 2017, Vol. 47
2017, Vol. 47
 , 李红, 牛元哲, 刘璠, 董杨坤
, 李红, 牛元哲, 刘璠, 董杨坤 已有研究表明, 青藏高原新生代鲤科鱼类化石的研究, 不仅对了解鲤科鱼类的早期演化很有意义, 而且可以为青藏高原隆升、西部干旱化过程的研究提供重要的化石证据[1-3]。柴达木盆地位于青藏高原北部, 是高原上最大的断陷盆地, 有巨厚的新生代沉积物, 是研究高原新生代生物与环境的一个理想地点。近年来,柴达木盆地鱼类化石的新发现, 如伍氏献文鱼[4]等, 以其重要的生物演化和环境变迁意义, 引人瞩目[3-5]。前人曾在柴达木盆地渐新统中发现过相当数量的鲤科鱼类化石, 不仅为了解当时当地的气候环境提供了化石依据, 而且对了解鲤科鱼类的早期演化和分布提供了线索, 对生物演化和环境变迁研究具有相当重要的意义[1, 6]。
近年来, 作者在柴达木盆地进行野外地质工作的过程中, 采集到一件保存较完整的渐新世鲃亚科鱼类化石。前人在柴达木盆地已采集的渐新世鲃亚科化石包括咽骨、咽齿、匙骨、腹鳍骨和后缘带锯齿的不分枝鳍条等, 但没有完整的鱼体化石[1]。本次发现的鲃亚科化石标本保存相对完整, 在研究区属首次发现。鉴于鱼类化石研究在恢复古环境、古气候中的重要意义, 本文对新发现的化石标本进行了研究。文中测量、描述方法及术语综合参考《中国动物志·硬骨鱼纲·鲤形目》,Cyprinid Fishes,《鱼类比较解剖学》等著作[7-10]。本次使用的板状化石CT仪器为160-Micro-CT, 检测电压105 kV, 检测电流50 μA, 分辨率11 μm, 重建解释软件为IHEP-ICLCT, 测试在中国科学院脊椎动物演化与人类起源重点实验室完成。
化石赋存于柴达木盆地西北缘茫崖地区渐新统下干柴沟组[11](见图 1), 地理坐标为38°39′N, 90°38′E,化石产地海拔3 410 m, 岩性为灰绿色—浅灰绿色钙质泥岩、灰褐色砂岩互层, 泥岩略厚, 发育水平层理, 延伸约300 m, 风化后颜色常呈土黄色。前人对研究区干柴沟组岩性组合及腹足类Hydyrobia sp.、介形类Eucypris sp.等化石、孢粉组合等的综合研究[12]认为,研究区所处地层为渐新统。此外, 张跃中等[13]对该剖面进行了高密度采样和古地磁分析, 得出该区上干柴沟组地层距今31.5~22 Ma。该化石位于下干柴沟组顶部, 地质年代约为31.5 Ma。

|
图 1 研究区位置 Fig. 1 Location of working area |
骨鳔超目 Superorder Ostariophysi Sagemehl, 1885
鲤形目 Order Cypriniformes Bleeker, 1859
鲤科 Cyprinidae Bonaparte, 1840
鲃亚科 Barbinae Berg, 1912
鲃亚科(属、种未定) Barbinae gen. et sp.indet.

|
注:af臀鳍;bfr分支鳍条;c c椎体;cf尾鳍;cs围眶骨;df背鳍;fr额骨;oc枕髁;op主鳃盖骨;pa顶骨;pf胸鳍;pha咽骨;pop前鳃盖骨;pt咽齿;sf硬棘刺;sop下鳃盖骨;vf腹鳍;vr腹肋;比例尺= 20 mm 图 2 示鲃亚科(属、种不定)化石(NWUV1487-NWUV1488) Fig. 2 Barbinae gen. et sp. indet. |

|
注:比例尺=30 mm 图 3 鲃亚科(属、种未定)头部骨骼 Fig. 3 Skull of Barbinae gen. et sp.indet. |

|
注:1.齿面后端,2后肢后端点;比例尺=3 mm 图 4 鲃亚科(属、种未定)左侧咽骨(NWUV1488) Fig. 4 Left pharyngeal bone (NWUV1488) of Barbinae gen. et sp. indet. |

|
注:比例尺=10 mm 图 5 鲃亚科(属、种未定)右侧咽骨(NWUV1488,未出露) Fig. 5 Right pharyngeal bone(NWUV1488)of Barbinae gen. et sp. indet. (not exposed) |

|
注:c椎体;cf尾鳍;e尾上骨;h尾下骨;hs脉棘;phy副尾下骨;u尾杆骨;un尾髓骨; sf硬棘刺; 比例尺= 10 mm 图 6 示鲃亚科(属、种未定)尾骨骼及硬棘刺(NWUV1487-1488) Fig. 6 Caudal skeleton and serrated unbranched fin ray of barbinae gen. et sp. indet. (NWUV1487-1488) |
|
|
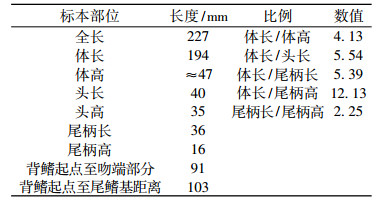
表 1 鲃亚科(属、种未定)测量 Tab. 1 Measurement of Barbinae gen. et sp.indet. |
|
|
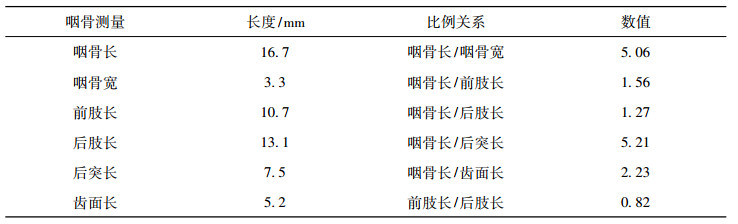
表 2 鲃亚科(属、种未定)右侧咽骨测量 Tab. 2 Measurements of right pharyngeal bone of Barbinae gen. et sp. indet. |
材料 一较完整的正负面保存的鱼体化石NWUV1487—1488(西北大学新生代地质与环境研究所标本编号)。
产地和层位 柴达木盆地西北缘, 茫崖; 上干柴沟组底部, 渐新统。
描述 体延长, 稍侧扁, 吻端圆, 鱼体侧面保存, 化石全长(自吻端至尾鳍末端的水平距离)227 mm, 体长(吻端到尾鳍基部最后一枚椎骨末端的水平距离)194 mm。尾柄长为尾柄高的2.25倍。测量见表 1。
头部骨骼 头部骨骼保存不完整(见图 3), 一些骨片已移位。围眶骨(cs)残存; 额骨(fr)相对完整, 一对, 呈长宽比约为3:1的不规则长方形, 背面见眶上感觉管及其开孔; 两额骨中线相接; 额骨后缘与顶骨(ps)相接。顶骨(pa), 一对, 呈长宽比约为1.5:1的长方形, 背面相对平整, 两块顶骨中线相接。镜下可见一细长三角形、尖端有“Y”形分叉并与筛骨相连的骨片, 推测其为上颌骨(ma)。鳃盖骨骨片没有保存, 但骨片轮廓清晰可辨。前鳃盖骨(pop)呈反“L”形, 折点处宽大, 其下方间鳃盖骨部分出露。鳃盖骨(op)的前缘与前鳃盖骨(pop)的后缘腹面相接, 整体略呈方形, 内侧光滑, 背前方向腹后方发出致密的放射纹。此外, 在主鳃盖骨右下方(见图 3pf)可见相对清晰的胸鳍, 部分鳍条未保存, 可见鳍条8个。
咽骨咽齿 在标本NWUV1488内赋存一对咽骨及其附着咽齿, 其中部分出露(见图 4), 部分在岩石内部(见图 5)。对咽齿的观察主要通过光学显微镜及板状化石CT两种手段。在光学显微镜下观察, 出露的咽骨部分为左侧咽骨(见图 4), 其腹侧面清晰可见, 呈不规则“L”形, 后支后端残存, 印痕清晰, 前支被覆盖, 部分保存完好的咽齿出露; 通过CT(见图 4B)可以清晰的看到被覆盖的前肢前端有一咽齿(A1)存在, 其长度小, 齿径小, 顶端尖弯, 主行第二咽齿(A2)最为粗壮。
未出露部分的咽骨为右侧咽骨(见图 5), 其整体细长, 除A1,A2,B3保存欠佳外, 骨骼形态及其它咽齿保存完好, 可见咽齿主行(A行) A2,A3,A4,A5, 中间行(B行)B1,B2,B3和最外行(C行)C1,C2, 齿式/5·3·2。咽齿总体细长, 呈不规则“L”形, 长宽比为5.06, 后肢略长于前肢, 具体测量数据见表 2。其A行A2最为粗壮(见图 5B, C, D), 且与A3之间有三角形空隙(见图 5B), A3,A4,A5细长(见图 5C,D), A行所有咽齿顶部膨胀并有锐尖弯, 指向后肢方向; B行咽齿细长, 略短于A行,顶端膨胀且有锐尖弯, 也指向后肢方向; C行咽齿细长, 略短于B行, 明显短于A行, 顶端略有膨胀且有钝尖弯, 指向与A,B行相对(见图 5C, D)。
躯干部分 胸鳍保存, 略错位, 部分被围岩覆盖, 保存支鳍骨及分节鳍条8个(见图 3pf); 腹鳍鳍条保存尚好, 保存一支鳍骨及分节鳍8。背鳍支鳍骨保存完好, 见7根保存较好的背鳍支鳍骨, 第一个最大, 向后依次变小, 第一支鳍骨略向后上方错位。背鳍起点相对腹鳍起点靠前; 背鳍起点距吻端91 mm, 距尾鳍基部103 mm。在尾鳍支鳍骨的前上方有一粗壮的硬刺(见图 2sf), 推测该硬刺应属于背鳍(从形态上看, 臀鳍的支鳍骨较短小, 尚不足以支撑此硬棘刺)。背鳍硬棘上可见锯齿18个, 但总数应多于此(见图 6C, D)。椎体保存相对完好, 数目为40+4, 其中尾椎骨为22;腹肋骨数目为13~15, 较粗壮。
臀鳍 臀鳍保存有鳍条及印痕5个, 未见粗壮支鳍骨(见图 2af)。
尾骨骼 尾骨骼(见图 6A,B), 保存较好, 正型尾, 分支鳍条被围岩覆盖, 部分出露; 尾上骨(e)1个, 保存印痕。前缘为最末节椎体的髓棘; 尾杆骨(u)为“扇”形, 约呈50°角上翘, 与最后一节椎体末端连接; 尾下骨6个(h1~h6), 第3尾下骨和第4尾下骨之间有一三角形空隙。尾下骨与尾杆骨形似, 呈宽片状, 下端过渡为副尾杆骨(phy)和粗壮的脉棘(hs)。
3 讨论 3.1 系统位置根据目前《中国动物志·硬骨鱼纲·鲤形目》的分类方案, 鲤科鱼类以臀鳍数目分为两大支系, 臀鳍分支鳍条大于5为雅罗鱼系, 臀鳍仅有5条的为鲃系, 且雅罗鱼系背鳍少见硬棘刺[7-8]。在此基础上鲃系又分为鲃亚科、野鲮亚科、裂腹鱼亚科和鲤亚科4个亚科。野鲮亚科的背鳍末根不分枝, 鳍条柔软, 不为硬刺; 鲤亚科的界定存在争论, 笔者认同王幼槐[14]和中国动物志的观点, 以臀鳍是否具硬刺作为鲤亚科和鲃亚科的分类依据。本文材料臀鳍不具备硬刺, 与鲤亚科特征不符[5]。裂腹鱼亚科以臀鳞为显著特征[7, 15-16], 但其与鲃亚科在重要的骨骼特征上没有明显的区别[1]。从上述新发现的标本特征来看, 其咽骨(见图 5)细长, 附着咽齿3行, 主行为5个, 齿尖尖弯, 咀嚼面凹入, 均表现出鲃亚科的原始特征, 对应早期鲃系鱼类进化的原始等级[17];而裂腹鱼亚科中部分属的咽齿行数和主行咽齿数已有减少的特化表现, 例如裸鲤属和裂尻鱼属的咽齿齿式为4·3/3·4 [7-8, 14, 17]。其次, 根据目前可以参考的鱼类资料[8, 18-19], 裂腹鱼亚科中的多数属的背鳍不分枝鳍条,以较细弱或不存在硬刺为主要特征。脊椎数目也是进行分类的标准之一, 就鲃亚科和裂腹鱼亚科来说, 鲃亚科脊椎数目在(36+4)~(42+4)(新标本的椎体数目为40+4), 而裂腹鱼亚科的脊椎数目往往大于42+4[15, 20], 现有的鱼类资料[6-7]也说明了这一点。本研究的化石出自于下干柴沟组和上干柴沟组交界处, 地质年代为古近纪渐新世[21], 有研究认为,此时中国范围内仅存有鲃亚科、(鱼丹)亚科、鲤亚科、鱊亚科[7]。裂腹鱼亚科的起源与青藏高原的隆升直接相关, 渐新世时期该区域尚未明显隆升[22], 研究区此时也可能并不具备裂腹鱼亚科起源的客观条件。综合分析显示, 本文新发现的标本应为鲃亚科鱼类。
中国已发现的鲃亚科,化石地点先后有山东临朐、北京周口店第十四地点[23]、三水盆地[24]、仑坡拉盆地[15]、柴达木盆地[1]等。杨钟健和张春霖(1936)记述的临朐两种中新世鲤科化石臀鳍具有粗壮且带锯齿的硬刺[23], 根据目前普遍认可的分类标准应归入鲤亚科; 刘宪亭(1954)记述的北京周口店第十四地点的早上新世鲤科化石有2属4种鲃亚科,化石仅短头鲃骨骼形态与新材料相似, 但脊椎数目为39, 比新材料脊椎数目少5个, 陈宜瑜(1998)认为,根据现有的分类,其定名标准不一定正确, 但应属于原始的鲃亚科; 李国藩(1979)记述了三水盆地始新世鲃亚科化石3个属, 唇纹鱼属材料和四须鲃属材料的体长/头长均不大于3.5, 突吻鱼属体长/头长值为4, 均小于本文材料的此项比值[22], 也可与新标本区分, 陈宜瑜(1998)和周家健(1991)认为三水盆地记述的鲃类化石保存不好, 定名与归类存在问题; 武云飞(1980)记述了仑坡拉盆地上新世鲤科化石, 将其归入裂腹鱼亚科, 后被归入鲃亚科[7, 25], 但其体长/头长≈3, 而新标本显示的此项比值大于5, 明显不同; 陈耿娇和刘娟(2007)记述了柴达木盆地渐新世鲤科化石, 其根据咽骨咽齿特征将其部分材料暂归入鲃亚科。
现代鲃亚科包括14属61种。从咽齿特征上看, 新标本咽骨较窄, 长宽比值为5.02, 附咽齿三行, 齿式/5·3·2。现代鲃亚科中, 盲鲃属咽齿行数为2, 金线鲃属、短吻鱼属、袋唇鱼属、长臀鲃属齿式均不为/5·3·2, 白甲鱼属咽骨极宽[24], 舟齿鱼属咽骨极窄, 新标本与之相比, 明显不同。从背鳍特征来看, 新标本背鳍前不具平卧倒刺, 末根不分枝鳍条特化为强硬刺, 强锯齿18~20个, 倒刺鲃属背鳍前方具平卧倒刺。现代盲鲃属和裂峡鲃属背鳍不具硬刺,与新材料不同; 从体型和脊椎数目来看, 新标本体延长, 稍侧扁, 体长194 mm, 脊椎数40+4, 属中型鱼类, 可以与现代小型鱼类、体长小于100 mm的小鲃属相区别[26];鲈鲤属背鳍后移[27], 背鳍硬刺末端分节, 脊椎数为(44+4)~(45+4)[28], 似鳡属体型似鳡, 呈棒状, 方口鲃属和结鱼属体颇高, 且方口鲃属脊椎数约为30+4, 结鱼属背鳍硬刺有一半分节柔软,这些均可与新材料相区别[7]。光唇鱼属、圆唇鱼属和四须鲃属的外部形态特征相似[29-30],主要以头部结构相区分。新标本头部保存欠佳, 无法针对头部结构进行细致对比(外部形态的其他主要鉴别特征比较一致),但新材料的体长与头长的比值为5.54, 依《中国动物志·硬骨鱼纲·鲤形目》数据, 以上3属各现生种的体长与头长比值均不大于5。因此, 从目前掌握的证据来看, 新材料与光唇鱼属、圆唇鱼属和四须鲃属3属属征相近, 但又存在一定区别。
综合分析显示, 新材料与已发现的化石属种明显不同, 与现生的鲃亚科各属也有区别, 考虑到新材料的头部骨骼保存欠佳,因此暂将新标本归入鲃亚科(属、种未定)。
3.2 分布现生的鲃亚科在中国的分布区域主要在北纬35°以南, 如贵州长江水系(多鳞四须鲃)[7, 30], 广西、云南(金线鲃)[31], 陕西(中华倒刺鲃)[32]。青藏高原地区海拔高, 仅有高度特化的裂腹鱼家族、爬鳅科(Balitoridae)、高原鳅属(Triplohpysa)以及鮡科(Sisoridae)分布,目前未见其他鱼类(Zhao et al., 2008)[33]。鲃亚科在世界范围也主要分布在北纬35°以南,以非洲、亚洲的东南部分布最广[8, 18, 34-35]。中国鲃亚科的化石分布除本次新发现的柴达木盆地西北缘渐新世的鲃亚科化石以外, 还有鸭湖、伦坡拉等地的材料[1]。中国以外哈萨克斯坦东部有始新世和早渐新世鲃亚科化石的报道, 哈萨克斯坦中部有渐新世中上新世的鲤科化石报道[36]。欧洲地区也有鲃亚科化石报道, 但时代都晚于渐新世, 东南亚地区泰国也有鲃亚科化石的报道[37]。将已有的鲃亚科化石与现生种类分布地点进行比对后发现,现今鲃亚科的分布明显环绕青藏高原, 并趋近于更加温暖湿润的区域, 与早期的分布明显不同。这可能是在青藏高原隆升的过程中, 鲃亚科由于不能适应高寒气候逐步退出青藏高原[2]或特化为某种能够适应青藏高原特殊气候的类群。张弥曼等(2016)认为, 原始裂腹鱼亚科鱼类和鲃亚科鱼类在高原形成之前的分布比现在更广, 它们很可能遍及中亚各地, 和目前分布在高原周边地区的情况不同, 原始等级的裂腹鱼亚科鱼类或鲃亚科鱼类那时分布到青藏高原中部地区, 甚至分布到目前环境条件不允许鱼类生存的地区。新标本鱼化石产地为柴达木西北缘, 现今属高原大陆性气候, 以干旱为主要特点, 年降水量自东南部的200 mm递减到西北部的15毫米, 这里的盐湖大都已经干涸, 仅个别低地有残余盐湖[38], 海拔高度在3 400~3 600 m, 未见有淡水鱼类分布记录[8, 18, 39]。就化石特点及其沉积环境而言, 分布的海拔高度范围为1 250~2 500 m, 当时鱼体生存在相对稳定的淡水湖泊环境中, 有稳定、较为丰富的食物供给。由于受到水域的严格限制, 化石鱼类能够比较准确地反映当时的气候环境[3, 6, 40-42]。因此, 本次在柴达木盆地西北边缘渐新统首次发现的鲃亚科化石材料可作为上述古环境变迁的最新证据。
致谢
在野外考察及论文撰写过程中, 得到了中国科学院古脊椎动物研究所张弥曼院士、陈耿娇研究员、吉林大学全成教授的悉心指点和帮助; 中国科学院古脊椎动物与古人类研究所侯叶茂工程师、尹鹏飞助理工程师、西北大学地质学系张宏发、朱坤显高级工程师、贲杭生技师、研究生谢坤、李佳佳等为化石的检测、修复及素描提供了帮助,在此谨致谢忱!
| [1] |
CHEN G J, LIU J. First fossil barbin (Cyprinidae, teleostel) from oligocene of Qaidam Basin in Northern Tibetan Plateau[J]. Vertebrata Palasiatica, 2007, 45(4): 330-341. |
| [2] |
NING W, CHANG M M. Pliocene cyprinids (Cypriniformes, Teleostei) from Kunlun Pass Basin, northeastern Tibetan Plateau and their bearings on development of water system and uplift of the area[J]. 中国科学:地球科学, 2010, 53(4): 485-500. |
| [3] |
张弥曼, MIAOD S. 青藏高原的新生代鱼化石及其古环境意义[J]. 科学通报, 2016, 61(9): 981-995. |
| [4] |
CHEN G J, CHANG M M. A new early cyprinin from Oligocene of South China[J]. 中国科学:地球科学, 2011, 54(4): 481-492. |
| [5] |
CHANG M, WANG X, Liu H, et al. Extraordinarily thick-boned fish linked to the aridification of the Qaidam Basin (northern Tibetan Plateau)[J]. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(36): 13246-13251. DOI:10.1073/pnas.0805982105 |
| [6] |
WANG N, WU F. New Oligocene cyprinid in the central Tibetan Plateau documents the pre-uplift tropical lowlands[J]. Ichthyological Research, 2015, 62(3): 274-285. DOI:10.1007/s10228-014-0438-3 |
| [7] |
中国科学院中国动物志委员会. 中国动物志硬骨鱼纲鲤形目中卷[M]. 北京: 科学出版社, 1998.
|
| [8] |
中国科学院中国动物志委员会. 中国动物志硬骨鱼纲鲤形目下卷[M]. 北京: 科学出版社, 2000.
|
| [9] |
WINFIELD I J, NELSON J S. Cyprinid Fishes[M]. Berlin: Springer Netherlands, 1991.
|
| [10] |
孟庆闻. 鱼类比较解剖[M]. 北京: 科学出版社, 1987.
|
| [11] |
WANG X, QIU Z, LI Q, et al. Vertebrate paleontology, biostratigraphy, geochronology, and paleoenvironment of Qaidam Basin in northern Tibetan Plateau[J]. Palaeogeography Palaeoclimatology Palaeoecology, 2007, 254(3): 363-385. |
| [12] |
孙崇仁. 青海省岩石地层[M]. 北京: 中国地质大学出版社, 1997.
|
| [13] |
张跃中, 张敏, 马富强, 等. 柴达木盆地西部阿尔金斜坡地区盆山演化与油气勘探[J]. 中国石油勘探, 2006, 11(6): 26-32. DOI:10.3969/j.issn.1672-7703.2006.06.005 |
| [14] |
王幼槐. 中国鲤亚科鱼类的分类、分布、起源及演化[J]. 水生生物学报, 1979, 3(4): 419-438. |
| [15] |
武云飞, 陈宜瑜. 西藏北部新第三纪的鲤科鱼类化石[J]. 古脊椎动物与古人类, 1980, 18(1): 15-20. |
| [16] |
武云飞. 青藏高原鱼类[M]. 成都: 四川科学技术出版社, 1992.
|
| [17] |
CHU Y T. Comparative studies on the scales and on the Pharyngeals and their teeth in Chinese cyprinids, with particular reference to taxonomy and evolution[D].Shanghai: Biol Bull St John's Univ, 1935.
|
| [18] |
伍献文. 中国鲤科鱼类志[M]. 上海: 上海科学技术出版社, 1982.
|
| [19] |
CHEN G J, WEI L, LEI X Q, et al. First fossil cobitid (Teleostei: Cypriniformes) from Early-Middle Oligocene deposits of South China[J]. Vertebrata Palasiatica, 2015, 53(4): 299-309. |
| [20] |
WANG Y M, TANG W Q. The number of vertebrae from Chinese species of the Cypriniformes and Its ecological adaptation[J]. Chinese Journal of Zoology, 2014, 49(1): 1-12. |
| [21] |
马新民, 刘池洋, 罗金海, 等. 柴达木盆地上干柴沟组时代归属及代号变更建议[J]. 现代地质, 2014, 28(6): 1266-1274. DOI:10.3969/j.issn.1000-8527.2014.06.018 |
| [22] |
LI J J, FANG X M, MA H Z. Late Cenozoic intensive uplift of Qinghai-Xizang Plateau and its impacts on environments in surrounding area[J]. Quaternary Sciences, 2001, 21(5): 381-391. |
| [23] |
刘宪亭. 周口店第十四地点鱼化石[M]. 北京: 中国科学院出版社, 1954.
|
| [24] |
信强.广义白甲鱼属的物种分类整理及其种间系统发育关系分析[D].武汉: 中科院水生生物研究所, 2008. http://www.irgrid.ac.cn/handle/1471x/207093
|
| [25] |
李国藩, 王将克. 广东三水盆地早第三纪鱼化石种属记述[J]. 中山大学学报(自然科学版), 1979, 26(2): 43-52. |
| [26] |
单乡红. 中国小鲃属的分类整理及小鲃属的分类学讨论(鲤形目:鲤科:鲃亚亚科)[J]. 动物分类学报, 2000, 25(1): 114-115. DOI:10.3969/j.issn.1000-0739.2000.01.020 |
| [27] |
陈世骧. 进化论与分类学[M]. 2版. 北京: 科学出版社, 1987.
|
| [28] |
崔桂华, 褚新洛. 鲤科鱼类鲈鲤的亚种分化和分布[J]. Zoological Systematics, 1990, 5(1): 118-123. |
| [29] |
袁乐洋.光唇鱼属单系性、亲缘关系和分类整理[D].北京: 中国科学院水生生物研究所, 2009. http://ir.ihb.ac.cn/handle/342005/12230?mode=full
|
| [30] |
武云飞, 谭齐佳. 青藏高原鱼类区系特征及其形成的地史原因分析[J]. 动物学报, 1991, 37(2): 135-152. DOI:10.3321/j.issn:0001-7302.1991.02.001 |
| [31] |
LI W X. A new species of the genus sinoycocheilus living in cave from Shilin, Yunnan, China(Cypriniformes, Cyprinidae)[J]. Acta Zootaxonomica Sinica, 2007, 32(1): 226-229. |
| [32] |
李仲辉, 许涛清. 陕西鲃亚科鱼类咽骨咽齿的研究[J]. 河南师范大学学报(自然科学版), 1987, 54(2): 89-92. |
| [33] |
赵亚辉, 张洁, 张春光. 青藏高原鱼类的多样性[J]. 生物学通报, 2008, 43(7): 8-10. DOI:10.3969/j.issn.0006-3193.2008.07.003 |
| [34] |
BIANCO P G. Diversity of Barbinae fishes in southern Europe with description of a new genus and a new species (Cyprinidae)[J]. Italian Journal of Zoology, 1998, 65(1): 125-136. |
| [35] |
RAINBOTH W J. Cyprinids of South East Asia[M]. Berlin: Springer Netherlands, 1991.
|
| [36] |
SYTCHEVSKAYA E K. Paleogene freshwater fish fauna of The USSR and Mongolia[M].Moscow: Joint Soviet-Mongolian Paleont Exped Trans, 1986.
|
| [37] |
CAVENDER T M. The fossil record of the Cyprinidae[M]. Berlin: Springer Netherlands, 1991.
|
| [38] |
杜庆. 初探青海湖地区生态环境演变的起因[J]. 生态学报, 1990, 10(4): 317-322. DOI:10.3321/j.issn:1000-0933.1990.04.013 |
| [39] |
卢娜, 金晓媚. 柴达木盆地植被覆盖分布规律及影响因素[J]. 人民黄河, 2015, 37(1): 94-98. DOI:10.3969/j.issn.1000-1379.2015.01.024 |
| [40] |
周家健. 山东山旺中中新世鲤科化石[J]. 古脊椎动物学报, 1990, 28(2): 95-127. |
| [41] |
CHANG M M. Discovery of fossil Nemacheilids (Cypriniformes, Teleostei, Pisces) from the Tibetan Plateau, China[J]. 中国科学:地球科学, 2012, 55(5): 714-727. |
| [42] |
WU F X, MIAO D S, CHANG M M, et al. Fossil climbing perch and associated plant megafossils indicate a warm and wet central Tibet during the late Oligocene[J]. Sci Rep, 2017, 7(1): 1-7. DOI:10.1038/s41598-016-0028-x |