzyme system involved in p-terphenyl biosynthesis. ACS Chem Biol. 2021;16: 2816-24. CrossRef PubMed Google Scholar
-
183.Liu J-K. Natural terphenyls: developments since 1877. Chem Rev. 2006;106: 2209-23. CrossRef PubMed Google Scholar
-
184.Ji N-Y, Wang B-G. Mycochemistry of marine algicolous fungi. Fungal Divers. 2016;80: 301-42. CrossRef PubMed Google Scholar
-
185.Song Y, Miao F, Liu X, Ji N. Responses of marine-derived Trichoderma fungi to seawater and their potential antagonistic behavior. J Oceanol Limnol. 2019;37: 525-34. CrossRef PubMed Google Scholar
-
186.Pinedo-Rivilla C, Aleu J, Durán-Patrón R. Cryptic metabolites from marine-derived microorganisms using OSMAC and epigenetic approaches. Mar Drugs. 2022;20: 84. CrossRef PubMed Google Scholar
-
187.Shang Z, Salim AA, Capon RJ. Chaunopyran A: co-cultivation of marine mollusk-derived fungi activates a rare class of 2-alkenyl-tetrahydropyran. J Nat Prod. 2017;80: 1167-72. CrossRef PubMed Google Scholar
-
188.Kossuga MH, Ferreira AG, Sette LD, Berlinck RGS. Two polyketides from a co-culture of two marine-derived fungal strains. Nat Prod Commun. 2013;8: 721-4. PubMed Google Scholar
-
189.Zhang L, Niaz SI, Khan D, Wang Z, Zhu Y, Zhou H, Lin Y, Li J, Liu L. Induction of diverse bioactive secondary metabolites from the mangrove endophytic fungus Trichoderma sp. (strain 307) by co-cultivation with Acinetobacter johnsonii (strain B2). Mar Drugs. 2017;15: 35. CrossRef PubMed Google Scholar
-
190.Zhang L, Niaz SI, Wang Z, Zhu Y, Lin Y, Li J, Liu L. α-Glucosidase inhibitory and cytotoxic botryorhodines from mangrove endophytic fungus Trichoderma sp. 307. Nat Prod Res. 2018;32: 2887-92. CrossRef PubMed Google Scholar
-
191.Marmann A, Aly AH, Lin W, Wang B, Proksch P. Co-cultivation-a powerful emerging tool for enhancing the chemical diversity of microorganisms. Mar Drugs. 2014;12: 1043-65. CrossRef PubMed Google Scholar
-
192.Noguchi A, Inohara-Ochiai M, Ishibashi N, Fukami H, Nakayama T, Nakao M. A novel glucosylation enzyme: molecular cloning, expression, and characterization of Trichoderma viride JCM22452 α-amylase and enzymatic synthesis of some flavonoid monoglucosides and oligoglucosides. J Agric Food Chem. 2008;56: 12016-24. CrossRef PubMed Google Scholar
-
193.Yang Y, Ding Y, Zhang T. Biotransformation of geniposide into genipin by immobilized Trichoderma reesei and conformational study of genipin. Biomed Res Int. 2018;2018: 2079195. PubMed Google Scholar
-
194.Leutou AS, Yang G, Nenkep VN, Siwe XN, Feng Z, Khong TT, Choi HD, Kang JS, Son BW. Microbial transfromation of a monoterpene, geraniol, by the marine-derived fungus Hypocrea sp. J Microbiol Biotechnol. 2009;19: 1150-2. PubMed Google Scholar
-
195.Saravanakumar K, Kathiresan K. Bioremoval of the synthetic dye malachite green by marine Trichoderma sp. Springerplus. 2014;3: 631. CrossRef PubMed Google Scholar
-
196.Vacondio B, Birolli WG, Ferreira IM, Seleghim MHR, Gonçalves S, Vasconcellos SP, Porto ALM. Biodegradation of pentachlorophenol by marine-derived fungus Trichoderma harzianum CBMAI 1677 isolated from ascidian Didemnun ligulum. Biocatal Agric Biotechnol. 2015;4: 266-75. CrossRef PubMed Google Scholar
-
197.Alvarenga N, Birolli WG, Nitschke M, de de O Rezende MO, Seleghim MHR, Porto ALM. Biodegradation of chlorpyrifos by whole cells of marine-derived fungi Aspergillus sydowii and Trichoderma sp. J Microb Biochem Technol. 2015;7: 133-9. PubMed Google Scholar
-
198.Dini I, Alborino V, Lanzuise S, Lombardi N, Marra R, Balestrieri A, Ritieni A, Woo SL, Vinale F. Trichoderma enzymes for degradation of aflatoxin B1 and ochratoxin A. Molecules. 2022;27: 3959. CrossRef PubMed Google Scholar
Copyright information
© The Author(s) 2024
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
