 2021, Vol.41
2021, Vol.41


河姆渡文化是长江下游地区的一处新石器时代考古学文化, 时间范围大约为距今7000~6000年[1]。对河姆渡文化动物资源利用方式的研究主要来自于浙江省余姚市两处保存较好的遗址, 分别是20世纪70年代发掘的河姆渡遗址和2004年开始发掘的田螺山遗址。河姆渡遗址共发现哺乳动物34种, 鸟类、鱼类和爬行类共计24种, 超过了田螺山遗址目前记录的18种哺乳动物和6种鱼类[2~3]。由于发掘时间较早, 河姆渡遗址的动物材料大多为头骨和鹿角等鉴定特征最为明确的部位, 而缺少颅后骨。遗址出土了数量较多的猪科和鹿科动物遗存, 研究报告中记载了74件猪上颌骨和172件猪下颌骨, 鹿角的数量超过1400件, 大多较为破碎, 但比较完整的梅花鹿角仍然有40件之多[3]。尽管河姆渡遗址动物遗存的种类以野生动物为主, 20世纪七、八十年代对生业经济的讨论主要集中在猪、狗和水牛驯化, 并有研究者尝试讨论麋鹿是否在当时也已经开始了驯化的历程, 这一研究倾向与当时对农业起源的关注密切相关, 与此相对应的是植物考古领域对河姆渡遗址水稻的粳、籼性状之争[4~5]。21世纪以来, 随着研究的深入, 考古学界对河姆渡文化生业经济的性质进行了反思, 田螺山遗址的发现为重新认识这一问题提供了新的机会。田螺山遗址的发掘应用了科学发掘方法和系统取样, 并进行了多学科综合研究, 从考古年代学、环境考古、植物考古和动物考古多个角度进行研究[6], 揭示出一种渔猎采集为主、作物栽培和动物饲养为辅的经济形态。
渔猎经济是新石器时代长江流域生业经济的重要组成部分[7~8], 但对该地区渔猎经济的内涵仍然缺少系统研究。对田螺山遗址经济形态的复原可以从空间、时间、技术和社会几个层面来考虑。在空间层面上, 主要目的是追溯狩猎和捕鱼的区域范围, 是在居址附近, 还是需要长途跋涉的地点[9], 对狩猎地点的选择一方面反映了人类对经济活动的规划, 另一方面也反应了人类利用与改造自然环境的能力, 需要建立在古环境复原和动物生态研究的基础上。在时间层面, 渔猎活动是否存在季节性的差异, 这也是动物考古学研究中经常讨论的问题[10~13]。日本绳纹时代生业经济的研究对季节性的分析非常具有代表性, 通过整合不同的动物性食物资源(野猪、梅花鹿、淡水鱼、海鱼、海生哺乳动物和贝类等)和植物性资源(果实、种子、花和嫩芽), 绘制出不同季节生业活动的时间表, 又叫做“绳纹日历(Jomon Calendar)”[14]。绳纹人春季采集植物嫩芽, 夏季捕捉各种洄游的海鱼和海生哺乳动物, 秋季采集植物果实与种子, 冬季狩猎野猪和梅花鹿。中岛经夫等[15]对田螺山一个鱼骨坑的研究也认为是季节性捕鱼的结果, 时间可能在春夏之交。渔猎经济的技术层面可以进一步分为具体的捕猎技术(technique)和策略(strategy), 后者包括对猎物个体性别、年龄等特征的选择, 表现出渔猎行为的规划性。通过以上3个角度的分析, 我们可以进一步了解遗址对人力资源的分配和人群的规模, 将对生业经济的研究与社会结构相连, 以达到考古学复原古代社会的目的。
如上所述, 本文采用了新的研究方法, 从空间层面来讨论田螺山遗址的生业经济。研究方法的核心是动物的“生境”, 也就是动物考古学研究中的三大要素人、动物和环境三者交汇的地点, 从而对生业经济有更加立体的认识。
1 生物分类生境指数介绍生境(habitat)又称为栖息地, 是生态学中的常用概念, 指有机体所处的物理环境[16]。Evans等[17]和Andrews[18]在研究化石动物群时, 提出将动物生境进行量化计算, 进而重建古代生态的方法, 即生物分类生境指数(Taxonomic Habitat Index, 简称THI)。
动物的生境可能是单一的也可能是多样的。Evans等[17]和Andrews[18]把某一物种的生境总体视作“1”, 然后衡量每一种生境的重要性, 并赋予其“0”和“1”之间的数值, 保证它们的总和等于“1”, 这些数值便是生物分类生境指数“THI”(见表 1)。按照这种方法, 每个物种的生境都可以拆分成一组THI数值:如果一种动物仅栖息于一种生境, 那么这种生境类型的THI为1, 例如栖息于灌丛生境的黄麂;广适性的动物由于其栖息地多样, THI的赋值须建立在已有的动物生态学研究基础之上。下一步是将研究地点每种生境下的THI相加, 并计算生境间的相对比例, 从而衡量该地区的古代生态分布情况。以下将以英国Westbury化石动物群的研究为例来说明THI方法的应用。
| 表 1 英国Westbury洞穴遗址动物群的种类构成及生境赋值[18] Table 1 Species and Taxonomic Habitat Index(THI)weighing of the Westbury fauna, UK[18] |
Westbury位于英国西南部, 是一处中更新世的石灰岩洞穴化石地点。该地点时间跨度长, 化石种类丰富且构成较为复杂。动物群主要由小型哺乳动物构成, 也包括少量爬行类和鸟类。其中小型哺乳动物大多为现生种, 如小鼩鼱(Sorex minutus)、雪兔(Lepus timidus)、红松鼠(Sciurus vulgaris)、欧鼹鼠(Talpa europaea)等, 少量灭绝种也都属于现生属或科, 如水鼩鼱属(Neomys)、田鼠属(Microtus)和鼹科(Talpidae)[18]。表 1统计了Westbury动物群的全部31种动物及其生物分类生境指数的分布。
Westbury化石地点的堆积复杂多样, 动物群的组成也各自不同[18]。例如, Unit 11包含两个特征显著不同的动物群, 其中subunit 11/4动物群的种类在整个Westbury化石地点最为多样化, 而subunit 11/1动物群的种类较少且单一。Unit 15/8动物群的规模最大, 骨骼数超过45000, 最小个体数(MNI)为3826, 包含种类多样, 对于反映古生态有很强的代表性;该动物群也表现出种类分布集中的特征, 90 %以上为松田鼠(Pitymys gregaloides)和普通田鼠/黑田鼠(Microtus arvalis/agrestis)。由于种类和生境的差异, Westbury化石地点各个堆积的THI分布表现出区别, 反映了古生境随着时间的变化[18](图 1)。Unit 10和11仍然代表较为温暖的环境, 表现为落叶阔叶林比值较高的特点, 其中Unit 11到达了间冰期的顶点。Unit 12堆积形成时期, 气候略微变得干冷, 表现为干草原和荒漠生境比值有所回升。回冷趋势在Unit 13和14最为明显, 泰加林和针叶林生境大幅增加, 环境条件与目前斯堪的纳维亚半岛南部类似。Unit 15堆积形成时期的环境气候变化较大, 表现为各个subunit之间的波动。在Unit 15形成的前期, 气候变得温暖, 表现为subunit 15/1至15/4落叶阔叶林生境大幅回升, 泰加林、针叶林、干草原和荒漠等生境回落, 其中subunit 15/4基本达到Unit 11的顶点水平, 然后继续回冷, Unit 15/8变得更为干冷, 与现在斯堪的纳维亚半岛北部相似, 甚至接近冰期的水平。

|
图 1 Westbury化石地点西侧断崖堆积第10至第15单元THI分布图(数据来源Andrews[18]) Fig. 1 The THI spectra for the faunas from Units 10-15 of the Western Bluff sequence, Westbury(original data from Andrews[18]) |
从生物分类生境指数方法的分析过程可以看出, 这种方法旨在通过分析动物群的所有种类来讨论古代生态, 而非仅仅依靠少数指示物种(indicator species), 克服了动物群种类不等于指示种的问题。与之相似, 考古遗址动物群作为人类活动的遗存, 以广适性的动物为主, 也缺少指示物种, 因此THI拆分动物生境的方法对于讨论古代人类与生态环境之间的关系具有重要意义。
2 材料与方法 2.1 遗址背景与材料田螺山遗址位于浙江省余姚市三七市镇相岙村(30°01′27″N, 121°22′46″E), 距离河姆渡遗址直线距离仅7 km。遗址地处宁绍平原姚江流域的一处小山谷内, 当前海拔高度只有5 m[19]。自2004年发现以来, 田螺山遗址出土了木结构建筑、食物储藏坑、墓葬、古稻田等多种遗迹现象, 以及大量河姆渡文化的遗物, 碳十四年代约为5000~4000 cal.BC[20]。动植物遗存数量巨大且保存良好, 为进一步研究河姆渡文化生业经济提供了重要材料。
本文选取田螺山遗址2004年至2010年间发掘的哺乳动物和鱼类遗存作为研究对象[2]。这批材料均出自河姆渡时期文化层, 并根据研究者对地层的划分进一步分为早、中、晚期3个阶段进行统计[19]。地层的堆积均经过湿筛, 哺乳动物遗存主要通过较为细致的手选法获得, 共计20185件;鱼类遗存均在过筛后拣选获得, 总数为178403件。所有动物骨骼均按照出土单位逐一鉴定、测量、记录观测到的信息并录入数据库。哺乳动物种属的鉴定主要参考北京大学考古文博学院标本室所藏现生标本, 也参考了《动物骨骼图谱》等文献[21~22];鱼类种属和部位的鉴定主要根据笔者制作的现生鱼类比对标本。定量统计方法采用了动物考古学常用的可鉴定标本数(Number of Identified Specimens, 简称NISP)和最小个体数(Minimum Number of Individuals, 简称MNI)[23~24]。
20185件哺乳动物遗存中, 可鉴定标本数(NISP)为6378, 包括18种动物, 名称与相对比例见表 2。所有动物中, 狗(Canis familiaris)是明确的驯化动物, 相关研究认为此时已经出现了猪的饲养[25], 其余均为野生动物。偶蹄类动物是主要的食物来源, 其中数量最多的是梅花鹿(Cervus nippon), 超过总数的1/3, 其次为黄麂(Muntiacus reevesi), 猪类(包括家猪和野猪)的比例在10 %左右。鱼类遗存以6种淡水鱼类为主, 分别是鲤鱼(Cyprinus capio)、鲫鱼(Carassius auratus)、鲌鱼(Culter sp.)、鲶鱼(Silurus sp.)、乌鳢(Channa argus)和鲈鱼(Lateolabrax sp.), 其中以乌鳢、鲫鱼的数量为最多, 其次是鲤鱼(表 3), 它们的共同特征是对环境的适应性强, 尤其是对缺氧水体具有一定的耐受性[26~27]。
| 表 2 田螺山遗址哺乳动物可鉴定标本数(NISP)及比例 Table 2 The NISP and proportions of mammalian taxa from Tianluoshan |
| 表 3 田螺山遗址鱼类可鉴定标本数(NISP)及比例 Table 3 The NISP and proportions of fish taxa from Tianluoshan |
在植物考古和环境考古研究的基础上, 参考动物的习性和生活史, 我们将田螺山遗址的环境分为6种生境:林地(forest)、灌丛(shrub)、草地(grassland)、湿地(wetland)、河湖(river & lake)、和海岸(sea shore), 大致分布如图 2所示。其中高大乔木(亚热带常绿阔叶林)构成的林地主要分布在遗址以南二、三十公里外的四明山海拔较高的区域, 在六、七千年前气候较为温暖湿润的时期, 这里可能是大型木材的来源[28]。灌丛指低矮丘陵上以灌木为主的植被, 栖息有梅花鹿、黄麂、野猪等有蹄类动物;湿地指遗址周围大面积的浅水水域, 生长有菱角、芡实、莎草等湿地植物;草地指位于湿地和灌丛之间较为干燥的区域, 生长有茂密的草本植物;河湖指河流、湖泊等大型水体, 与湿地之间可能相连, 所以湿地和河湖的动物组成有一定的相似性。由于分析对象为哺乳动物, 而且分析的遗存中没有出现海洋哺乳动物, 用“海岸”指代距离海洋比较近的陆地生境。

|
图 2 田螺山遗址哺乳动物和鱼类HEI值分布图 Fig. 2 The HEI distribution of the Tianluoshan, mammals and fish calculated separately |
对每种动物THI的设置参考了动物志、已经发表的动物生态学研究以及网络动物科学数据库[29~31]。动物生态调查中会设置一系列样方, 记录样方的植被型、郁闭度、坡向、水源距离等生态因子, 以动物的粪便、足迹和卧迹等新鲜活动痕迹作为是否作为动物栖息地的依据[32]。下面以几种鹿类动物为例说明。小型鹿类动物中, 黄麂个体小、行动灵活, 灌丛是它们的唯一生境, 既为它们提供了橡果等食物来源, 低矮多刺的丛林也为它们提供了躲避天敌的条件[30]。因此, 黄麂的THI设置中, “灌丛”的赋值为1, 其余生境为0(表 4)。哺乳动物中, 梅花鹿对环境的适应性较强, 因此在为不同生境赋值时主要参考了与田螺山遗址梅花鹿同为华南亚种的江西桃红岭自然保护区和浙江清凉峰自然保护的梅花鹿种群[32~33], 综合了不同季节梅花鹿对栖息地选择的研究。所有的研究都表明, 梅花鹿喜欢在草甸-沼泽和灌丛栖息地中活动, 不喜欢阔叶林[33]。由于相关研究中没有梅花鹿在不同生境出现概率的具体数据, 而且灌丛下层也分布有大量草本植物, 研究中灌丛和草甸往往同时出现, 于是给二者的赋值数据相当;据记载, 梅花鹿在繁殖季节会偏爱茅草丛, 因此, 草地的THI赋值(0.5)略高于灌丛(0.4);有的研究提到梅花鹿会在接近水源的地区活动[33], 因此湿地有0.1的赋值(表 4)。总之, 本文对生境的赋值时建立在对大量文献的阅读和估算上, 虽然难以避免有主观差异, 但数据仍然能够反映动物生境的分布范围以及对生境选择的倾向性。数字是反映分布特征的手段, 在引入MNI统计数据之后, 会达到强调分布趋势和降低主观影响的作用。
| 表 4 田螺山遗址哺乳动物生物分类生境指数(THI)赋值与最小个体数(MNI)统计 Table 4 Taxonomic Habitat Index(THI)weighing and Minimum Number of Individuals(MNI) of the mammalian species from Tianluoshan |
Evans等[17]和Andrews[18]的THI方法建立在对化石地点的研究上, 其成因主要是自然因素。与之不同的是, 考古动物遗存是人类生业活动的结果, 它们的生境代表了人类渔猎活动所到达的地点, 而不单纯是古环境。因此, 在将THI方法应用到动物考古中时, 它所反映的含义会相应地发生变化, 代表了人类对动物生境的开发利用情况。
如果将THI方法照搬到考古遗址中, 便会出现一个问题:动物种类不论数量多少都被同样对待, 这样会导致少数种类的重要性被夸大。动物个体数在动物考古中具有重要意义, 反映了不同动物资源在生业经济中的重要程度, 换算到空间层面, 动物个体数也反映了人类对这些动物栖息地环境利用程度的强弱差异。因此, 本研究将THI与MNI进行了整合, MNI值代表了对生境利用的频次, MNI值越大, 表示对这类生境的利用强度越大。通过引入MNI, 将研究目标从复原古代生态转化成对人类利用环境与资源的探讨, 从而将环境考古和生态学的研究结果与动物考古学研究结合起来, 充分发挥多学科交流的优势。计算方法用如下公式表示:
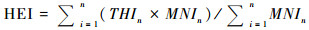
|
其中HEI指Habitat Exploitation Index, 用来反映对生境利用的情况;所有生境的HEI之和为1。n代表动物物种, 分母则表示最小个体数之和。
田螺山遗址早、中、晚期的动物MNI分布如表 4所示。表 5显示的是THI与对应的MNI相乘的结果将各个生境的THI值与MNI相乘得出的数值, 然后每一列相加得到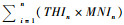
| 表 5 田螺山遗址早、中、晚期THIn×MNIn与HEI值统计 Table 5 Calculating the THIn×MNIn and HEI values in the Early, Middle, and Late phases of Tianluoshan |
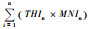
通过对田螺山遗址哺乳动物和鱼类所代表HEI值的计算, 最终得到图 2所示的分布模式。哺乳动物的猎获主要来自于灌丛, 其次为草地和湿地;林地的HEI值反而很低, 主要原因是数量最多的梅花鹿和黄麂都不进入林地。从时间上看, 遗址早、中、晚期的HEI分布变化不大, 仅仅略有起伏, 灌丛始终是最主要的狩猎地点;晚期草地的THI值略有升高, 可以与孢粉学的研究结果相呼应, 可能与气候转凉以及人类活动影响到植被分布有关[34]。
鱼类资源主要来自于湿地, 河流和湖泊等淡水水体的贡献相比之下要小许多, 海洋环境的贡献几乎可以忽略不计。
4 讨论 4.1 田螺山遗址动物资源利用的空间分布环境考古和植物考古研究对田螺山遗址和宁绍平原一带的古代植被进行了复原[35~38], 具体描述如下。遗址周围存在着大面积的湿地, 生长着菱角(Trapa)、芡实(Euryale)、荇菜(Nymphoides)等水生植物, 近水边生长着黍属(Panicum)、稗属(Echinochloa)、野葡萄(Vitis)等草本植物。丘陵山脚地带和山上虽然都为亚热带落叶阔叶林, 但是植物的构成根据海拔的高低有所不同。海拔100 m以上的混交林以常绿栎树为主, 如石栎属(Lithocarpus)和青冈属(Cyclobalanopsis)。河谷边缘和低矮山脚地带是湿地-草地向林地的过渡地带, 分布的亚热落叶阔叶混交林以灌木和小乔木为主, 林间生长着悬钩子属(Rubus)和甜瓜属(Cucumis)等草本植物, 该地区便是本文分类中的灌丛生境。遗址所在河谷以南的四明山麓分布有亚热带落叶阔叶林, 海拔较高的区域则有亚热带落叶阔叶和针叶混交林, 遗址中出现的樟树、柏树和银杏便来自于这一带。
遗址周围地形复杂, 在河谷的小范围内, 生境种类多样, 以遗址为中心向外, 依次分布有湿地、草地、灌丛和河流几种主要的生境类型, 在小范围内即可以满足动物多样化的狩猎需求, 其代表性的渔猎中心便是湿地和灌丛。根据学者们对田螺山遗址资源域的研究, 湿地分布在距离遗址1~3小时的活动半径范围内, 而灌丛则在不超过2~4小时活动半径内[38], 来自这部分区域的资源构成了食物的主要组成部分(图 3)。对动物资源的季节性研究表明, 渔猎活动全年进行, 尽管有高峰和低谷, 但在时间上却从无间断[39], 与这些食物资源容易获取的特性是密切相关的。相较之下, 森林生境多分布在距离遗址几十公里之外的四明山麓, 活动距离超过12小时, 甚至高达20小时以上, 并不适合作为日常消费的食物资源的主要来源。

|
图 3 田螺山遗址周围地貌形态及动物资源主要分布范围 Fig. 3 Physiographic setting and major distribution of animal resources surrounding Tianluoshan |
结合动、植物考古的研究可以发现, 植物性食物和动物性食物的来源地是基本重合的。田螺山遗址最早期的单位中出土了大量橡果, 是青冈属和石栎属的果实, 都可以在灌丛和低海拔林地中获得。遗址最主要的食物是菱角、芡实和水稻(Oryza), 都是湿地植物。可见, 湿地和灌丛是整个生业资源空间分布的核心。生业活动也会深入到较远的森林生境, 例如采摘南酸枣(Choerospondias axillaris)等野生果实, 和狩猎某些动物。稳定的食物资源及其空间分布的便利性可能是田螺山村落延续使用了上千年的原因之一。
4.2 湿地环境在生业经济中的作用湿地是田螺山遗址稳定食物来源的核心。湿地为多种鸟类(鸭、大雁等)、爬行类(龟、鳖、短吻鳄等)和两栖类(蛙类等)提供了栖息环境, 也是一些哺乳动物的栖息地之一, 包括麋鹿、水牛和水貂等依赖于水源的动物, 梅花鹿和獐也会到湿地附近活动[30, 40~41]。从生态学的角度, 另外很重要的一点是, 湿地是多种淡水鱼类的繁殖场所, 大量的挺水植物和沉水植物为鱼类产卵及幼鱼的生长提供庇护, 很多鱼类会因此在湿地与平时的栖息地之间季节性地洄游[41~42]。湿地植物的果实也可以食用。可见对人类社会, 湿地是一块资源相对密集的生境。
田螺山遗址的鱼类遗存中的物种组成十分特别, 乌鳢的比例占可鉴定标本数的一半左右甚至更高(表 3), 这在国内外的研究中从未出现过。我国的鱼类考古研究尚处于起步阶段, 长江流域的两处经过系统研究的遗址, 重庆中坝遗址和湖南高庙遗址, 鱼类的组成都是以植食性的鲤科鱼类为主[43~44]。但是乌鳢是一种凶猛的肉食性鱼类, 它的食谱非常广, 昆虫、鱼、蛙都可能成为它们的食物。由于处在食物链顶端, 乌鳢在自然水体中的比例比较低, 据调查, 洪湖的乌鳢比例在1 %至3 %之间[45], 太湖乌鳢等多种数量较少的鱼类总体在5.5 %至19.6 %之间[46]。
研究认为, 乌鳢的高比例可能是环境和人为选择双重因素作用的结果[2, 39]。乌鳢的适应性极强, 既能耐寒又能耐热, 可以在0~30 ℃的环境中生存, 由于身体的特殊结构, 可以直接从空气中呼吸氧气, 因此能够在缺氧的水体中生存;其强适应性还表现在可以在陆地上短距离移动, 以及用休眠的方式度过旱季[27, 47]。这些特征使得湿地环境成为它们理想的栖息地。一般来说, 湿地鱼类的多样性要远远低于河流、湖泊等淡水环境, 其主要限制因素是水体的含氧量低;另外湿地的水位浅、昼夜温差大, 会进一步加剧缺氧性[41~42]。乌鳢可以看作是湿地的代表性动物, 鲶形目鱼类和以鲤鱼、鲫鱼为代表的一些鲤科鱼类对湿地环境也有较强的适应能力。
人类活动对乌鳢比例的影响则主要表现在捕鱼方法上。与大量鱼类遗存形成鲜明对比的是, 田螺山遗址中并没有发现鱼钩、鱼镖和网坠等捕鱼工具, 河姆渡遗址的情况与之类似, 仅发现了两件鱼镖和两件网坠[48]。那么这成千上万条鱼是如何捕获的?根据民族学材料和国内外对捕鱼方法的研究, 捕鱼方法成千上万, 真正能够留下考古学遗存基本只有鱼钩、鱼镖和网坠几种, 特殊的保存条件下一些有机质的工具或许也能保留下来[49]。捕鱼方法是由目标鱼类的行为特征和周围环境的特性决定的。湿地环境水位浅、挺水植物和沉水植物众多, 并不适合钓鱼和网鱼, 所有捕鱼工具中最适合湿地环境的包括鱼篓、罩笼、围坝和徒手[50], 但是这些方法都很难保存下来, 或者会散落在野外不容易被发现。乌鳢表现出很强的行为特点, 包括集群行动和攻击性强。鱼类在繁殖季节会汇集成群, 而这时是捕鱼的理想时期[51~52], 乌鳢在繁殖季节之外也经常成群行动。此外, 乌鳢是一种凶猛的肉食鱼类, 对移动物体会表现出很强的攻击性, 这一特征在繁殖季节会尤为突出, 因此渔民们经常利用乌鳢的这一特点, 用绳子拴上活青蛙作为诱饵来捕鱼[53]。
田螺山遗址也出现了早期的稻作农业。水稻在植物遗存中的比例较低, 是作为一种补充性质的资源存在的[36, 54~55], 考古调查中也发现了水稻田遗迹[56]。从生态的角度讲, 水稻田本质上是一种人工湿地, 具有湿地的生态特征。这个时期的水稻田还比较简单, 只是在湿地的边缘地带对土地进行整治使其变得适合栽种水稻, 水稻田与自然水体仍然相连, 仍然是湿地的一部分。随着稻作农业的发展, 出现了给排水系统, 水稻田不再需要依靠自然湿地, 而是形成了新的生态系统, 不止提供水稻, 也可以作为捕鱼的地点[57~58]。湿地以自然和人工两种形态在生业经济系统中发挥着作用。
5 结论河姆渡文化生业经济的研究对于讨论长江下游地区新石器时代考古学文化的发展和稻作农业的起源具有重要意义。本文分析了田螺山遗址2004年至2010年间发掘的哺乳动物和鱼类遗存, 将已有的生物分类生境指数(THI)分析方法与动物考古学的统计方法(MNI)相结合, 提出了新的HEI指数, 对生境进行量化分析, 以衡量古代人类对多种环境资源的使用情况。这种方法有两个优点:1)将定性分析转为定量分析, 可以直观地了解人类利用环境资源的选择权重;2)THI拆分生境的方法使得我们可以将大量广适性的动物纳入资源利用的空间分析, 而不是完全倚重指示物种。由于同时引用了THI和MNI两个参数, HEI所描述动物资源的空间分布体现出了不同遗址之间生业经济的个性区别, 解决了以往动物考古学中复原古代环境比较相似的问题。
经过分析发现, 渔猎活动对生境有明显的选择性, 狩猎活动主要发生在灌丛至草丛一带, 对有高大乔木的森林地带涉足较少;湿地是主要的捕鱼地点, 造成的一个显著结果是遗址鱼类组成以对低氧环境耐受性强的乌鳢、鲫鱼和鲤鱼为主。通过参考古环境和资源域复原的结果, 狩猎和捕鱼活动主要发生在距离村落不超过4小时的活动半径范围内, 属于日常的生计活动。结合植物考古的结果来看, 湿地是田螺山遗址生业经济的核心区域, 除了日常消费的鱼类之外, 还生产菱角和芡实等食物。更为重要的是, 湿地环境与水稻的起源直接相关, 对于研究新石器时代长江下游地区生业经济形态的发展将有重要意义。
致谢: 感谢审稿专家和编辑部杨美芳老师宝贵的修改意见
| [1] |
中国社会科学院考古研究所. 中国考古学: 新石器时代卷[M]. 北京: 中国社会科学出版社, 2010: 451-460. The Institute of Archaeology, Chinese Academy of Social Sciences. Chinese Archaeology: Neolithic[M]. Beijing: China Social Sciences Press, 2010: 451-460. |
| [2] |
Zhang Ying. Animal Procurement in the Late Neolithic of the Yangtze River Basin: Integrating the Fish Remains into A Case—Study from Tianluoshan[D]. London: The Doctorate Dissertation of University College London, 2015: 1-51.
|
| [3] |
魏丰, 吴维棠, 张明华, 等. 浙江余姚河姆渡新石器时代遗址动物群[M]. 北京: 海洋出版社, 1989: 90. Wei Feng, Wu Weitang, Zhang Minghua, et al. The Fauna from the Neolithic Site at Hemudu, Zhejiang[M]. Beijing: China Ocean Press, 1989: 90. |
| [4] |
游修龄. 对河姆渡遗址第四文化层出土稻谷和骨耜的几点看法[J]. 文物, 1976(8): 20-23. You Xiuling. Study on the rice grains and bone spade from layer 4 of Hemudu[J]. Cultural Relics, 1976(8): 20-23. |
| [5] |
汤圣祥, 张文绪, 刘军. 河姆渡·罗家角出土稻谷外稃双峰乳突的扫描电镜观察研究[J]. 作物学报, 1999, 25(3): 320-327. Tang Shengxiang, Zhang Wenxu, Liu Jun. The study on the bi-peak-tubercle on lemma of Hemudu and Luojiajiao ancient excavated rice grains with Electric Scanning Microscope[J]. Acta Agronomica Sinica, 1999, 25(3): 320-327. DOI:10.3321/j.issn:0496-3490.1999.03.009 |
| [6] |
北京大学中国考古学研究中心, 浙江省文物考古研究所. 田螺山遗址自然遗存综合研究[M]. 北京: 文物出版社, 2011: 1-294. Centre for the Study of Chinese Archaeology in Peking University, Zhejiang Province Institute of Archaeology and Cultural Heritage. Integrated Studies on the Natural Remains from Tianluoshan[M]. Beijing: Cultural Relics Press, 2011: 1-294. |
| [7] |
Yuan J, Flad R, Luo Y. Meat-acquisition patterns in the Neolithic Yangzi River valley, China[J]. Antiquity, 2008, 82(316): 351-366. DOI:10.1017/S0003598X0009685X |
| [8] |
袁靖. 中国动物考古学[M]. 北京: 文物出版社, 2015: 179-180. Yuan Jing. Zooarchaeology of China[M]. Beijing: Cultural Relics Press, 2015: 179-180. |
| [9] |
Reitz E J, Wing E S. Zooarchaeology[M]. Cambridge: Cambridge University Press, 2008: 251-286.
|
| [10] |
Legge A J, Rowley-Conwy P. Star Carr Revisited: A Re-analysis of the Large Mammals[M]. London: Centre for Extra-Mural Studies, 1988: 1-145.
|
| [11] |
Noe-Nygaard N. The importance of aquatic resources to Mesolithic man at inland sites in Denmark[M]//Clutton-Brock J, Grigson C. Animals and Archaeology 2: Shell Middens, Fishes and Birds. Oxford: British Archaeological Reports International Series, 1983: 125-142.
|
| [12] |
van Neer W, Löugas L, Rijnsdorp A D. Reconstructing age distribution, season of capture and growth rate of fish from archaeological sites based on otoliths and vertebrae[J]. International Journal of Osteoarchaeology, 1999, 9(2): 116-130. DOI:10.1002/(SICI)1099-1212(199903/04)9:2<116::AID-OA465>3.0.CO;2-H |
| [13] |
Wheeler A, Jones A K G. Fishes[M]. Cambridge: Cambridge University Press, 1989: 154-161.
|
| [14] |
Habu Junko. Ancient Jomon of Japan[M]. Cambridge: Cambridge University Press, 2004: 61-62.
|
| [15] |
中岛经夫, 中岛美智代, 孙国平, 等.田螺山遗址K3鱼骨坑内的鲤科鱼类咽齿[M]//北京大学中国考古学研究中心, 浙江省文物考古研究所.田螺山遗址自然遗存综合研究.北京: 文物出版社, 2011: 206-236. Nakajima T, Nakajima M, Sun Guoping, et al. Pharyngeal tooth remains from the fish bone pit(K3) at Tianluoshan[M]//Centre for the Study of Chinese Archaeology in Peking University, Zhejiang Province Institute of Archaeology and Cultural Heritage. Integrated Studies on the Natural Remains from Tianluoshan. Beijing: Cultural Relics Press, 2011: 206-236. |
| [16] |
孙儒泳, 王德华, 牛翠娟, 等. 动物生态学原理[M]. 第四版. 北京: 北京师范大学出版社, 2019: 20. Sun Ruyong, Wang Dehua, Niu Cuijuan, et al. Principles of Animal Ecology[M]. fourth edition. Beijing: Beijing Normal University Publishing Group, 2019: 20. |
| [17] |
Evans E M N, Van Couvering J A H, Andrews P. Palaeoecology of Miocene sites in western Kenya[J]. Journal of Human Evolution, 1981, 10(1): 99-116. DOI:10.1016/S0047-2484(81)80027-9 |
| [18] |
Andrews P. Owls Caves and Fossils: Predation, Preservation and Accumulation of Small Mammal Bones in Caves, with An Analysis of the Pleistocene Cave Faunas from Westbury-sub-Mendip, Somerset, UK[M]. London: Natural History Museum Publications, 1990: 107-177.
|
| [19] |
孙国平.田螺山遗址第一阶段(2004-2008年)考古工作概述[M]//北京大学中国考古学研究中心, 浙江省文物考古研究所.田螺山遗址自然遗存综合研究.北京: 文物出版社, 2011: 7-39. Sun Guoping. Report on the 2004-2008 excavation at Tianluoshan[M]//Centre for the Study of Chinese Archaeology in Peking University, Zhejiang Province Institute of Archaeology and Cultural Heritage. Integrated Studies on the Natural Remains from Tianluoshan. Beijing: Cultural Relics Press, 2011: 7-39. |
| [20] |
吴小红, 秦岭, 孙国平.田螺山遗址的14C年代数据[M]//北京大学中国考古学研究中心, 浙江省文物考古研究所.田螺山遗址自然遗存综合研究.北京: 文物出版社, 2011: 40-46. Wu Xiaohong, Qin Ling, Sun Guoping. Radiocarbon date of Tianluoshan[M]//Centre for the Study of Chinese Archaeology in Peking University, Zhejiang Province Institute of Archaeology and Cultural Heritage. Integrated Studies on the Natural Remains from Tianluoshan. Beijing: Cultural Relics Press, 2011: 40-46. |
| [21] |
伊丽莎白·施密德著; 李天元译.动物骨骼图谱[M].北京: 中国地质大学出版社, 1992: 1-103. Schmid E; Translated by Li Tianyuan. Atlas of Animal Bones[M]. Beijing: China University of Geosciences Press, 1972: 1-103. |
| [22] |
西蒙·赫森著; 侯彦峰, 马萧林译.哺乳动物骨骼和牙齿鉴定方法指南[M].北京: 科学出版社, 2012: 1-116. Hillson S; Translated by Hou Yanfeng, Ma Xiaolin. Mammal Bones and Teeth: An Introductory Guide to Methods of Identification[M]. Beijing: Science Press, 2012: 1-116. |
| [23] |
O'Connor T. The Archaeology of Animal Bones[M]. Stroud: Sutton Publishing, 2000: 54-67.
|
| [24] |
Lyman R L. Quantitative Paleozoology[M]. Cambridge: Cambridge University Press, 2008: 21-82.
|
| [25] |
南川雅男, 松井章, 中村慎一, 等.由田螺山遗址出土的人类与动物骨骼胶质碳氮同位素组成推测河姆渡文化的食物资源与家畜利用[M]//北京大学中国考古学研究中心, 浙江省文物考古研究所.田螺山遗址自然遗存综合研究.北京: 文物出版社, 2011: 262-269. Minagawa M, Matsui A, Nakamura S, et al. Subsistence economy and animal husbandry: Stable isotope analysis on the human and animal remains from Tianluoshan.[M]//Centre for the Study of Chinese Archaeology in Peking University, Zhejiang Province Institute of Archaeology and Cultural Heritage. Integrated Studies on the Natural Remains from Tianluoshan. Beijing: Cultural Relics Press, 2011: 262-269. |
| [26] |
浙江动物志委员会. 浙江动物志淡水鱼类[M]. 杭州: 浙江科学技术出版社, 1991: 109-216. Editorial Committee of Faunal of Zhejiang. Fauna of Zhejiang: Freshwater Fishes[M]. Hangzhou: Zhejiang Science and Technology Publish, 1991: 109-216. |
| [27] |
湖北省水生生物研究所鱼类研究室. 长江鱼类[M]. 科学出版社, 1976: 44-213. Hubei Provincial Institute of Hydrobiology. The Fishes of the Yangtze River[M]. Beijing: Science Press, 1976: 44-213. |
| [28] |
铃木三男, 郑云飞, 能城修一, 等.浙江省田螺山遗址出土木材的树种鉴定[M]//北京大学中国考古学研究中心, 浙江省文物考古研究所.田螺山遗址自然遗存综合研究.北京: 文物出版社, 2011: 108-161. Suzuki M, Zheng Y, Noshiro S, et al. Identification of excavated wood materials from Tianluoshan, Zhejiang[M]//Centre for the Study of Chinese Archaeology in Peking University, Zhejiang Province Institute of Archaeology and Cultural Heritage. Integrated Studies on the Natural Remains from Tianluoshan. Beijing: Cultural Relics Press, 2011: 108-161. |
| [29] |
盛和林, 大泰司纪之, 陆厚基. 中国野生哺乳动物[M]. 北京: 中国林业出版社, 1999: 42-274. Sheng Helin, Ohtaishi Noriyuki, Lu Houji. The Mammalian of China[M]. Beijing: Chinese Forestry Press, 1999: 42-274. |
| [30] |
盛和林. 中国鹿类动物[M]. 上海: 华东师范大学出版社, 1992: 89-233. Sheng Helin. The Deer in China[M]. Shanghai: East China Normal University Press, 1992: 89-233. |
| [31] |
中国动物主题数据库[DB/OL]. http://www.zoology.csdb.cn/ China Animal Scientific Database[DB/OL]. http://www.zoology.csdb.cn/ |
| [32] |
付义强, 贾小东, 胡锦矗, 等. 江西桃红岭自然保护区夏季梅花鹿对生境的选择性[J]. 四川动物, 2006, 25(4): 185-187. Fu Yiqiang, Jia Xiaodong, Hu Jinchu, et al. Summer habitat selection by sika deer in Taohongling Nature Reserve, Jiangxi Province[J]. Sichuan Journal of Zoology, 2006, 25(4): 185-187. |
| [33] |
马继飞, 张恩迪, 章叔岩, 等. 清凉峰自然保护区梅花鹿秋季对栖息地利用的初步分析[J]. 动物学杂志, 2004, 39(5): 35-39. Ma Jifei, Zhang Endi, Zhang Shuyan, et al. Preliminary analysis on the habitat use by sika deer in Qingliangfeng National Natural Reserve in Autumn[J]. Chinese Journal of Zoology, 2004, 39(5): 35-39. DOI:10.3969/j.issn.0250-3263.2004.05.008 |
| [34] |
马春梅, 王冰翎, 刘泽雨, 等. 浙江田螺山遗址区孢粉学记录的中全新世植被、环境和人类活动初步研究[J]. 第四纪研究, 2018, 38(5): 1304-1312. Ma Chunmei, Wang Bingling, Liu Zeyu, et al. A preliminary study on vegetation, environment and human activity of pollen records during Middle-Late Holocene in Tianluoshan site, Ningshao Plain[J]. Quaternary Sciences, 2018, 38(5): 1304-1312. |
| [35] |
莫多闻, 孙国平, 史辰羲, 等.浙江田螺山遗址及河姆渡文化环境背景探讨[M]//北京大学中国考古学研究中心, 浙江省文物考古研究所.田螺山遗址自然遗存综合研究.北京: 文物出版社, 2011: 249-261. Mo Duowen, Sun Guoping, Shi Chenxi, et al. Environmental background of Tianluoshan and the Hemudu culture[M]//Centre for the Study of Chinese Archaeology in Peking University, Zhejiang Province Institute of Archaeology and Cultural Heritage. Integrated Studies on the Natural Remains from Tianluoshan. Beijing: Cultural Relics Press, 2011: 249-261. |
| [36] |
郑云飞, 陈旭高, 孙国平.田螺山遗址出土植物种子反映的食物生产活动[M]//北京大学中国考古学研究中心, 浙江省文物考古研究所.田螺山遗址自然遗存综合研究.北京: 文物出版社, 2011: 97-107. Zheng Yunfei, Chen Xugao, Sun Guoping. Food production at Tianluoshan: the analysis of macro-archaeobotanical remains[M]//Centre for the Study of Chinese Archaeology in Peking University, Zhejiang Province Institute of Archaeology and Cultural Heritage. Integrated Studies on the Natural Remains from Tianluoshan. Beijing: Cultural Relics Press, 2011: 97-107. |
| [37] |
傅稻镰, 秦岭, 赵志军, 等.田螺山遗址的植物考古分析[M]//北京大学中国考古学研究中心, 浙江省文物考古研究所.田螺山遗址自然遗存综合研究.北京: 文物出版社, 2011: 47-96. Fuller D Q, Qing Ling, Zhao Zhijun, et al. Archaeobotanical Analysis at Tianluoshan: Evidence for wild-food gathering, rice cultivation and the process of the evolution of morphologically domesticated rice[M]//Centre for the Study of Chinese Archaeology in Peking University, Zhejiang Province Institute of Archaeology and Cultural Heritage. Integrated Studies on the Natural Remains from Tianluoshan. Beijing: Cultural Relics Press, 2011: 47-96. |
| [38] |
秦岭, 傅稻镰, 张海. 早期农业聚落的野生食物资源域研究——以长江下游和中原地区为例[J]. 第四纪研究, 2010, 30(2): 245-261. Qin Ling, Fuller D Q, Zhang Hai. Modelling wild food resource catchments amongst early farmers: Case studies from the lower Yangtze and the Central China[J]. Quaternary Sciences, 2010, 30(2): 245-261. |
| [39] |
Zhang Ying. Exploring the wetland: Integrating the fish and plant remains into a case study from Tianluoshan, a middle Neolithic site in China[M]//Evangelia Pi NA08B kin, Arkadiusz Marciniak, Marta Bartkowiak. Environmental Archaeology. Cham: Springer, 2018: 199-227.
|
| [40] |
曹克清. 麋鹿研究[M]. 上海: 上海科技教育出版社, 2005: 26-77. Cao Keqing. Milu deer(Elaphurus davidianus)Research[M]. Shanghai: Shanghai Scientific Education Press, 2005: 26-77. |
| [41] |
van der Valk A. The Biology of Freshwater Wetlands[M]. Oxford: Oxford University Press, 2012: 13-86.
|
| [42] |
Keddy P A. Wetland Ecology: Principles and Conservation[M]. Cambridge: Cambridge University Press, 2010: 1-41.
|
| [43] |
付罗文, 袁靖. 重庆忠县中坝遗址动物遗存的研究[J]. 考古, 2006(1): 79-88. Flad Rowan, Yuan Jing. Research on the animal remains from Zhongba, Chongqing City[J]. Archaeology, 2006(1): 79-88. DOI:10.3969/j.issn.1001-6406.2006.01.007 |
| [44] |
莫林恒.高庙遗址出土鱼类遗存研究[D].长沙: 湖南大学硕士学位论文, 2011: 20-57. Mo Linheng. Preliminary Study on the Fish Remains Unearthed in Gaomiao Site[D]. Changsha: The Master's Dissertation of Hunan University, 2011: 20-57. |
| [45] |
卢山, 胡军华, 肖成云, 等. 洪湖鱼类物种组成及影响因素评价[J]. 野生动物学报, 2006, 27(6): 14-17. Lu Shan, Hu Junhua, Xiao Chengyun, et al. Fish species composition in Honghu Lake and estimation of influencing factors[J]. Chinese Wildlife, 2006, 27(6): 14-17. DOI:10.3969/j.issn.1000-0127.2006.06.005 |
| [46] |
Chen Weimin, Wu Qinglong. Fish compositions and fisheries[M]//Qin Boqiang. Lake Taihu, China: Dynamics and Environmental Change. Berlin: Springer Science & Business Media, 2008: 276-284.
|
| [47] |
Courtenay W R, Williams J D. Snakeheads(Pisces, Channidae): A Biological Synopsis and Risk Assessment[M]. Denver: US Geological Survey, 2004: 45-52.
|
| [48] |
浙江省文物考古研究所. 河姆渡: 新石器时代遗址考古发掘报告[M]. 北京: 文物出版社, 2003: 81-99. Zhejiang Provincial Institute of Archaeology and Cultural Heritage. Hemudu: Excavation Report of A Neolithic Site[M]. Beijing: Cultural Relics Press, 2003: 81-99. |
| [49] |
Colley S M. Fishing for facts. Can we reconstruct fishing methods from archaeological evidence?[J]. Australian Archaeology, 1987(24): 16-26. |
| [50] |
Gabriel O, Lange K, Da hm, et al. Von Brandt's Fish Catching Methods of the World(fourth edition)[M]. New Jersey: John Wiley & Sons, 2008: 12-375.
|
| [51] |
Matsui A. Salmon exploitation in Jomon archaeology from a wetlands point of view[J]. Journal of Wetland Archaeology, 2005, 5(1): 49-63. DOI:10.1179/jwa.2005.5.1.49 |
| [52] |
Nakajima T, Nakajima M, Mizuno T, et al. On the pharyngeal tooth remains of crucian and common carp from the Neolithic Tianluoshan site, Zhejiang Province, China, with remarks on the relationship between freshwater fishing and rice cultivation in the Neolithic Age[J]. International Journal of Osteoarchaeology, 2012, 22(3): 294-304. DOI:10.1002/oa.1206 |
| [53] |
蓝家森. 介绍几种捕捉乌鳢(乌鱼)的方法[J]. 生物学通报, 1958(12): 45. Lan Jiasen. Introduction of the methods of capturing snakeheads[J]. Bulletin of Biology, 1958(12): 45. |
| [54] |
Fuller D Q, Qin L, Zheng Y, et al. The domestication process and domestication rate in rice: Spikelet bases from the lower Yangtze[J]. Science, 2009, 323(5921): 1607-1610. DOI:10.1126/science.1166605 |
| [55] |
Fuller D Q, Qin L. Declining oaks, increasing artistry, and cultivating rice: The environmental and social context of the emergence of farming in the Lower Yangtze Region[J]. Environmental Archaeology, 2010, 15(2): 139-159. DOI:10.1179/146141010X12640787648531 |
| [56] |
Yunfei Z. Rice fields and modes of rice cultivation between 5000 and 2500 BC in East China[J]. Journal of Archaeological Science, 2009, 36(12): 2609-2616. DOI:10.1016/j.jas.2009.09.026 |
| [57] |
Nakajima T, Nakajima M, Yamazaki T. Evidence for fish cultivation during the Yayoi Period in western Japan[J]. International Journal of Osteoarchaeology, 2010, 20(2): 127-134. |
| [58] |
Gregory R, Guttman H. Capture and culture ricefield fisheries in Cambodia[M]//Nesbitt Harry J. Rice Production in Cambodia, Manila, International Rice Research Institute, 1997: 99-106.
|
Abstract
The Hemudu culture is one of the most important Neolithic cultures in the lower Yangtze River region, dating back to approximately 7000~6000 a B.P. The discussion of Hemudu subsistence has been focused on the origin and development of rice agriculture; meanwhile, studies on the exploitation of animal resources are scarce, and cannot meet the academic demands of current archaeology and zooarchaeology. Fortunately, rich and well-preserved animal and plant remains were unearthed from another Hemudu culture site, Tianluoshan(30°01'27″N, 121°22'46″E), which was first excavated in 2004, providing plenty materials for further investigation of Hemudu subsistence economy. Zooarchaeological research suggests a subsistence pattern predominated by hunting and fishing. This research proposed a brand new approach to interpret the subsistence economy from the spatial perspective, by integrating the Taxonomic Habitat Index analysis which was first employed by Andrews and Evans, and a common zooarchaeological quantitative method Minimum Number of Individuals. By studying the mammal and fish remains retrieved from the 2004 to 2010 excavation seasons, this research manages to evaluate the importance of each habitat to hunting and fishing activities. The results shows that hunting activities mostly occurred in the shrub and grassland habitat, and fishing was mainly practiced in wetland, which could be approached within 2~4 hours walking away from the Tianluoshan settlement, suggesting that these habitats were ideal for daily consumption. It also reveals that wetland is the source for stable food production, as the habitat for fish, water chestnuts, foxnut, and rice cultivation, and thus the key area for the entire subsistence economy. This research contributes zooarchaeological study in three ways:(1)a new method to interpret the subsistence economy, (2)detailed analysis of the Hemudu subsistence, and (3)an interdisciplinary attempt to integrate zooarchaeological and archaeobotanical research.