 2016, Vol. 37
2016, Vol. 37


Plexins是轴突导向因子Semaphorins家族的受体[1]。在脊椎动物体内发现Plexins家族共有9个成员。Plexin A1是多个Semaphorins亚家族的受体,其在神经系统形成、胚胎发育、血管生成、免疫反应方面的作用已有研究,但Plexin A1在生殖系统中的作用尚不明确。本实验从蛋白和mRNA水平研究了Plexin A1在小鼠卵巢不同发育阶段的表达情况,为进一步探讨Plexin A1在调控卵泡生长和卵巢周期性血管生成中的作用奠定基础。
1 材料与方法 1.1 实验动物C57BL/6j小鼠购自华中科技大学同济医学院实验动物中心。不同年龄小鼠卵巢内卵泡的发育阶段不同。3 d乳鼠的卵巢中主要为始基卵泡,5 d可见初级卵泡,7 d次级卵泡形成,2周的小鼠其卵泡大多发育至窦前卵泡阶段,生后3周的小鼠为性成熟前期,窦状卵泡开始发育。选择3 d、5 d、7 d、2周、3周龄小鼠,共设不同年龄段组别5组,每组10只小鼠。
1.2 免疫组织化学方法测定Plexin A1蛋白的表达以SP法行免疫组织化学标记。一抗为羊抗鼠Plexin A1多克隆抗体购自R & D公司,兔抗山羊SP试剂盒购自北京中杉金桥生物医药有限公司,显色试剂盒DAB KIT购自北京中山生物技术有限公司。实验设阴性对照组以PBS代替一抗作用。
1.3 real-time PCR方法检测Plexin A1 mRNA水平的表达PubMed中小鼠Plexin A1基因序列号NM_008881.2,RTPrimerDB中小鼠Plexin A1引物PrimerBank ID253683485b2,由Invitrogen公司合成。另以3-磷酸-甘油脱氧酶(GAPDH)为内参。Plexin A1引物序列:上游5′-GACAGGAGAGGTATATGTTGGCG-3′,下游5′-GGGGTGGGTAGCATTTCTCA -3′,扩增产物长度为119 bp。GAPDH引物序列:上游5′-TGTGTCCGTCGTGGATCTGA-3′,下游5′-TTGCTGTTGAAGTCGCAGGAG -3′,GAPDH扩增产物长度为150 bp。两对引物在连续荧光检测系统(BIO-RAD 96孔real-time PCR仪)中扩增。扩增反应体系20 μl,95 ℃预变性5 min,95 ℃变性15 s,59 ℃退火20 s,72 ℃延伸30 s,共40个循环。反应完毕后从95-60 ℃,每下降0.5 ℃记录一次荧光强度做熔解曲线。每一样本设3个复孔。
1.4 结果判断 1.4.1 免疫组织化学结果判断参照Axiotis标准进行半定量评分。染色强度分值标准:细胞完全不着色记0分,浅黄色记1分,棕黄色记2分,深染黄褐色记3分。阳性细胞占所有同类型细胞百分比分值标准:着色细胞数少于细胞总数10%记0分,细胞总数11%-40%的细胞有阳性着色记1分,41%-70%的细胞有阳性着色记2分,71%以上的细胞有阳性着色记3分。记分结束后将两项评分相加,总分0-1分则染色为(-),2分则为(+)弱阳性,3-4分为(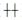
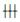
Plexin A1的cDNA从基线水平进入PCR指数增长期的起始点所对应的循环次数为循环阈值(Cycle threshold,Ct),以每个样本在同一批反应中的GAPDH为内参,计算样本△Ct值,再以3 d小鼠卵巢组的△Ct值为参照,计算其它各实验组△△Ct值,根据公式计算Plexin A1 mRNA相对表达量。△Ct=Ct Plexin A1-Ct GAPDH;△△Ct=△Ct实验组-△Ct3d组;mRNA相对表达量=2-△△Ct。
1.5 统计学分析数据分析采用SPSS 19.0软件,进行ANOVA检验和Kruskal Wallis检验,P<0.05为差异有统计学意义。
2 结果 2.1 免疫组化结果不同年龄阶段小鼠卵巢中Plexin A1蛋白的表达水平见图 1,表 1、2。Plexin A1蛋白在各级卵泡的卵母细胞和颗粒细胞中均有表达。随着卵泡的不断发育,卵母细胞和颗粒细胞中Plexin A1蛋白的表达水平均逐渐升高。始基卵泡卵母细胞中的Plexin A1蛋白水平显著低于初级、次级和窦状卵泡的卵母细胞。初级卵泡卵母细胞中的Plexin A1蛋白表达水平低于次级和窦状卵泡的卵母细胞。

|
图 1 Plexin A1在卵泡不同发育阶段的表达 A:3 d;B:5 d;C:7 d;D:2周;E、F:3周。始基卵泡(红色箭头),初级卵泡(黄色箭头),次级卵泡(绿色箭头),窦状卵泡(紫色箭头),闭锁卵泡(蓝色箭头)。O:卵母细胞,G:颗粒细胞,T:卵泡膜细胞。阳性染色为黄色 |
| 表 1 Plexin A1蛋白在小鼠颗粒细胞的表达(n/N) |

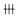
| 表 2 Plexin A1蛋白在小鼠颗粒细胞的表达(n/N) |


始基卵泡颗粒细胞中未见明显Plexin A1蛋白表达,初级卵泡阶段可见其表达,但表达水平显著低于次级及窦状卵泡。而闭锁卵泡中的Plexin A1呈强阳性表达,明显高于其他各级卵泡。
2.2 real-time PCR结果不同年龄阶段小鼠卵巢中Plexin A1 mRNA的表达水平见图 2。5组间两两比较,3 d与2周小鼠卵巢表达水平无显著差异,余各组间两两比较均有显著差异,且其在卵巢组织中的表达水平随周(日)龄呈先升高后降低的趋势,5 d、7 d、3周小鼠卵巢中Plexin A1 mRNA表达水平分别为3 d小鼠的1.56, 1.26, 0.79倍(P<0.05)。

|
图 2 Plexin A1 mRNA在不同年龄小鼠卵巢中的表达 与3 d、2周相比,*P<0.05 |
Plexins在脊椎动物体内共发现有9个成员,分为A-D四个亚家族[1]。Plexins的基本序列在各成员及进化过程中高度保守。最早对Plexins的研究集中在神经系统发育过程,近来发现Plexins和Semaphorins在血管生成[2]、心肺肾骨等器官的发育[3]、免疫反应[4]、肿瘤发生和转移[5]方面也发挥着作用。Plexins与Semaphorins相互作用,可促进细胞的增殖和黏附、细胞骨架的重塑以及新生血管的生成。这些生物学行为在卵泡发育中则表现为颗粒细胞增殖、形态由扁平至立方、细胞层数增加;卵泡直径增大,获得窦腔形成窦状卵泡并最终排卵以及周期性的血管形成。本文首次对小鼠卵巢不同发育阶段中Plexin A1蛋白及mRNA的表达情况进行了探讨。
Plexins是Semaphorins的细胞表面受体,两者结合后向细胞内传导信号可影响轴突导向、神经细胞凋亡、器官形态发生、血管生成和肿瘤侵袭转移[1-5]。研究证实Plexin A1与血管内皮生长因子受体(VEGFR2)可组成受体复合物,与Sema6D结合后增强血管内皮生长因子(VEGF)诱导的VEGFR2的磷酸化,从而促进鸡胚圆锥动脉干处内皮细胞的迁移,促进血管生成[6]。
卵泡发育是一个复杂精密的过程。卵泡经过一系列形态学变化,按始基卵泡、初级卵泡、次级卵泡、窦状卵泡逐级发育,最后成为成熟卵泡并排卵。卵巢血管周期性形成的过程与卵泡发育密切相关[7]。青春期前卵巢无独立的血管网,其始基卵泡的血供来源于在卵泡周围散在分布的间质血管,而在青春期以后,始基卵泡开始发育。当卵泡开始出现窦腔时,卵泡膜细胞层开始有微血管网出现,当卵泡继续向窦状卵泡发育时,微血管网密度不断增加,而优势卵泡的选择则有赖于微血管网密度和血管壁通透性的增加,为其提供足够的血供以继续发育成熟并排卵。我们研究发现Plexin A1蛋白在整个卵泡发育过程中均有表达,主要表达于各发育阶段卵泡的卵母细胞和颗粒细胞胞质中,且随着卵泡的发育,Plexin A1蛋白的表达逐渐增强,其在窦状卵泡中表达为强阳性。这提示Plexin A1在卵泡发育过程中发挥着极为重要的作用,其作用可能是通过调节卵泡血管形成而实现。Dalhia[8]研究了参与血管形成最重要的血管生成因子VEGF及其受体VEGFR2在卵泡发育过程中的表达,发现VEGF的弱阳性开始出现在窦前卵泡和早期窦状卵泡的颗粒细胞和卵泡膜细胞的胞质中,而随着卵泡的进一步发育成熟,卵丘颗粒细胞也表达VEGF,在排卵前,颗粒细胞VEGF的表达呈中等强度,而卵泡膜细胞则为VEGF强阳性。VEGFR2在窦前卵泡的颗粒细胞中无表达、卵泡膜细胞中低表达,早期窦状卵泡的颗粒细胞和卵泡膜细胞的胞质中则有VEGFR2的中等强度表达,排卵前卵泡中强阳性表达。本实验研究发现Plexin A1蛋白与VEGF及其受体有类似的表达模式,推测在卵泡发育过程中,Plexin A1与VEGFR2可能组成受体复合物,调控血管周期性的生长。这一推测在胃癌组织中得到过验证[9]:胃癌组织中Plexin A1的表达高于正常胃黏膜,其表达水平随微血管密度的增高而升高,即肿瘤组织中有较多的新生血管生成,同时伴随Plexin A1表达的增高。但其在卵巢中的作用模式有待进一步研究。
本实验发现Plexin A1蛋白在闭锁卵泡中的表达明显高于生长卵泡,提示Plexin A1可能在卵泡的闭锁过程中发挥一定的作用。卵泡发生闭锁的早期表现即有内皮细胞DNA合成功能的减退和卵泡血管的减少[8]。这一现象提示Plexin A1可能通过其他信号通路作用于卵泡闭锁,以抵消其促进血管生成的作用。各个级别的卵泡均有可能发生闭锁,而卵母细胞凋亡启动的卵泡闭锁是一个极为重要的机制[10]。Simona等[11]研究发现Sema3A可通过Plexin A1/neuropilin-1受体复合物作用于细胞骨架,使Fas转位至脂筏微结构域后,再与其受体FasL接触,引起脂筏结构的破坏。因此,我们推测在卵巢组织中,Plexin A1可能作为共受体传导Fas死亡受体信号,引起卵母细胞凋亡,进而导致卵泡的闭锁。
本实验在mRNA水平的研究也证实了Plexin A1在卵巢组织中的表达。卵巢发育的不同阶段mRNA的表达水平不同,可能与处于不同发育时期的卵泡比例有关。随着小鼠的生长,其始基卵泡数目逐渐减少,初级、次级、窦状卵泡数目逐步上升,总的卵泡数目因为排卵消耗和卵泡闭锁的不断发生而逐渐减少。
卵泡发育的调控是一个复杂的过程,神经内分泌因素和卵泡所处微环境中多种因子均可影响卵泡发育的进程。卵泡发育过程中卵泡募集、优势卵泡选择和各级卵泡闭锁的具体机制仍不完全清楚。本研究提示Plexin A1在卵泡发育微环境中的作用可能有助于阐明卵泡发育的机制,并为体外卵泡培养环境的改善提供新的思路。有关Plexin A1在卵泡发育及闭锁过程中的地位及作用机制尚需进一步的研究。
| [1] | Yazdani U, Terman JR. The semaphorins[J]. Genome Bio, 2006, 7: 211. DOI: 10.1186/gb-2006-7-3-211. |
| [2] | Adams RH, Eichmann A. Axon guidance molecules in vascular patterning[J]. Cold Spring Harb PerspectBiol, 2010, 2: 501. |
| [3] | Perala NM, Immonen T, Sariola H. The expression of plexins during mouse embryogenesis[J]. Gene Expr Patterns, 2005, 5: 355-362. DOI: 10.1016/j.modgep.2004.10.001. |
| [4] | Bismuth G, Boumsell L. Controlling the immune system through semaphorins[J]. Science, 2002, 4: 172-175. |
| [5] | Ch'ng ES, Kumanogoh A. Roles of Sema4D and PlexinB1 in tumor progression[J]. Mol. Cancer, 2010, 9(251). |
| [6] | Toyofuku T, Zhang H, Kumanogoh A, et al. Dual roles of Sema6D in cardiac morphogenesis through region-specific association of its receptor, Plexin A1, with off-track and vascular endothelial growth factor receptor type 2[J]. Genes Dev, 2004, 18: 435-447. DOI: 10.1101/gad.1167304. |
| [7] | Geva E, Jaffe RB. Role of vascular endothelial growth factor in ovarian physiology and pathology[J]. FertilSteril, 2000, 74: 429-437. |
| [8] | Dalhia A, Alejandra RC, Fatima H, et al. Spatiotemporal analysis of the protein expression of angiogenic factors and their related receptors during folliculogenesis in rats with and without hormonal treatment[J]. Reproduction, 2009, 137: 309-320. DOI: 10.1530/REP-08-0130. |
| [9] | Xiang YZ, Lin C, Yu HL, et al. Plexin A1 expression in gastric carcinoma and its relationship with tumor angiogenesis and proliferation[J]. World J Gastroenterol, 2007, 13: 6 558-6 561. DOI: 10.3748/wjg.13.6558. |
| [10] | John J. Oocyte control of ovarian follicular development and function in mammals[J]. Reproduction, 2001, 122: 829-838. DOI: 10.1530/rep.0.1220829. |
| [11] | Simona M, Antonio P, Raffaella L, et al. Semaphorin3A signaling controls Fas (CD95)-mediated apoptosis by promoting Fas translocation into lipid rafts[J]. Blood, 2008, 11: 2 289-2 299. |