 2015, Vol.35
2015, Vol.35



树轮气候学(Dendroclimatology)以树木生理学为基础,以其生长特点为依据,研究树木生长与环境变化的关系,旨在获取气候代用资料,重建历史时期的气候变化规律[1]。树轮资料以其定年准确、 分辨率高、 分布范围广、 可与器测数据进行校准等优点而成为过去气候变化研究的重要手段之一[2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13]。树轮气候重建结果为揭示区域气候变化规律、 理解气候变化机制以及预估未来气候变化情景都提供了宝贵的基础资料。然而,目前大多数的树轮气候重建研究是基于树轮序列与器测记录之间的统计关系实现的,缺乏树轮生理机制方面的解释和验证。况且,树木生长与外界环境因子的关系比较复杂,经验的统计校准方法并不能用来研究树木生长究竟是如何被气候因子驱动的,也就不能量化不同气候指标对树木生长的贡献权重,使得在深入理解树木生长与环境变化关系方面受到局限[14]。以上统计方法的不足正是树轮生理模型的优势所在。
树轮生理模型从树木生长与气候因子响应的关系出发,同时考虑多个外界因子,用这些因子的变化模拟树木的生长过程。模型允许不同时期有不同的生长限制因子,避免了树木生长对气候响应的敏感性随时间变化等问题[15, 16, 17]。此外,模型研究还能够分析树木生长与外界环境的非线性关系,量化不同因子对树木生长的贡献权重,从更深层次揭示树木生长与环境因子间的复杂关系,为基于经验的统计关系提供生理依据,提高树轮气候重建的精度和可信度。
1 树轮生理模型的发展历程树轮生理模型是用数学方程来描述树木生长与外部环境之间的关系。模型的建立遵循树轮气候学研究中的均一性原理、 限制因子原理、 敏感性原理、 交叉定年以及复本原理[1]。建立模型的主要目的是模拟在气候变化驱动下树木生长特征的变化,分析不同环境因子与树木生长的非线性关系。20世纪60年代,Fritts[18]对树轮解剖特征的研究奠定了树轮-气候机理探索的基石。
1968年,Wilson和Howard[19]初步设计了树轮形成的基本情景,建立模型(简称Wilson-Howard模型)模拟了树木形成层区域细胞个数、 细胞大小、 细胞壁厚度的变化特征。模型的输入项由14个参数组成,各参数随时间而变化,因此必须对生长季内每天的参数值进行量测,有固定的数值之后才能应用该模型。另外,该模型无法模拟外部环境对形成层细胞生长的影响,也无法对其结果进行很好的校准和验证。随后,作者对其模型进行了改进[20, 21],但并没有克服其主要缺陷,只是对形成层细胞分化过程的不断优化。
1975年,Stevens[22]通过改进Wilson-Howard模型而建立了ARBOR模型。该模型基于22个输入参数对树木的生长和形成层区域进行了描述。然而该模型与Wilson-Howard的模型没有本质区别,只是将Wilson-Howard模型中以时间变量的数字形式转换为连续的曲线,同样没有涉及外部环境对树木生长的影响。
1990年,Vaganov[23]建立的TRACH模型,综合考虑了外界环境与形成层细胞生长之间的关系,但是模型缺乏土壤水分平衡、 树木光合作用以及蒸腾作用之间的内在关联。之后,Vaganov和Shashkin[24]联合建立了Vaganov-Shashkin模型(简称V-S模型),有效克服了以上缺陷。
1999年,Fritts等[25]融合了V-S和TRACH模型,对TRACH模型的细节部分进一步改进,提出了TREE-RING模型。此后,诸多基于树轮生理过程的模型应运而生[26, 27, 28, 29, 30, 31, 32],如Drew 等[33]在CABALA模型[34]的基础上建立了CAMBIUM模型模拟在每日尺度上,从髓芯到树皮的木材密度、 纤维、 管胞等各种解剖参数随外界环境因子的变化过程; Huang等[35]发现加拿大东部松柏类树木木质部的形成可以表述为形成层活动、 芽、 枝、 根的生长以及树种和采样点的函数。该研究揭示出,在树体内存在着最优机制来分配不同时间所积累的光合产物及储存的非结构性碳水化合物以用于不同器官或组织的生长。在此,树轮气候学家或树轮生态学家更为关注树干木质部的生长,即当年生长与上一年(多年)之间的联系,也就是树轮序列中的自相关问题[36]。其实,Le Roux等[37]较早曾提出此前树轮生理模型中需要改进的关键点: 碳在植物体内的存储、 分配及再转移问题。这些问题在MAIDEN模型[38]中得到了优化与改进。此外,该模型还包括了简化的水循环过程,如植物的蒸腾作用、 截流、 土壤蒸发、 排水及径流。在此基础上,MAIDENiso模型[27]的发展和应用则可为工业革命以来CO2浓度的增加是否促进了树木生长的争论[39, 40, 41, 42, 43, 44]提供有效的解决途径。
不同模型所涉及的生理过程和模拟内容不同,在实际应用中应根据需求而选取。Guiot等[45]列举了目前应用较普遍的模型所包含的不同生理过程: 水循环(water cycle)、 碳循环(carbon cycle)、 树体构成(tree compartments)、 木结构(wood structure)和同位素(isotopes)。其中,V-S模型没有包括同位素模块,MAIDENiso模型缺乏形成层结构模块,TREE-RING模型则相对全面。从树轮气候学和树轮生态学研究所需角度出发,本文将着重介绍V-S、 TREE-RING和MAIDENiso模型的机理、 运行环境和应用。
2 树轮生理模型V-S、 TREE-RING和MAIDENiso介绍(1)V-S模型
V-S模型[24]主要是针对形成层区域的研究,其机理是通过对影响细胞生长过程的外界主要气候因子的控制来模拟细胞的变化。该模型简化了树木生长的复杂过程,假定其他因子不构成限制树木生长的情况下,树木形成层的生长速率G(t)由光照GE(t)、 温度GT(t)和土壤湿度GW(t)各自影响的生长速率所共同决定,即:
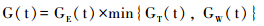
其中,光照对树木生长的影响GE(t)忽略了地球绕太阳运行时的离心率以及大气透射率的作用,可表示为纬度()、 太阳入射角(θ)和日长(hs)的函数。
温度对树木生长的影响GT(t)可表述为: 当积温达到给定温度(Tbeg)时,树木开始生长,遵循温度三基点原则(图 1): 当气温低于最低生长温度(Tmin)时,树木不生长; 当气温高于Tmin而低于最适生长温度下限(Topt1)时,树木生长速率随着气温的升高以线性速度增加; 当气温在Topt1和最适生长温度上限(Topt2)之间时(Topt2>Topt1),生长速度保持在最高水平,为一常数; 当气温高于Topt2低于最高生长温度(Tmax)时,随着气温升高树木生长呈线性下降。

|
图 1 V-S模型中树木生长对温度的响应曲线 Fig. 1 Typical curve of growth rate response to temperature used in the V-S model |
土壤湿度对树木生长的影响GW(t)类似于温度,也用分段函数来表示。包含了生长的最低土壤湿度(Wmin),生长的最适土壤湿度下限(Wopt1),生长的最适土壤湿度上限(Wopt2)和生长的最高土壤湿度(Wmax)4个参数,它们对应的含义同上,但其计算过程较温度更为复杂[46]。
V-S模型运行时需要输入的气象数据是日平均温度(T)和日降水(P)。还需输入模拟样点的纬度(phi),模拟起始年(syear)和结束年(eyear)。对其他参数的设置则需根据采样点的具体情况而进行调整。如果有足够的关于采样点的实验数据或多年观测数据,如三基点温度(Tmin、 Topt1、 Topt2和Tmax)、 开始生长的积温(Tbeg)、 植物根深(rootd)、 使土壤饱和的最大日降水量(Pmax)、 渗透到土壤的降水占总降水的比例系数(k1,与研究点的坡度、 树冠、 土壤特性等有关)等,这样就可以预先确定好各个参数,直接用模型进行模拟; 如果实验或观测数据不充分,则需根据对采样点的描述和部分实验数据,先对各个参数给定一个近似的经验值,然后用此近似值进行模拟,比较拟合效果,当效果比较好时,即模拟数据和实测数据变化形式比较吻合时,所选择的参数就作为最终的拟合参数对样点的其他数据进行拟合。如果各参数之间的比例关系调整不吻合,则可能导致模拟程序不运行。
模型输出项主要有模拟的轮宽值(trws)、 树木开始生长及结束生长日期、 树木生长速率G(t),包括由光照GE(t)、 温度GT(t)和土壤湿度GW(t)各自决定的相对生长速率等。一般而言,除模拟轮宽值与实测数据吻合或相关较好外,还需关注由树木开始生长及结束生长日期所决定的生长季的长短,需与实际样点观测到的生长季相匹配。在此基础上,分析由不同气候环境条件所决定的生长速率,界定影响研究区树木生长的主要外界因子。另外,可以统计生长季中由光照、 温度和土壤湿度分别决定的树木生长的天数,量化不同外界条件的贡献权重。根据模型,还可以预测理想状况下温度升高或降水减少对树木生长的影响。
近来,Tolwinski-Ward等[47]对V-S模型进一步简化,提出了V-S Lite 模型。该模型只取了V-S模型的生长模块,用来研究光照、 温度和土壤湿度对树木生长的影响。由于其舍弃了原来模型中对形成层区域描述的模块,因而并不能称之为完全的基于过程的模型。但通过调整模型中的参数值,即融合起始月份(integration start month,即Io)和融合终止月份(integration end month,即If),可以研究上年到当年不同月份气候环境变化对树木生长的影响。另外,该模型输入的是月尺度气候资料,降低了V-S模型对日尺度气候资料的要求,应用范围得到进一步扩展。
(2)TREE-RING模型
TREE-RING模型[25]是对V-S模型的优化,形成层细胞生长的计算采用改进的V-S模型的部分。通过把对树木光合速率的模拟与叶片的气孔活动、 光照、 温度、 土壤湿度和CO2浓度有效的联系起来,物理过程更加完整。模型除可以较好地模拟随外界气候变化的树木形成层活动、 光合速率、 呼吸速率、 同化速率以及树轮宽度、 树木根、 叶、 茎同位素的成分变化外,也可以用来诊断污染物对树轮结构的影响。
TREE-RING模型可划分为4个相关的模块: 1)微气候因子模块,气候资料被转化为直接影响树木生长机理过程的数值; 2)土壤水分平衡模块,通过对降水、 地表径流、 树木水分蒸散的描述来表达土壤水分平衡状况; 3)根、 茎、 叶的光合和呼吸模块,模拟了日光合作用,根、 茎、 叶中光合产物的分配,光合产物中一部分用于植物的生长,另外一部分用于保持各组织的呼吸作用; 4)形成层生长模块,模拟树根、 叶的生长,形成层细胞的分裂、 扩展和细胞壁增厚等过程,计算了形成层细胞的生长速率。 通过形成层细胞生长速率得出细胞个数、 细胞径向大小、 细胞壁厚度等解剖特征值,该模块是整个模型的核心部分。
模型以逐日最高(Tmax)和最低气温(Tmin)、 逐日降水量(P)和露点温度(Td)等为主要输入项,通过参数设置,充分考虑样点坡度、 林间郁闭度、 树木主根系的深度、 土层厚度及土壤最大含水量等因素的影响。但对土壤特性、 土壤中有机物、 树龄对树木生长等的影响未予考虑。此外,模型需要的同位素资料为大气CO2中δ13C及降水和水汽中 δ18O和δD值。TREE-RING模型中的参数主要包含在6个文件中,分别为Camb70.PAR、 iso.PAR、 trg70.PAR、 trg70.IO、 trg70.INF和Mcconst.CST。 其中Camb70.PAR是控制形成层生长的模块参数; iso.PAR是树木中碳、 氢、 氧的同位素参数; trg70.PAR是与局地树木生长条件有关的模型参数; trg70.IO用于设置模型初值参数,该参数设定的准确与否将直接影响最终的模拟效果; trg70.INF用来控制模型界面参数; Mcconst.CST是大气中的物理常数,如太阳常数、 大气密度、 标准大气压等参数。模型中与局地条件关系比较紧密的参数trg70.PAR是模拟调试的重点部分。
TREE-RING模型可以用来研究CO2浓度变化对树木生长的影响。即通过开展CO2浓度变化的敏感性试验,如实验一可假定CO2为目前实测的浓度值,实验二假定CO2是工业革命以前的水平。根据两组试验观测分析对树轮宽度及其他参数的模拟结果,探讨CO2浓度变化对树木生长的影响。
(3)MAIDENiso模型
MAIDEN模型[38]设计的主要目的是分析树木生长与外界环境的关系,模拟的树干径向增长量尤其适用于树轮气候学或树轮生态学研究。将同位素模块植入后,就形成了MAIDENiso模型[27]。MAIDENiso属于完全的基于过程的树木生理模型,因为该模型的输入项(气候因子)与输出项(树轮参数,如树轮宽度、 树轮碳氧同位素)之间的关系并没有依据传统经验预先给出特定的参数值,而只是作为数学方程式在模型中被明确地描述出来。模型通过使用气候数据来模拟外界环境对树木光合作用和呼吸作用的影响,并以此来确定固碳量和树干径向增长量。该树干径向增长量可以看作类似于树轮宽度数据进而与实测资料进行对比分析。
与V-S和TREE-RING模型相比,MAIDENiso模型的功能更为强大。首先,与TREE-RING模型类似,MAIDENiso模型也可以用来分析CO2浓度变化对树木生长的影响; 其次,模型能够模拟研究区的物候变化过程,捕捉与蒸腾作用、 气孔导度、 呼吸作用和光合作用密切相关的气候因子的影响; 第三,由于该模型整合了碳氧同位素在树体内的形成过程,不仅可以用来研究与光合作用密切相关的木材产量的变化,而且还能够追踪森林生态系统中的水循环过程,尤其便于了解大气中的水汽通过土壤再到树体内的整个转换过程。
模型需要输入的参数主要有: 树轮宽度指数、 树轮δ13C和 δ18O值、 大气CO2浓度值、 大气δ13C及降水 δ18O同位素值等。其输出的结果则有模拟的温度、 降水、 树轮宽度指数等。树木生长的生理过程及模型参数的调试与V-S和TREE-RING模型类似,在此不予复述。
总体而言,V-S、 TREE-RING和MAIDENiso模型在一定程度上可以说是一个逐步完善的过程。但是,其各自的优势又是不能被相互取代的。如V-S模型的物理过程简单,需要输入的参数特征值在一般的树轮研究样点都可以得到满足,适用范围广泛。而TREE-RING和MAIDENiso模型完善的过程所需要的更多监测数据与其有限的研究样点可能相伴而行。在实际应用时应根据不同的需求而合理选择。
3 树轮生理模型V-S、 TREE-RING和MAIDENiso的应用作为客观且精细的描述工具,树轮模型已被广泛用于分析树木生长与环境变化关系的研究中。另外,树轮模型对于过去气候的研究也具有相当大的潜力,有助于研究者深入理解树轮—气候档案形成的过程,提高基于经验统计方法而进行气候重建的可靠性。因此,自树轮生理模型建立之日起就得到了比较广泛的关注和应用。
(1)V-S模型的应用
V-S模型目前主要用来模拟树轮宽度的变化过程,区分光照、 温度和土壤湿度对树木生长的影响,界定限制树木生长的主要外界因子。如Evans等[48]成功模拟了北美和俄罗斯境内的198条树轮序列。通过V-S模型分析得到的美国东南部树轮序列与气候因子间的相关关系[49]对该区树轮气候重建研究具有重要的指示意义。除了其起源地北美和俄罗斯外,在突尼斯地区[50]该模型也得到了较好应用。
此外,V-S模型也被应用到了我国树轮研究的部分地区。Zhang等[51]使用该模型模拟了柴达木盆地东缘祁连圆柏(Sabina przewalskii)生长与气候因子的关系,发现在该干旱区,祁连圆柏的生长主要受当地水分条件的控制。在贺兰山地区,Shi等[52, 53, 54]模拟了该区油松(Pinus tabulaeformis)生长的气候响应过程,发现4月到5月中旬树木的生长主要受温度条件的控制,从5月下旬到8月底,降水则成为主要限制因子。模拟结果为统计的树轮-气候关系提供了机理解释。
在此,本文也尝试了利用V-S模型模拟青藏高原东北部德令哈样点祁连圆柏的生长过程(轮宽原始数据来自Yang等[4]),经过数次实验发现,本样点模拟过程受模型中Wmin、 Wopt1、 Tmin和Topt1参数变化的显著影响。经过调试,模拟得到的树轮宽度序列与实测序列(图 2)在1957~2011年的相关系数达0.64(p<0.001)。一阶差和5年滑动后两者的相关系数分别为0.55和0.69。

|
图 2 模拟的德令哈树轮宽度序列(虚线图) 与实测序列(实线图)在1957~2011年的对比 Fig. 2 Comparison between modelled(dot-dashed line) and actual(straight line)tree-ring width series over the common period of 1957~2011 at the Delingha site |
根据模拟结果,该地区的温度条件在4月中旬后便可满足树木生长的需求(图 3),但是土壤湿度到5月下旬才能达到生长的最低需求,树木在此时开始生长。在树木生长的主要时期(5月下旬到8月中旬),土壤湿度条件是其主要限制因子。8月下旬到生长季结束,土壤湿度条件完全满足了树木生长的需求,温度条件限制树木的生长。整个生长季,由光照条件决定的生长速率尚不能构成限制树木生长的主要条件。模拟结果所显示的土壤湿度条件是当地树木生长的主要限制因子验证了此前基于统计相关而进行降水重建[4, 55]的可靠性。

|
图 3 模拟得到的由不同外界环境因子所决定的 德令哈地区树木平均(1957~2011年)相对生长速率 Fig. 3 Mean integrated relative growth rate(G,black line)and partial growth rates depending on solar radiation(GE,violet line),temperature(GT,red line)and soil moisture(GW,blue line)during the period of 1957~2011 for Delingha region |
总之,V-S模型凭借其自身的优势得到了较为广泛的应用,已从其起源地扩展到了其他区域和其他树种。模型的优势[56]主要在于简化了树木生长的复杂过程。在生长季的一定时间段内,树木生长的速率不能超过其重要限制因子所允许的范围。另外,模型需要输入的数据在一般的树轮研究样点就可得到满足。然而,V-S模型没有包括同位素模块,不能用来研究CO2浓度变化对树木生长的影响。
(2)TREE-RING模型的应用
TREE-RING模型[25]将外界环境控制树木形成层径向增长的过程与涉及光合作用、 呼吸作用等过程的碳平衡有机联系在一起,被认为是目前最为完整的生理模型[56]。但模型构建后,目前对其应用的相关报道比较有限。
在中国,李雁等[57]对柴达木盆地东缘青海云杉(Picea crassifolia)的生长过程进行了模拟,表明限制该区树木生长的主要气候因子为土壤湿度状况,尤其是生长季前期的土壤湿度状况,与用V-S模型在该区对祁连圆柏模拟的结果相一致[51]; 尹红等[58]对小兴安岭红松(Pinus koraiensis)的生长过程进行了模拟; 此外,尹红[59]通过简单的敏感性实验还发现,CO2浓度的增加对红松生长有助长作用,即存在施肥效应; 进一步的预测结果[60]表明:CO2浓度的不断增加伴随温度的持续上升会导致红松树木径向生长量的不断增加。上述结果对未来树木生长的适应研究及制定相应的森林生态系统管理措施都提供了基本参考依据。
TREE-RING模型较V-S模型设置了更多的参数特征值来对树木的生长过程进行精细描述。但是,模型对较多监测或实验观测数据的输入要求(如日降水中 δ18O的值,目前在全球的绝大多数区域都是缺失的)有可能是其没有得到广泛应用的主要原因。
(3)MAIDENiso模型的应用
树木生长如何响应工业革命以来CO2浓度的增加引起了广泛关注[40, 61, 62]。如研究发现[63]最近几十年澳大利亚西部白柏树(Callitris columellaris)的径向增长并没有对CO2浓度的增加做出响应; Gedalof和Berg[64]的分析说明CO2的施肥效应仅影响了全球范围20%的树轮研究样点; 类似其他从树轮序列中检测各种信号并进行归因的研究也反对CO2的施肥效应[40, 65, 66]。但是,也有实验揭示近几十年来CO2的施肥效应显著,改善了树木的生长状况,提高了水分利用效率和森林生产力[41, 44, 67]。从树轮气候学研究的角度出发,这些争论必须得到解决。倘若树轮宽度序列确实叠加了工业革命以来CO2浓度增加的印记,那么进行气候重建所用的转换函数方法是值得怀疑的。因为不能假定器测记录时段以来的树轮序列与气候因子关系在工业革命以前CO2浓度较低的情况下依然是存在的。
解决上述争端,目前有两种方法[68]。一是采用数据同化技术[69, 70],但是该方法并不太适用于树轮资料,因为树轮序列中的气候信号与噪音还不能被明确地分离出来;另外一种方法即使用反演模型,用具有一定生理意义的模型反演过去的气候变化。Tolwinski-Ward等[47]所提出的V-S Lite模型堪称这一方法的先驱。但是,该模型的算法比较简单,许多重要的生理过程,如气孔导度、 光合作用、 呼吸作用等均没有包括在内。另外,该模型仅仅使用了树轮序列中的单一指标(轮宽数据),而不同指标(碳氧同位素)的综合对比研究才能更有效地提取树轮资料中的气候信息[71]。 包括了碳循环和同位素模块的树轮生理模型MAIDENiso[27]的构建则能够较好地弥补V-S Lite模型的缺陷。
Boucher等[68]将两种不同浓度的CO2序列分别输入MAIDENiso模型,模拟得到的夏季温度序列相关达0.75,说明在高频变化上两者区别不显著。但是,在低频趋势上,固定CO2浓度值280ppm所重建的夏季温度序列(图 4,红线)存在一个下降的趋势,与观测到的上升趋势不吻合。只有将实际的CO2浓度值输入反演模型中,才能更合理地与观测到的夏季温度序列在高低频变化上均相匹配(图 4,黑线)。该研究揭示出CO2浓度增加对研究区树木的生长具有肥化作用。

|
图 4 使用MAIDENiso模型重建的不同CO2浓度情景下法国Fontainebleau林区夏季温度(a)和降水(b)序列[68] 黑线及相应的95%置信区间是实测到的CO2浓度输入模型中得到的重建值,红线及相应95%置信区间是工业革命以前固定的 CO2浓度值(280ppm)输入模型后得到的重建结果,绿虚线是实测的1953~1999年的温度和降水值,相应的直线代表长期趋势 Fig. 4 MAIDENiso model results of the reconstructed summer temperature(a) and precipitation(b) at two different CO2 concentration scenarios for the Fontainebleau tree-ring site,France[68]. The black curves are the results from realistic CO2 data,the red curves are from fixed CO2 data(280ppm),and the green curves are the instrumental data from the year 1953 to 1999. The shaded areas are the associated 95% confidence level. The corresponding linear lines indicate the multidecadal trends in reconstructed and observed climate series |
根据该研究不难看出,MAIDENiso模型的反演算法较传统的转换函数方法更有优势,其中最关键的即是其能够单独分离出CO2浓度增加和气候变化对树木生长的贡献权重。用该反演方法进行树轮气候重建不仅能够减少人为温室气体排放所导致的信息偏差,同时还兼备了完善的树轮生理基础。该模型的应用需要具备大量的监测数据的支持[27, 68],这可能在一定程度上限制了其在大空间范围的应用。但是,凭借其自身的优势,在未来的研究中应该会有广阔的应用平台。
4 结语树轮生理模型的建立、 优化和应用在国外均取得了蓬勃的发展,突出表现为已有模型的多样化、 研究内容的丰富化和研究方法的灵活化。已有模型在分析全球变化与森林生产力的关系[72]、 研究森林生态系统中的水循环[73]、 界定CO2浓度变化对树木生长的影响[14]以及剖析木材结构变化[33]等方面都发挥了其独特的优势。诚然,这些方法的应用并不意味着传统的经验统计方法应该舍弃,而只是表明随着认识的深入和技术的进步,人们分析这些自然代用指标的手段得到了不断扩充。正是由于传统树轮气候学或树轮生态学的成熟与发展,人们才积累了足够的资料来建立各种模型,从机理上剖析树木生长与外界各种环境因子的关系,以不断提高树轮气候重建的精度和可信度。
以模型较高的真实性、 精准性和通用性为标准[74],未来研究一方面应更加关注CO2浓度变化对树木生理生态过程,如气孔导度、 呼吸作用、 光合作用、 水分利用效率、 养分的吸收和利用等的长期影响,以不断提高人们对其复杂交互过程的认识水平,改进模型中的计算模块; 另一方面,应提高所建树轮序列的质量,改进从树轮序列中提取气候信息的技术,方能更好地与模拟结果进行对比。此外,目前绝大多数模型主要是用于研究自然状态下气候变化对森林生态系统的影响,当需要考虑人为管理策略或非气候因子,如树木的死亡和与之相关的立地条件的改变[75],虫害[76]或火灾[77, 78]等的干扰时,模型的应用就会受到一定程度的限制。还有,土壤矿物成分及树龄对树木生长的影响一般也没有被考虑到,以上不足也将是未来树轮模型研究发展的方向。
具体到我国,树轮模拟研究起步较晚,并且都是利用国外建立好的模型,因此首先对模型的异地应用提出了挑战,再加上我国也缺乏长期实测数据的支持而导致模型参数的调试工作具有较大困难,目前研究内容比较单调,所取成果相对有限。然而,考虑到树轮模型研究方法具有其他方法(如经验统计方法)所不可替代的优势,我国目前的树轮研究样点也已分布广泛,从事树轮研究工作的机构和人数逐渐增加,因此,期待更多的研究者参与其中,提升我国树轮生理模型研究的整体水平。
致谢 感谢俄罗斯西伯利亚联邦大学Vladimir V. Shishov教授对V-S模型的运行环境、 参数调试等工作的悉心指导; 另外,感谢Etienne Boucher教授提供本文中的图 4。衷心感谢审稿专家和编辑部老师对本文提出的修改意见。
| 1 | Fritts H C. Tree Rings and Climate. New York:Academic Press, 1976. 1~576 |
| 2 | Hughes M K, Swetnam T W, Diaz H F. Dendroclimatology:Progress and Prospect. New York:Springer Science+Business Media B. V., 2011. 1~365 |
| 3 | Büntgen U, Trnka M, Krusic P J et al. Tree-ring amplification of the early-19th century summer cooling in central Europe. Journal of Climate, 2015,doi:10.1175/JCLI-D-14~00673.1 |
| 4 | Yang B, Qin C, Wang J L et al. A 3500-year tree-ring record of annual precipitation on the northeastern Tibetan Plateau.Proceedings of the National Academy of Sciences of the United States of America, 2014,111 (8):2903~2908 |
| 5 | 杨 保. 树轮记录的小冰期以来青藏高原气候变化的时空特征. 第四纪研究, 2012,32 (1):81~94 Yang Bao. Spatisl and temporal patterns of climate variations over the Tibetan Plateau during the period 1300~2010. Quaternary Sciences, 2012,32 (1):81~94 |
| 6 | 王亚军, 马玉贞, 鲁瑞洁等. 树木年轮记录的祁连山东段公元1895年来的气温变化. 第四纪研究, 2009,29 (5):905~912 Wang Yajun, Ma Yuzhen, Lu Ruijie et al. Reconstruction of mean temperatures of January to August since A.D. 1895 based on tree-ring data in the eastern part of Qilian Mountains. Quaternary Sciences, 2009,29 (5):905~912 |
| 7 | 李宗善, 刘国华, 伍 星等. 川西米亚罗地区岷江冷杉林过去223年森林净初级生产力重建. 第四纪研究, 2014,34 (4):830~847 Li Zongshan, Liu Guohua, Wu Xing et al. Tree ring-based net primary production of Abies faxoniana forest over the past 223 years in Miyaluo of western Sichuan, China. Quaternary Sciences, 2014,34 (4):830~847 |
| 8 | 刘昶智, 勾晓华, 方克艳等. 甘肃南部公元1824年以来降水重建. 第四纪研究, 2013,33 (3):518~525 Liu Changzhi, Gou Xiaohua, Fang Keyan et al. Precipitation reconstruction in southern Gansu Province since A.D. 1824. Quaternary Sciences, 2013,33 (3):518~525 |
| 9 | 勾晓华, 杨梅学, 彭剑峰等. 树轮记录的阿尼玛卿山区过去830年夏半年最高温变化. 第四纪研究, 2006,26 (6):991~998 Gou Xiaohua, Yang Meixue, Peng Jianfeng et al. Maximum temperature reconstruction for Anmaqing Mountains over past 830 years based on tree-ring records. Quaternary Sciences, 2006,26 (6):991~998 |
| 10 | 邵雪梅, 王树芝, 岩 徐等. 柴达木盆地东北部3500年树轮定年年表的初步建立. 第四纪研究, 2007,27 (4):477~485 Shao Xuemei, Wang Shuzhi, Xu Yan et al. A 3500-year master tree-ring dating chronology from the northeastern part of the Qaidam Basin. Quaternary Sciences, 2007,27 (4):477~485 |
| 11 | 刘晶晶, 杨 保. 西藏浪卡子地区树轮宽度特征值年表的建立. 第四纪研究, 2014,34 (6):1280~1287 Liu Jingjing, Yang Bao. Development of a tree-ring-width chronology using the Eigen analysis method in Langkazi regions of Tibet. Quaternary Sciences, 2014,34 (6):1280~1287 |
| 12 | 任军莉, 刘 禹, 宋慧明等. 甘肃临夏地区过去195年最高温度历史重建——基于紫果云杉树轮宽度资料. 第四纪研究, 2014,34 (6):1270~1279 Ren Junli, Liu Yu, Song Huiming et al. The historical reconstruction of the maximum temperature over the past 195 years, Linxia region, Gansu Province——Based on the data from Picea purpureaMast. Quaternary Sciences, 2014,34 (6):1270~1279 |
| 13 | 蔡秋芳, 刘 禹. 山西五鹿山油松树轮宽度年表的建立及过去百余年5~6月平均气温变化. 第四纪研究, 2013,33 (3):511~517 Cai Qiufang, Liu Yu. The development of a tree-ring width chronology and the May-June mean temperature variability in Wulu Mountain, north-Central China. Quaternary Sciences, 2013,33 (3):511~517 |
| 14 | Breitenmoser P, Brönnimann S, Frank D. Forward modelling of tree-ring width and comparison with a global network of tree-ring chronologies. Climate of the Past, 2014,10 (2):437~449 |
| 15 | Wilmking M, Juday G P, Barber V A et al. Recent climate warming forces contrasting growth responses of white spruce at treeline in Alaska through temperature thresholds. Global Change of Biology, 2004,10 (10):1724~1736 |
| 16 | D'Arrigo R, Wilson R, Liepert B et al. On the 'Divergence Problem’ in Northern Forests:A review of the tree-ring evidence and possible causes. Global and Planetary Change, 2008,60 (3~4):289~305 |
| 17 | Briffa K R, Schweingruber F H, Jones P D et al. Reduced sensitivity of recent tree-growth to temperature at high northern latitudes. Nature, 1998,391 (6668):678~682 |
| 18 | Fritts H C. Growth-rings of trees:Their correlation with climate. Science, 1966,154 (3752):973~979 |
| 19 | Wilson B F, Howard R A. A computer model for cambial activity. Forest Science, 1968,14 (1):77~90 |
| 20 | Wilson B F. A diffusion model for tracheid production and enlargement in conifers. Botanical Gazette, 1973,134 (3):189~196 |
| 21 | Howard R, Wilson B. A stochastic model for cambial activity. Botanical Gazette, 1972,133 (4):410~414 |
| 22 | Stevens D W. A computer program for simulating cambial activity and ring growth. Tree-Ring Bulletin, 1975, 35 :49~56 |
| 23 | Vaganov E A. Methods of Dendrochronology:Applications in the Environmental Sciences. Netherlands:Kluwer Academic Publishers, 1990.63~75 |
| 24 | Shashkin A V, Vaganov E A. Simulation model of climatically determined variability of conifers' annual increment(on the example of Scots pine in the steppe zone). Russia Journal of Ecology, 1993, 24 :275~280 |
| 25 | Fritts H C, Shashkin A, Downes G M et al. Tree-ring Analysis:Biological, Methodological and Environmental Aspects. London: CABI Publishing, 1999.3~32 |
| 26 | Davi H, Dufrêne E, Granier A et al. Modelling carbon and water cycles in a beech forest. Ecological Modelling, 2005,185 (2~4):387~405 |
| 27 | Danis P A, Hatté C, Misson L et al. MAIDENiso:A multiproxy biophysical model of tree-ring width and oxygen and carbon isotopes. Canadian Journal of Forest Research, 2012,42 (9):1697~1713 |
| 28 | Dufrêne E, Davi H, Franois C et al. Modelling carbon and water cycles in a beech forest. Ecological Modelling, 2005,185 (2~4):407~436 |
| 29 | Eglin T, Franois C, Michelot A et al. Linking intra-seasonal variations in climate and tree-ring δ 13 C:A functional modelling approach. Ecological Modelling, 2010,221 (15):1779~1797 |
| 30 | Ogee J, Barbour M M, Wingate L et al. A single-substrate model to interpret intra-annual stable isotope signals in tree-ring cellulose. Plant, Cell and Environment, 2009,32 (8):1071~1090 |
| 31 | Deckmyn G, Evans S P, Randle T J. Refined pipe theory for mechanistic modeling of wood development. Tree Physiology, 2006, 26 :703~717 |
| 32 | Kramer E M. A mathematical model of pattern formation in the vascular cambium of trees. Journal of Theoretical Biology, 2002,216 (2):147~158 |
| 33 | Drew D M, Downes G M, Battaglia M. CAMBIUM, a process-based model of daily xylem development in Eucalyptus. Journal of Theoretical Biology, 2010,264 (2):395~406 |
| 34 | Battaglia M, Sands P, White D et al. CABALA:A linked carbon, water and nitrogen model of forest growth for silvicultural decision support. Forest Ecology and Management, 2004,193 (1~2):251~282 |
| 35 | Huang J G, Deslauriers A, Rossi S. Xylem formation can be modeled statistically as a function of primary growth and cambium activity. New Phytologist, 2014,203 (3):831~841 |
| 36 | Berninger F, Hari P, Nikinmaa E et al. Use of modeled photosynthesis and decomposition to describe tree growth at the northern tree line. Tree Physiology, 2004, 24 :193~204 |
| 37 | Le Roux X, Lacointe A, Escobar-Gutiérrez A et al. Carbon-based models of individual tree growth:A critical appraisal. Annals of Forest Science, 2001, 58 :469~506 |
| 38 | Misson L. MAIDEN:A model for analyzing ecosystem processes in dendroecology. Canadian Journal of Forest Research, 2004,34 (4):874~887 |
| 39 | Andreu-Hayles L, Planells O, GutiÉRrez E et al. Long tree-ring chronologies reveal 20th century increases in water-use efficiency but no enhancement of tree growth at five Iberian pine forests. Global Change of Biology, 2011,17 (6):2095~2112 |
| 40 | Girardin M P, Bernier P Y, Raulier F et al. Testing for a CO2 fertilization effect on growth of Canadian boreal forests. Journal of Geophysical Research, 2011,116 (G1):G01012 |
| 41 | Keenan T F, Hollinger D Y, Bohrer G et al. Increase in forest water-use efficiency as atmospheric carbon dioxide concentrations rise. Nature, 2013,499 (7458):324~327 |
| 42 | Levesque M, Siegwolf R, Saurer M et al. Increased water-use efficiency does not lead to enhanced tree growth under xeric and mesic conditions. New Phytologist, 2014,203 (1):94~109 |
| 43 | Peñuelas J, Canadell J G, Ogaya R. Increased water-use efficiency during the 20th century did not translate into enhanced tree growth. Global Ecology and Biogeography, 2011,20 (4):597~608 |
| 44 | Norby R J, Delucia E H, Gielen B et al. Forest response to elevated CO2 is conserved across a broad range of productivity.Proceedings of the National Academy of Sciences of the United States of America, 2005,102 (50):18052~18056 |
| 45 | Guiot J, Boucher E, Gea-Izquierdo G. Process models and model-data fusion in dendroecology. Frontiers in Ecology and Evolution, 2014, 2 :1~12 |
| 46 | Vaganov E A, Hughes M K, Shashkin A V. Growth Dynamics of Conifer Tree Rings:Images of Past and Future Environments. New York:Springer Science & Business Media, 2006.189~243 |
| 47 | Tolwinski-Ward S E, Evans M N, Hughes M K et al. An efficient forward model of the climate controls on interannual variation in tree-ring width. Climate Dynamics, 2011,36 (11~12):2419~2439 |
| 48 | Evans M N, Reichert B K, Kaplan A et al. A forward modeling approach to paleoclimatic interpretation of tree-ring data. Journal of Geophysical Research, 2006,111 (G3):G03008 |
| 49 | Anchukaitis K J, Evans M N, Kaplan A et al. Forward modeling of regional scale tree-ring patterns in the Southeastern United States and the recent influence of summer drought. Geophysical Research Letters, 2006,33 (4):L04705 |
| 50 | Touchan R, Shishov V V, Meko D M et al. Prosess based model sheds light on climate sensitivity of Mediterranean tree-ring width. Biogeosciences, 2012,9 (3):965~972 |
| 51 | Zhang Yongxiang, Shao Xuemei, Xu Yan et al. Process-based modeling analyses of Sabina przewalskiigrowth response to climate factors around the northeastern Qaidam Basin. Chinese Science Bulletin, 2011,56 (4):1518~1525 |
| 52 | Shi J, Liu Y, Vaganov E A et al. Statistical and process-based modeling analyses of tree growth response to climate in semi-arid area of north Central China: A case study of Pinus tabulaeformis. Journal of Geophysical Research, 2008,113 (G1):G01026 |
| 53 | 史江峰, 刘 禹, Vaganov E 等. 贺兰山油松生长的气候响应机制初步探讨. 第四纪研究, 2005,25 (2):245~251 Shi Jiangfeng, Liu Yu, Vaganov E et al. A primary discussion on the climate response of Pinus tabulaeformis in the Helan Mountain. Quaternary Sciences, 2005,25 (2):245~251 |
| 54 | 史江峰, 刘 禹, 蔡秋芳等. 油松(Pinus tabulaeformis)树轮宽度与气候因子统计相关的生理机制——以贺兰山地区为例. 生态学报, 2006,26 (3):697~705 Shi Jiangfeng, Liu Yu, Cai Qiufang et al. A case study of physiological characteristics of statistical correlation between Pinus tabulaeformis tree-ring width and climate factors. Acta Ecologica Sinica, 2006,26 (3):697~705 |
| 55 | Sheppard P R, Tarasov P E, Graumlich L J et al. Annual precipitation since 515 BC reconstructed from living and fossil juniper growth of northeastern Qinghai Province, China. Climate Dynamics, 2004,23 (7~8):869~881 |
| 56 | Vaganov E A, Anchukaitis K J, Evans M N. How well understood are the processes that create dendroclimatic records?A mechanistic model of the climatic control on conifer tree-ring growth dynamics. In:Hughes M K, Swetnam T W, Diaz H F eds. Dendroclimatology, Developments in Paleoenvironmental Research.New York:Springer Science+Business Media, 2011.37~75 |
| 57 | 李 雁, 邵雪梅, 国 咖. 柴达木盆地东缘地区青海云杉生长的气候响应机制初步探讨. 高原气象, 2010,29 (2):349~358 Li Yan, Shao Xuemei, Guo Ka. Primary discussion of the climate mechanism response of Picea crassifolia on east edge of Qaidam Basin. Plateau Meteorology, 2010,29 (2):349~358 |
| 58 | 尹 红, 刘洪滨, 郭品文等. 小兴安岭低山区红松生长的气候响应机制. 生态学报, 2009,29 (12):6333~6341 Yin Hong, Liu Hongbin, Guo Pinwen et al. An analysis on the climate response mechanism of the growth of Pinus koraiensis in the lower mountains of Xiaoxing'anling. Acta Ecologica Sinica, 2009,29 (12):6333~6341 |
| 59 | 尹 红. 基于TREE-RING模型的小兴安岭红松树轮生长气候响应研究. 南京: 南京信息工程大学博士毕业论文, 2009. 1~141 Yin Hong. The Study on the Climatic Response of the Growth of Pinus koraiensisin the Xiaoxing'anling Based on the TREE-RING Eco-physiological Model.Nanjing:The Ph.D Thesis of Nanjing University of Information Science and Technology, 2009. 1~141 |
| 60 | 尹 红, 王 靖, 刘洪滨等. 小兴安岭红松径向生长对未来气候变化的响应. 生态学报, 2011,,31 (24):7343~7350 Yin Hong, Wang Jing, Liu Hongbin et al. A research on the response of the radial growth of Pinus koraiensisto future climate change in the Xiaoxing'anling. Acta Ecologica Sinica, 2011,,31 (24):7343~7350 |
| 61 | Handa I T, Körner C, Hättenschwiler S. A test of the treeline carbon limitation hypothesis by in situ CO2 enrichment and defoliation. Ecology, 2005,86 (5):1288~1300 |
| 62 | van der Sleen P, Groenendijk P, Vlam M et al. No growth stimulation of tropical trees by 150 years of CO2 fertilization but water-use efficiency increased. Nature Geoscience, 2015,8 (1):24~28 |
| 63 | Li G, Harrison S P, Prentice I C. A model analysis of climate and CO2 controls on tree growth in a semi-arid woodland. Biogeosciences Discussions, 2015,12 (6):4769~4800 |
| 64 | Gedalof Z, Berg A A. Tree ring evidence for limited direct CO2 fertilization of forests over the 20th century. Global Biogeochemical Cycles, 2010,24 (3):GB3027 |
| 65 | Silva L C R, Anand M, Leithead M D. Recent widespread tree growth decline despite increasing atmospheric CO2. PloS One, 2010,5 (7):e11543 |
| 66 | Silva L C, Horwath W R. Explaining global increases in water use efficiency:Why have we overestimated responses to rising atmospheric CO2 in natural forest ecosystems?PloS One, 2013,8 (1):e53089 |
| 67 | Norby R J, Wullschleger S D, Gunderson C A et al. Tree responses to rising CO2 in field experiments:Implications for the future forest. Plant, Cell and Environment, 1999,22 (6):683~714 |
| 68 | Boucher É, Guiot J, Hatté C et al. An inverse modeling approach for tree-ring-based climate reconstructions under changing atmospheric CO2 concentrations. Biogeosciences, 2014,11 (12):3245~3258 |
| 69 | Garreta V, Miller P A, Guiot J et al. A method for climate and vegetation reconstruction through the inversion of a dynamic vegetation model. Climate Dynamics, 2010,35 (2~3):371~389 |
| 70 | Goosse H, Crespin E, de Montety A et al. Reconstructing surface temperature changes over the past 600 years using climate model simulations with data assimilation. Journal of Geophysical Research, 2010,115 (D9):D09108 |
| 71 | McCarroll D, Loader N J. Stable isotopes in tree rings. Quaternary Science Reviews, 2004,23 (7~8):771~801 |
| 72 | Rathgeber C, Nicault A, Kaplan J O et al. Using a biogeochemistry model in simulating forests productivity responses to climatic change and CO2 increase:Example of Pinus halepensis in Provence(south-East France). Ecological Modelling, 2003,166 (3):239~255 |
| 73 | Berninger F, Nikinmaa E. Implications of varying pipe model relationships on Scots pine growth in different climates. Functional Ecology, 1997,11 (2):146~156 |
| 74 | Hanninen H. Effects of climatic change on trees from cool and temperate regions:An ecophysiological approach to modelling of bud burst phenology. Canadian Journal of Botany, 1995,73 (2):183~199 |
| 75 | Bigler C, Bugmann H. Predicting the time of tree death using dendrochronological data. Ecological Applications, 2004,14 (3):902~914 |
| 76 | Hogg E H, Brandt J P, Kochtubajda B. Growth and dieback of aspen forests in northwestern Alberta, Canada, in relation to climate and insects. Canadian Journal of Forest Research, 2002,32 (5):823~832 |
| 77 | Covington W, Fule P, Hart S et al. Modeling ecological restoration effects on ponderosa, pine forest structure. Restoration Ecology, 2001,9 (4):421~431 |
| 78 | Hogg E H, Wein R W. Impacts of drought on forest growth and regeneration following fire in southwestern Yukon, Canada. Canadian Journal of Forest Research, 2005,35 (9):2141~2150 |
Abstract
To thoroughly understanding the relationships between tree-ring growth and climate factors, we need to relay on tree physiological models. Simulations on one hand could enhance the interpretation of statistical response functions from a physiological view, on the other hand, may explain the divergences detected in the response function results in some cases, and thus provide additional information about climate effects on tree growth in different environments. Furthermore, the partial influences of each environment on tree growth can be quantified by models. Hence, the advantages of tree physiological models cannot be ignored when engaging in dendroclimatology or dendroecology studies. In this study, we reviewed the V-S, TREE-RING and MAIDENiso models in detail, including their physical mechanisms, operating environment, and their advantages as well as disadvantages. Specifically, with the V-S model, we monitored juniper tree growth process in the northeastern Tibetan Plateau, and found that moisture variability was the limiting factor during the main growing season. Modeled results verified statistical analysis in the study region. Compared to TREE-RING and MAIDENiso models, the physical mechanism of V-S is limited, which cannot be used to systematically analyze the process of carbon allocation, storage and remobilization, and thus, could not present tree growth responses to increasing atmospheric CO2 concentration over the past 150 years. However, the model only needs daily mean temperature and accumulated precipitation as inputs, enabling ring width simulations in any location where daily climate records exist. Therefore, V-S model has been widely used up to now. Perhaps the most complete of tree physiological models, incorporating processes linking environmental conditions to tree-ring characteristics via photosynthesis, respiration, carbon storage, and cambial processes, are TREE-RING and MAIDENiso models. They can be used to investigate the imprints of increasing CO2 concentration on tree growth. But, the main limitation of the two models is the δ18O data of daily precipitation unavailable in most regions. Overall, tree physiological models have been continuously developed during the past years, which improved our understanding of tree growth and climate factors to some extent. More detailed information in the complicated tree ecophysiological process and other non-climatic influences under current climate change context will help to the thorough improvement of tree growth models.