 2017, Vol. 47
2017, Vol. 47
锥叶蕨属(Coniopteris)是最古老的蚌壳蕨科(Dicksoniaceae)植物, 起源于早侏罗世或更早, 中侏罗世达到了繁盛, 至早白垩世晚期逐渐衰退。其化石记录较多, 是世界性分布种, 主要分布于北半球。全球已描述的锥叶蕨属植物有60余种(马清温, 1996)[1], 同时仍有新种不断被建立(Cantrill, 2005;Kostina, 2013; Scanu, 2015)[2-4]。锥叶蕨属植物常在温带、亚热带、热带区域大量出现, 因而有很多学者(Vakhrameev, 1991;王永栋, 2002)[5-6]认为该属是温暖、湿润环境的指示分子。邓胜徽等(2002)认为,锥叶蕨属植物曾是一种具有匍匐根状茎的蕨类植物[7]。Bystritskaya(2013)对西西伯利亚锥叶蕨进行了再研究, 分析描述了6种锥叶蕨植物化石[8]。Balame(1995)对锥叶蕨属的原位孢子进行了研究, 系统总结了十多种锥叶蕨属植物化石原位孢子的形态特征[9]。陈芬等(2001)对中国东北地区早白垩世真蕨类植物化石繁殖器官微细构造进行了细致的研究[10]。
Coniopteris spectabilis是中侏罗世的代表分子, 同时在早侏罗世晚期也有分布, 最先发现于费尔干纳东部早侏罗世晚期的查阿尔塔什组至中侏罗世早期的津丹含煤组。中国已经发现的锥叶蕨属植物化石达40种,但目前对锥叶蕨属的研究, 尤其是C.spectabilis的微细构造研究相对较少, 仅李佩娟(1988)对其大化石进行了较细致描述[11]。同时,C. spectabilis大化石发现相对较少, 仅在大兴安岭南部(杨学林, 1985)、青海柴达木盆地(邓文诗, 2009)、新疆吐哈盆地(商平, 1999)、辽西义县(丁秋红, 2003)、甘肃靖远王家山盆地(厉宝贤, 1982)、宝积山盆地(陈应权, 2017)和窑街盆地(孙柏年, 1986)有发现[12-18]。本研究区窑街盆地不仅含有丰富的植物大化石,而且含有大量孢粉化石, 因而是研究侏罗纪植物的理想地区之一。孙柏年等(1986)对甘肃窑街的植物大化石进行了研究[18];闫德飞(2004)在甘肃窑街中侏罗统窑街组发现了穆雷似管状叶Solenites murrayana L.et H., 并对其表皮构造进行了研究[19];张成君等(2004)分析研究了中侏罗统窑街组化石银杏以及现代银杏角质层的有机质组分, 从而进行了古环境重建[20];辛存林等(2006)分析了窑街盆地中侏罗世孢粉化石,建立了窑街盆地中侏罗世的孢粉组合[21];辛存林等(2010)研究了甘肃窑街中侏罗世5种蕨类植物的原位孢子与分散孢子的关系, 并于2011,2012年分别对C.margaretae和C.simplex进行了细致研究[22-24]。而本文对发现于兰州窑街盆地、保存较好的C. spectabilis生殖叶和营养叶化石进行了详细研究, 补充了其末次羽片、孢子囊及原位孢子方面的数据。同时,C. spectabilis大化石在甘肃窑街的发现不仅对研究锥叶蕨属系统演化提供了材料, 而且为准确分析该属的自然地理分布提供了有力的证据。
1 区域概况窑街盆地位于兰州市西北部(102°50′E, 36°25′N), 海拔约1 520 m, 位于甘肃、青海两省交界处(见图 1);其东西两侧分别为元古界和中、新生界构成的山地,大通河流经其间,为一典型的现代山间河谷盆地。现代地貌形态表现为南北向展布的狭长弧形盆地, 大地构造单元上属中祁连隆起带, 处于祁连褶皱山系与兰州古地块的交接部位, 是一个中、新生代形成和发展起来的内陆盆地。

|
图 1 化石采样点位置图 Fig. 1 Fossil plant locality in Yaojie Basin |
窑街组为一套中侏罗统含煤地层, 主要为河流—湖沼相岩石组合, 其间夹有含铁锰结核和条带的褐红色砂岩和黏土岩组成的风化壳, 总厚度约235 m。窑街组自下而上可分为5个岩性段(见图 2):砂砾岩段(J2 y1)、含煤岩段(J2y2)、铝质泥岩段(J2y3)、油页岩段(J2y4)、砂泥岩段(J2y5)。本文化石采集于砂泥岩段(J2y5), 厚70 m, 富含铁质结核, 与上覆晚侏罗世享堂组呈假整合接触, 含有丰富的植物化石。

|
图 2 甘肃窑街中侏罗统地层剖面柱状图 Fig. 2 Thestratigraphic section through the Yaojie Formation, Gansu Province |
本研究所用的化石标本, 其营养叶和生殖叶标本采于甘肃窑街盆地, 地层层位为中侏罗统窑街组砂泥岩段(J2 y5)。
化石原位孢子的处理方法:将生殖羽片上的孢子囊群用揭片法或用小刀剥离后, 先用盐酸除掉样品中的钙质; 然后将样品装入10~20 mL的塑料离心试管中, 加39%的氢氟酸处理, 以便除去样品中的硅质和黏土物质, 用氢氟酸冷处理24小时, 或用热水浴煮15~20分钟; 再用5%的稀硝酸处理; 将5%~10%的氢氧化钾加入到水洗至中性的样品, 若发现孢子粒纹清晰, 碳质碎片变黄, 则可停止处理。部分孢子样品用甘油胶制片, 在光学显微镜下观察、照相,并统计孢子产量, 另一部分孢子样品备用(邓胜徽等, 1995)[25]。
3 化石系统描述纲:真蕨纲Filicopsida
目:真蕨目Filicales
科:蚌壳蕨科Dicksoniaceae
属:锥叶蕨属Coniopteris Brongnart, 1948
种:奇丽锥叶蕨Coniopteris spectabilis Brick 1953(图 3, 图 4, 图 5)

|
A裸羽片, 示末次羽片及小羽片形态, 箭头所示为变态小羽片, 比例尺=1 cm; B裸羽片局部放大, 示变态小羽片形态, 比例尺=2 cm; C实羽片, 实羽片上部着生部分裸小羽片, 比例尺=1 cm; D实小羽片, 箭头所示为孢子囊群, 比例尺=0.5 cm 图 3 兰州窑街组Coniopteris spectabilis Harris蕨叶形态及生殖器官 Fig. 3 Leaves, sporangia and in situ spores of Coniopteris spectabilis Harris |

|
图 4 Coniopteris spectabilis末次羽片素描图 Fig. 4 Line drawing of pinna of Coniopteris spectabilis |

|
图 5 兰州窑街组Coniopteris spectabilis Harris原位孢子 Fig. 5 In situ spores of Coniopteris spectabilis Harris from Yaojie Formation |
产地及层位 兰州窑街, 中侏罗统窑街组砂泥岩段(J2 y5)。
化石描述 当前化石为二次羽状复叶, 羽轴宽1 mm, 具纵纹。末次羽片披针形或线形, 向顶端渐狭, 长约4.5 cm, 宽约1 cm, 最宽处在基部可达1.5 cm, 与羽轴成45°~65°角(见图 3A, 图 4)。小羽片长菱形, 长4~8 mm, 宽2~4 mm。正常小羽片深裂为三对裂片, 再加顶端一单个裂片, 共7枚裂片。裂片顶端钝尖, 呈锯齿或分叉, 或分裂成二叉或三齿状, 顶端裂片的三齿较为明显。基部上行第一枚小羽片明显变大, 其长度为相邻正常小羽片的1.5~2倍, 裂片数也增为9~11;下行第一枚小羽片变态, 深裂为线形裂片或呈鸟足状(见图 3B)。叶脉楔羊齿型, 中脉不明显, 侧脉分叉2~4次。
生殖羽片及孢子囊群 末次实羽片披针形或线形, 长约6 mm, 宽约1 mm, 混生型, 实羽片上部着生部分未发育完全的裸小羽片。实小羽片强烈退缩, 卵圆形, 具短柄, 其顶端着生圆形或卵圆形孢子囊群, 长径约0.8 mm。(见图 4C,D)。
原位孢子 孢子为四面体形, 极面观为三角形、圆三角形, 黄褐色, 顶角钝圆, 三边线平直或微凸(见图 5B,C,D)。赤道面观为半圆形、椭圆形, 近极微突出(见图 5E,F,G)。极面观直径38~54 μm, 平均46 μm。三射线长度为半径的4/5, 可达赤道边缘, 微弯, 部分隆起为脊线状, 通常为幼体孢子, 体积相对较小(见图 5B)。外壁近于光滑, 相对较厚。
4 比较与讨论真蕨类植物中, 囊群着生于叶的边缘, 仅见于蚌壳蕨科(Dicksoniaceae), 据此特征,毫无疑问可将当前化石归于蚌壳蕨科。当前化石与锥叶蕨属(Coniopteris)最为相似,且依据末次羽片披针形或线形, 向顶端渐狭, 与羽轴成45°~65°角, 小羽片长菱形, 正常小羽片深裂为三对裂片, 顶端着生圆形或卵圆形孢子囊群,可将其与其他属相区别(见表 1)。蚌壳蕨属(Dicksonia)与当前化石也较为相似, 叶脉都为楔羊齿型, 裸小羽片披针形, 囊群顶生, 但是蚌壳蕨属中轴较宽, 中下部宽2~3 mm, 叶脉较为稀疏, 且生殖羽片独立, 而当前化石中轴宽1 mm左右, 侧脉分叉2~4次。爱博拉蕨属(Eboracia)和屈囊蕨属(Gonatosorus)与当前化石都具变态小羽片, 但爱博拉蕨属的叶脉和裸小羽片都为栉羊齿型或枝脉蕨型, 而当前化石为楔羊齿型;屈囊蕨属(Gonatosorus)基部下排小羽片变态呈两瓣状, 且叶脉和小羽片为枝脉蕨型, 而当前化石羽片下侧基部下行第一枚小裂片呈三叉状的指状细裂片。刺蕨属(Acanthopteris)中轴粗, 具中等宽度的中肋, 侧脉稀少, 多数不分叉, 且裸小羽片栉羊齿型。因此, 依据上述蚌壳蕨科相关属的特征对比分析, 从形态学特征可鉴定当前化石标本应归为锥叶蕨属(Coniopteris)。
|
|
表 1 蚌壳蕨科相关属枝叶形态特征对比 Tab. 1 The comparison of frond morphology between the genera of Dicksoniaceae |
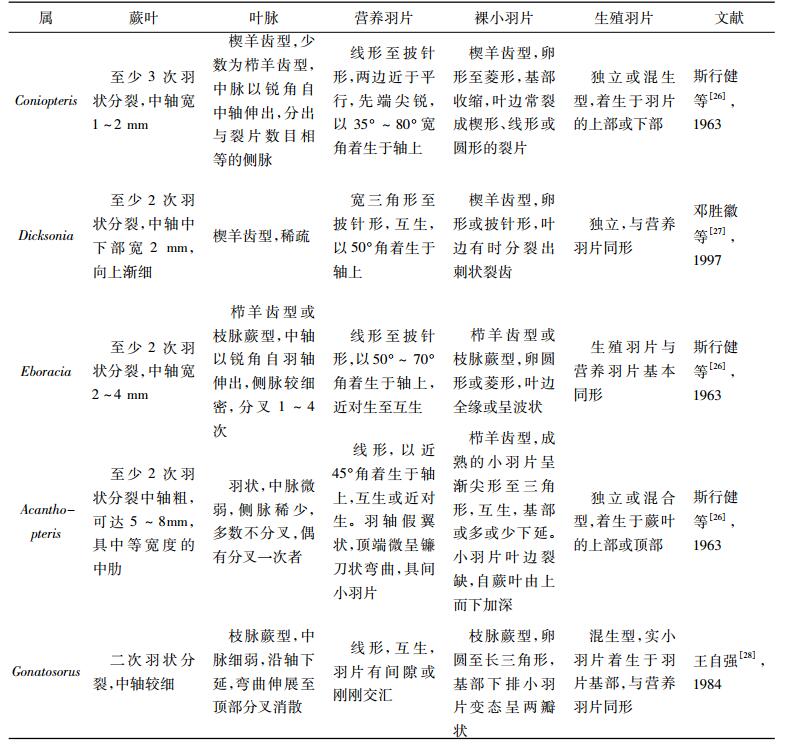
当前植物化石有两处明显的鉴定特征, 一为小羽片分裂程度高(正常小羽片分叉2~3次), 而小羽片的分裂程度也是锥叶蕨属植物化石鉴定的一个重要依据。根据己捜集文献中的文字描述和图版对照, 发现共有15种锥叶蕨属植物化石的小羽片分裂程度相对较高;对其蕨叶、叶脉、营养羽片和生殖羽片等特征进行了详细对比(见表 2)后发现, 其中Coniopteris concinna, C. zindanensis, C. huolinheensis, C. setacea, C.simplex, C. tatungensis这6种虽然分裂程度较高, 但近乎全裂, 且小裂片为刺状或细线状, 而当前化石深裂为顶端钝尖的卵形裂片, 可明显区分。与当前化石最为相似的是C.spectabilis, 小羽片都为菱形, 基部微微下延, 长约4 mm, 宽约2 mm, 多深裂为3对小裂片, 较大的小羽片裂片数可达11片, 且小裂片顶端钝尖, 这些特征都与当前化石高度一致。C.gansunsis小羽片分裂程度与当前化石较为相似, 但C.gansunsis小羽片呈卵形, 基部收缩, 长约7 mm, 宽约4 mm, 且末次羽片宽约1.2 cm, 长7 cm以上, 而当前化石末次羽片宽1 cm, 长仅4.5 cm; C. hymenophylloides小羽片卵形, 三角形或倒楔形, 边缘浅裂为5~6枚顶端钝圆或钝尖的小裂片, 但小羽片的大小、形态及边缘缺裂的大小、裂片数目均因所在位置不同而变化较大, 而当前化石小羽片大小形态较为一致; C. murrayana小羽片分裂程度也较高, 分裂呈2~3对卵形裂瓣, 裂瓣全缘或顶部圆齿状, 但较当前化石分裂得深, 近全裂; C.bella小羽片深裂或浅裂, 分裂为3~7个卵形小裂片, 但裂片顶端钝圆, 且小羽片长8 mm, 宽5 mm, 末次羽片长仅2 cm; C.tiehshanensis, C.vachrameevii, C.beijingensis, C.quinqueloba裂片数较少, 仅5片, 而当前化石常为7片, 最多可达11片, 可明显与当前化石区分。
|
|
表 2 Coniopteris spectabilis与其相似种类形态特征的比较 Tab. 2 The comparison between megafossils of Coniopteris spectabilis and other similar species |

另一鉴定特征为具变态小羽片。当前化石末次羽片基部上行第一枚小羽片明显扩大, 而下行第一枚小羽片深裂为线形裂片, 似鸟足状。侏罗纪有不少锥叶蕨具有变态的小羽片, 而羽片基部发育有线状变态小羽片的仅见于C.hymenophylloides, C.murrayana, C. bella, 结合小羽片的分裂特征及其蕨叶、叶脉、营养羽片和生殖羽片等特征, 这3种植物可与当前化石明显区分(见上述分析)。而与当前化石最为相似的C. spectabilis,目前已有的报道中并未发现有变态小羽片, 可能是由于:①目前C. spectabilis大化石发现较少, 较详细的研究仅基于几段羽片进行, 保存并不完整, 小羽片形态虽较为清晰, 但末次羽片形态模糊, 不排除变态小羽片破碎的可能; ②由于对中侏罗世锥叶蕨属变态小羽片的生态学意义还不甚清楚, 无法解释其出现的原因及机理, 当前也无法确定C.spectabilis基部变态裂片是否稳定存在的。基于上述分析, 将当前化石定为C. spectabilis是适宜的。
Coniopteris spectabilis此种的中文学名并不统一, 存在同物异名的情况。“spectabilis”翻译为“悦目”, 文献资料中主要存在两种中文用法, 奇丽锥叶蕨(辛存林等, 2010;陈应权等, 2017)[17, 22]与美丽锥叶蕨(李佩娟等, 1988;丁秋红等, 2003;孙克勤等, 2010)[11, 15, 29], 二者都能体现其好看的原意。而美丽锥叶蕨有学者也用来指C. bella, 二者的中文学名较为混乱。因此,对C. spectabilis和C. bella中文学名的统一是有必要的。“bella”翻译为中文即为美丽, 文献资料中主要存在4种中文名称:美丽锥叶蕨, 漂亮锥叶蕨, 优美锥叶蕨和贝尔锥叶蕨。沈阳地质研究所(1980)、陈芬等(1986)、米家榕等(1996)、郑少林等(2002)多数学者都主张用美丽锥叶蕨[30-33];邓胜徽等(2015)、赵怡等(2015)部分学者用漂亮锥叶蕨[34-35];少部分学者如叶美娜等(1986)主张用音译贝尔锥叶蕨[36], 孙全(2009)用优美锥叶蕨[37]。鉴于bella中文翻译即为美丽, 且美丽锥叶蕨的用法也被大多数学者认可, 本文认为C.bella中文学名用美丽锥叶蕨更适宜, 而C.spectabilis中文名称则采用奇丽锥叶蕨, 以便区分使用。
5 原位孢子与分散孢子的关系侏罗纪蚌壳蕨科的分散孢子可归入桫椤属(Cyathidites)或三角孢属(Deltoidospora), 这两属的主要区别在于后者三边不总是凹入, 三射线较短(Johanna et. al, 1989)[40]。除此之外,与蚌壳蕨科和桫椤科有亲缘关系的孢子还有光面三缝孢属(Leiotriletes), 金毛狗孢属(Cibotiumspora), 波缝孢属(Undulatisporites)等属(江德昕等, 2009)[41]。当前化石原位孢子与三角孢属(Deltoidospora)最为相似, 极面观三角形到圆三角形, 角部多数呈圆形; 三边直或微凸, 三射线简单, 其长度为孢子半径的2/3或更长, 外壁光滑, 具细褶皱。而桫椤属(Cyathidites)孢子三边明显凹入; 光面三缝孢属(Leiotriletes)孢子三边明显凹入或略微, 甚至强烈突出; 金毛狗孢属(Cibotiumspora)孢子三射线末端各有一与之正交或斜交的褶皱; 波缝孢属(Undulatisporites)孢子极面观亚三角形, 三射线为波状。这些特征与当前原位孢子特征不符,因此从形态学特征鉴定,当前化石标本应归为三角孢属(Deltoidospora)。窑街组地层中三角孢属(Deltoidospora)主要有Deltoidospora balowensis, D. irregularis(张望平等, 1985;马玉贞等, 1996;辛存林等, 2006)[21, 42-43]。Deltoidospora balowensis极面观为三角形, 三边线微凸或微凹, 极面观直径40~47 μm, 平均43.5 μm, 三射线长为孢子半径的1/2~2/3, 常微开裂。而当前化石原位孢子三射线窄而细长, 可达赤道, 且三边较为平直, 可与D. balowensis明显区分; D. irregularis极面观为三角形、圆三角形, 顶角宽圆, 三边线微凸或平直, 极面观直径36~55 μm, 平均45.5 μm, 三射线简单, 射线长为孢子半径的2/3或可达赤道, 表面光滑, 具细褶皱, 与当前化石原位孢子最为相似。因此,基于以上属种的比较,D. irregularis是Coniopteris spectabilis孢子, 后者为前者的母体植物。
6 锥叶蕨属的时空分布根据目前已搜集到的材料,中国已报道的锥叶蕨属有44种(叶美娜, 1986;陈芬, 2001;孙克勤, 2010)[10, 29, 36], 从已知的各个种的40多个化石产地来看,显示出中国锥叶蕨属的时空分布具有明显的规律性(见图 6)。气候是影响植物地理分布模式的重要因素, 因而研究锥叶蕨属时空分布模式对气候变化也有一定的指示作用。

|
图 6 中国中生代锥叶蕨属植物地理分布图 Fig. 6 The geographical distribution of the genus Coniopteris from Mesozoic in China |
中国锥叶蕨属植物最早发现于湖南观音滩组,所得化石的时代可追溯到早侏罗世早期。早侏罗世的锥叶蕨植物体较小, 但已有相当的分异, 共有15个种, 主要分布于昆仑山—秦岭—大别山一线以北, 东至郯庐断裂, 包括东北、华北及西北地区, 而这些地区基本都属于内陆地区, 煤层、煤线、菱铁矿层和暗色泥岩等指示潮湿气候的沉积普遍发育, 适宜锥叶蕨属植物生长。除此之外,其在湖南、湖北、重庆、四川也有分布, 表明在早侏罗世,这些地区也较为温暖湿润。但是,早侏罗世锥叶蕨属多样性不及中侏罗世丰富, 这可能与早侏罗世陆地生态系统发生全球性升温的事件有关, 导致锥叶蕨属发展受到了一定的限制。中侏罗世锥叶蕨属进入了繁盛阶段, 共有19个种, 蕨叶也逐渐变得粗壮起来, 这也是升温事件结束, 中侏罗世气温回落, 湿度增大的明显体现。此时锥叶蕨属植物仍主要分布于中国昆仑山—秦岭—淮河一线以北的地区, 但较之早侏罗世分布范围缩小, 且略向北移。在该线以南, 锥叶蕨属植物仅在湖北秭归和广西扶绥有分布, 体现出局部地区仍较为温暖湿润。晚侏罗世,锥叶蕨开始衰退, 种数急剧减少, 只有少数种存活下来, 代表分子如C.burejensis, 也反映出这一阶段中国气候发生了较大幅度的变化, 且变化方向并不利于锥叶蕨生长, 向干旱炎热型转变。此时锥叶蕨属在山东、北京、内蒙古、甘肃、青海有零星分布, 可以推测这些地区气候变化不如其他地区明显, 局部仍较为温暖湿润, 能够维持锥叶蕨植物的生长。早白垩世,锥叶蕨逐渐复苏, 并达到了第二次繁盛, 共有17个种, 分异度次高, 但仅在东北地区有分布。这可体现出中国内陆地区仍保持干热的状态, 而东北地区由于受到海洋气候的影响, 仍较为湿润。早白垩世以后,锥叶蕨化石的发现未见报道, 反映出早白垩世后期,中国气候有大幅度的变化, 而这种变化显然对喜湿润的锥叶蕨极为不利, 更是导致了锥叶蕨属的绝迹。基于上述分析可知,从早侏罗世到早白垩世,中国锥叶蕨属分布总体呈向北推移的趋势, 分布范围逐渐缩小, 到早白垩世仅在东北地区有分布; 且南北地区分异明显, 北方地区锥叶蕨属多样性较南方地区丰富。由此也可看出,中国这一时期暖温带气候区总体上有向北推移的趋势, 到晚侏罗—早白垩世, 中国境内仅东北地区较为温暖湿润。
7 结论1) 保存良好的营养叶与生殖叶化石在甘肃窑街的发现, 完善了Coniopteris spectabilis蕨叶形态资料, 补充了大化石时空分布的资料。同时,对Coniopteris spectabilis中文学名进行了厘定, 认为用“奇丽锥叶蕨”更适宜。
2) 对Coniopteris spectabilis原位孢子进行研究, 并与地层中的分散孢子建立联系, 确认Deltoidospora irregularis是Coniopteris spectabilis的孢子, 后者为前者的母体植物。
3) 早侏罗世至早白垩世,中国锥叶蕨属(Coniopteris)分布总体呈向高纬度推移的趋势, 分布范围逐渐缩小, 到早白垩世仅在东北地区分布;南北地区分异明显, 北方地区锥叶蕨属多样性较南方地区丰富。结合锥叶蕨属(Coniopteris)喜潮湿温暖这一特性可知,中国这一时期暖温带气候区总体上有向高纬度推移的趋势, 且北方主要为潮湿的暖温带气候, 南方则反映热带和亚热带干旱气候。
| [1] |
马清温, 王宇飞. 锥叶蕨属研究进展[J]. 植物学通报, 1996, 13(增刊): 28-34. |
| [2] |
CANTRILL D J, NAGALINGUM N S. Ferns from the Cretaceous of Alexander Island, Antarctica:Implications for Cretaceous phytogeography of the southern hemisphere[J]. Review of Palaeobotany and Palynology, 2005, 137: 83-103. DOI:10.1016/j.revpalbo.2005.08.004 |
| [3] |
KOSTINA E I, HERMAN A B. The Middle Jurassic flora of South Mongolia:Composition, age andphytogeographic position[J]. Review of Palaeobotany and Palynology, 2013, 193: 82-98. DOI:10.1016/j.revpalbo.2013.01.009 |
| [4] |
SCANU G G, KUSTATSCHER E, PITTAU P. The Jurassic flora of SardiniaA new piece in the palaeobiogeographic puzzle of the Middle Jurassic[J]. Review of Palaeobotany and Palynology, 2015, 218: 80-105. DOI:10.1016/j.revpalbo.2014.08.008 |
| [5] |
VAKHRAMEE V A. Jurassic and Cretaceous floras and climates of the Earth[M]. Cambridge: Cambridge University Press, 1991.
|
| [6] |
WANG Y D. Fern ecological implications from the Lower Jurassic in Western Hubei, China[J]. Review of Palaeobotany and Palynology, 2002, 119: 125-141. DOI:10.1016/S0034-6667(01)00133-6 |
| [7] |
DENG S H. Ecology of the Early Cretaceous ferns of Northeast China[J]. Review of Palaeobotany and Palynology, 2002, 119: 93-112. DOI:10.1016/S0034-6667(01)00131-2 |
| [8] |
BYSTRITSKAYAL. New finds of ferns from Middle Jurassic deposits of Ob Tazovsky facies area of Western Siberia[J]. Journal of Tomsk State University, 2013, 367: 170-176. |
| [9] |
BALAME B E. Fossil in situspores and pollen Grains: An annotated catalogue[J]. Review of Palaeobotany and Palynology, 1995, 87: 81-323. DOI:10.1016/0034-6667(95)93235-X |
| [10] |
陈芬, 邓胜徽. 中国东北地区早白垩世真蕨类植物[M]. 北京: 地质出版社, 2001.
|
| [11] |
李佩娟, 何元亮, 吴向武, 等. 青海柴达木盆地东北缘早、中侏罗世地层及植物群[M]. 南京: 南京大学出版社, 1988.
|
| [12] |
杨学林, 孙礼文. 大兴安岭南部早、中侏罗世地层[J]. 地层学杂志, 1985, 9(1): 10-20. |
| [13] |
邓文诗, 张丽霞, 成永盛. 青海省下中侏罗统含煤岩系沉积特征及聚煤作用[J]. 中国煤炭地质, 2009, 21(增刊2): 14-18. |
| [14] |
商平, 付国斌, 侯全政, 等. 新疆吐哈盆地中侏罗世植物化石[J]. 现代地质, 1999, 13(4): 403-407. |
| [15] |
丁秋红, 张立东, 郭胜哲, 等. 辽西地区义县组植物群古生态研究[J]. 地质与资源, 2003, 12(1): 9-18. DOI:10.3969/j.issn.1671-1947.2003.01.002 |
| [16] |
厉宝贤, 徐福祥, 马其鸿, 等. 甘肃靖远王家山盆地中侏罗世地层[J]. 地层学杂志, 1982, 6(1): 33-40. |
| [17] |
陈应权, 马利涛, 彭琳, 等. 甘肃宝积山盆地中侏罗世阿干镇似银杏(Ginkgoites aganzhenensis Yang)的古环境意义[J]. 沉积学报, 2017, 35(1): 57-66. |
| [18] |
孙柏年. 兰州窑街煤田中侏罗世植物化石的初步研究[J]. 兰州大学学报(自然科学版), 1986, 22(1): 113-118. |
| [19] |
阎德飞, 孙柏年. Solenites murrayana L.et H.在甘肃窑街煤田的发现及其地质意义[J]. 兰州大学学报(自然科学版), 2004, 40(3): 84-88. DOI:10.3321/j.issn:0455-2059.2004.03.019 |
| [20] |
张成君, 孙柏年, 阎德飞, 等. 甘肃窑街中侏罗世化石角质层有机质及其碳同位素组成与古环境的关系[J]. 西北地质, 2004, 148(37): 42-47. |
| [21] |
辛存林, 孙柏年, 韦利杰, 等. 甘肃窑街中侏罗世孢粉组合及其古环境意义[J]. 兰州大学学报(自然科学版), 2006, 42(3): 8-13. DOI:10.3321/j.issn:0455-2059.2006.03.002 |
| [22] |
辛存林, 孟健, 魏明, 等. 侏罗纪五种锥叶蕨植物原位孢子及其与分散孢子的关系[J]. 兰州大学学报(自然科学版), 2010, 46(4): 7-12. |
| [23] |
辛存林, 陈世伟, 都卫东, 等. 甘肃窑街中侏罗世Coniopteris margaretae的原位孢子与分散孢子的关系[J]. 地质科技情报, 2011, 30(5): 37-40. |
| [24] |
辛存林, 魏明, 陈世伟, 等. 甘肃窑街中侏罗世植物化石Coniopteris simplex的原位孢子特征研究[J]. 地质科学, 2012, 47(1): 211-220. |
| [25] |
邓胜徽, 姚立军. 蕨类植物化石繁殖器官微细构造研究方法及意义[J]. 现代地质, 1995, 9(1): 19-26. |
| [26] |
斯行健, 李星学, 李佩娟. 中国中生代植物(中国各门类化石, 中国植物化石)(第二册)[M]. 北京: 科学出版社, 1963.
|
| [27] |
邓胜徽, 任守勤, 陈芬. 内蒙古海拉尔地区早白垩世植物群[M]. 北京: 地质出版社, 1997.
|
| [28] |
天津地质矿产研究所. 华北地区古生物图册(中生代分册)[M]. 北京: 地质出版社, 1984.
|
| [29] |
孙克勤, 崔金钟, 王士俊. 中国化石蕨类植物, 中国化石植物志(第二卷)[M]. 北京: 高等教育出版社, 2010.
|
| [30] |
沈阳地质矿产研究所. 东北地区古生物图册(中新生代分册)[M]. 北京: 地质出版社, 1980.
|
| [31] |
陈芬, 窦亚伟, 孟祥营. 河北下花园玉带山群植物化石[J]. 中国矿业学院学报, 1986(2): 50-64. |
| [32] |
米家榕, 孙春林, 孙跃武, 等. 冀北辽西早、中侏罗世植物生态学及聚煤环境[M]. 北京: 地质出版社, 1996.
|
| [33] |
郑少林, 张武. 辽宁地质历史时期的植物界[J]. 地质与资源, 2002, 11(4): 193-215. DOI:10.3969/j.issn.1671-1947.2002.04.001 |
| [34] |
邓胜徽, 商平, 卢远征, 等. 内蒙古锡林浩特盆地早侏罗世晚期植物群:兼论中国早侏罗世晚期气候[J]. 古地理学报, 2015, 17(5): 617-634. |
| [35] |
赵怡.内蒙古锡林浩特盆地早侏罗世植物群[D].北京: 中国地质大学, 2015. http://cdmd.cnki.com.cn/Article/CDMD-11415-1015389817.htm
|
| [36] |
叶美娜, 刘兴义, 黄国清, 等. 川东北地区晚三叠世及早、中侏罗世植物[M]. 合肥: 安徽科学技术出版社, 1986.
|
| [37] |
孙全.内蒙古宁城和辽宁西部古生物地质遗迹评价及保护区划[D].北京: 中国地质大学, 2009. http://cdmd.cnki.com.cn/Article/CDMD-11415-2009075864.htm
|
| [38] |
曹正尧, 张亚玲. 甘肃侏罗系Coniopteris一新种[J]. 古生物报, 1996, 35(2): 241-247. |
| [39] |
任守勤, 陈芬. 内蒙古海拉尔五九煤盆地早白垩世大磨拐河组植物化石[J]. 古生物报, 1989, 28(5): 634-638. |
| [40] |
JOHANNA HA, CITTERT V K. Dicksoniaceous spores in situ from the Jurassic of Yorkshire, England[J]. Review of Palaeobotany and Palynology, 1989, 61: 273-301. DOI:10.1016/0034-6667(89)90035-3 |
| [41] |
江德昕, 王永栋, 魏江. 新疆乌恰中侏罗世孢粉植物群及其环境意义[J]. 古地理学报, 2009, 11(2): 205-213. |
| [42] |
张望平, 赵清顺. 甘肃窑街地区下侏罗统炭洞沟组的孢粉组合[J]. 地质论评, 1985, 31(1): 13-22. DOI:10.3321/j.issn:0371-5736.1985.01.002 |
| [43] |
马玉贞, 陶明信, 陈发源. 甘肃窑街寺湾沟—炭洞沟红层孢粉组合、地质时代与生态环境[J]. 沉积学报, 1996, 14(1): 93-102. |