 2015, Vol.35
2015, Vol.35

树木生长时从大气CO2中吸收碳,从土壤和大气水分中吸收氢和氧元素及其他营养成分,进而合成树木有机体。因此,树轮稳定碳氢氧同位素比率(δ13C、 δD和 δ18O)的变化能够反映树木生长期合成和利用有机碳水光合产物时的气候和环境条件。树轮稳定同位素指标以其相对明确的分馏机制、 较高的信噪比和受年龄效应影响较小等优势[1, 2],成为古气候、 古环境和全球变化生态学研究中的一项重要代用指标[3, 4, 5, 6]。例如,在欧洲利用树轮δ13C重建了过去千年夏季日照的变化[7]; 在亚洲利用树轮 δ18O开展了千年降水变化的研究[3]。在树轮气候学传统研究手段(宽度和密度)的限制因子不明显区域(如热带地区),树轮稳定同位素技术的应用可解决热带地区树木定年困难问题[8, 9, 10],能够较好地揭示区域气候变化的信息[9],成为树轮气候学研究中的一项重要和独特的代用资料。另一方面,树轮稳定同位素研究在全球变化生态学方面发挥着巨大作用。利用树轮稳定同位素记录可以评价气候变化(如干旱变化、 CO2浓度增加)对于树木生理的影响[11, 12, 13, 14, 15, 16],可用来研究树木生理学和稳定同位素分馏机理[17, 18, 19]; 同时,树轮稳定同位素研究为气候模式中植被-大气的相互作用提供一些基本认识[20],为认识大气CO2浓度升高对气候变化的贡献和森林生态系统的动态响应提供了一种有效的途径[21]。结合树轮稳定同位素记录近期研究进展,本文系统综述和评价利用树轮稳定同位素指标进行古气候研究的成果,评述了树轮稳定同位素方法在树木生理学研究方面的进展,并指出树轮稳定同位素研究在未来需要着重发展的方向。
1 树轮稳定同位素分馏机理树木通过光合作用将CO2 和H2O 合成有机物,树木吸收CO2并通过C3途径使其中的碳原子以碳水化合物的形式进入植物的有机组织。因此,树木中碳同位素分馏的主要步骤大致可以分成3个阶段: 第一阶段发生在大气CO2通过扩散作用进入气孔腔的过程。该过程主要受风速、 大气温度、 湿度(降水量)、 大气CO2分压、 光照强度等因素的影响(图 1a); 第二阶段是CO2由气孔腔进入叶肉细胞的过程,这一过程所遇到的阻力很大。这两个阶段中的碳(C)没有发生形态上的变化。只是由于质量差异引起碳同位素动力分馏,其分馏效应约占总效应的1/3; 第三个阶段是CO2被羧基多肽酶固定,并进一步合成淀粉、 多糖、 纤维、 蛋白质、 脂肪的过程。C由无机形态转变为有机形态,同时C稳定同位素发生羧化分馏(net fractionation due to carboxylation),约占总效应的2/3。第二阶段中的C的分馏受到与树木生长密切相关的环境因子的影响。因此,稳定碳同位素比率变化记录了动力分馏时的环境信息(风速、 大气温度、 CO2浓度、 日照等); 而在羧化分馏阶段,则主要反映了与树木光合、 蒸腾强度相关联的水分利用效率变化等特性[22]。据此,Farquhar等[23]建立了C3 植物碳同位素分馏模式公式:

|
图 1 树木生理和生物化学效应的综合模型(a)稳定碳同位素分馏模型;(b)稳定氧同位素分馏模型,其中生理效应包括叶片蒸发导致的同位素富集、树干水的稳定同位素组成和土壤水流向叶片的水同位素组成是一致的; 改绘于文献[25] Fig. 1 Composite tree-ring stable isotope fractionation models of physiological and biochemical effects of trees. (a)Tree-ring stable isotope fractionation model and (b) stable oxygen isotope fractionation model in tree rings. Revised from the reference[25] |
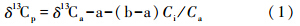
公式(1)中,下标p、 a分别代表植物和大气,a(≈4.4‰)代表CO2的扩散分馏系数,b(≈27‰)为羧化分馏系数,Ci和Ca分别代表植物叶片内部和大气CO2的浓度(图 1a)。
树木有机物中氧(O)的来源有3种:CO2、 H2O和O2。CO2和O2对树木纤维素中的氧同位素比率没有影响[24],树木有机体氢氧同位素比率由吸收利用水的稳定同位素组成决定。树木吸收的水分大部分来自于土壤,而土壤水大部分源自降水。因而推断,树轮中的稳定氧同位素记录的信号主要来自于降水氧同位素组成变化的信号[25, 26]。然而,降水稳定同位素信号被固定在树木的过程之前,存在潜在的同位素分馏过程。首先,土壤水蒸发会改变降水稳定同位素比率的信号,不同深度土壤水的稳定同位素值存在差异; 其次,树木吸收和运输水的过程中一般不会发生同位素分馏,但叶片蒸腾作用会富集18O,其值可以达到20‰[27, 28]。此外,光合作用过程也会导致同位素发生分馏[29]。生化反应过程中的氧同位素分馏是CO2-H2O 之间的氧同位素交换平衡的结果[30],组成树木纤维素的糖分和树干木质部水分发生的交换并不是完全的[31, 32](图 1b)。因此,纤维素中的 δ18O可能反映的是源水 δ18O和叶片蒸腾富集二者的混合信号[33]。蒸腾作用使得叶片18O富集,蒸腾量取决于气孔导度和水汽压差,这两者都与空气的相对湿度变化有关[32]。最终,树轮纤维素 δ18O是树木吸收水分和木质部水的氧原子交换的综合结果。因此树木 δ18O记录的信号可能主要是降水 δ18O信号和生长季的相对湿度变化,但二者的信号强度因树木生境变化有所不同[25]。
Craig-Gordon模型较好地描述了叶片表面蒸腾水的同位素组成变化[34]。叶片水的 δ18O信号受叶片周围大气水汽压和叶片内部水汽压之比(ea/ei)的影响,同样大气中水汽的 δ18O组成,茎杆水的稳定同位素比率以及动力、 平衡分馏系数都会影响到叶片水的稳定同位素比率。假定叶片水的同位素值和大汽水的同位素值是平衡的,叶片表面温度和空气的温度是一致的,就可以利用相对湿度来代替ea/ei的值[35]。Waterhouse 等[26]在上述假设的前提下将Craig-Gordon模型进行了改进,考虑到后光合作用、 树木中间营养物质(糖类和淀粉)的同位素变化以及叶片同位素蒸腾富集等因素,综合提出了植物纤维素 δ18O的分馏模型:

公式(2)中, f0为纤维素氧原子和木质部水分氧原子交换比例,通常取值在0.4~0.6之间[35],ε0是氧同位素生物分馏因子(≈27‰±3‰[36]),h为相对湿度,δ18Ocell和 δ18Os是纤维素和土壤水(降水)的 δ18O值,εk和εe为动力分馏系数和平衡分馏系数。以上模型反映了树木年轮稳定氧同位素的生物物理和化学分馏过程,为树轮稳定氧同位素记录进行古气候和环境变化研究奠定了理论基础。
2 树轮稳定同位素序列的建立树轮α-纤维素的化学成分相对比较稳定,结构和化学性质清楚,有固定的分子式,且易于从原木中提取[37]。因此,广泛采用树轮α-纤维素来分析δ13C和 δ18O。对于树轮δ13C序列的建立,一般4~6棵树的样芯就能够有效地代表样点的信息(EPS>0.85)[38, 39, 40, 41]。对于树轮 δ18O的研究表明,4~5棵树就能够保证样点的树轮 δ18O年表相对较为可靠,较好地反映样点水平的变化信息[40, 42, 43, 44, 45]。树轮δD序列建立需要5棵树样品测定就可达到EPS>0.85的要求[46, 47]。在我国干旱和半干旱地区树木的生长相对较慢,为了保证气体稳定同位素质谱分析的样品量,同时兼顾到样点序列的代表性,一般采用4~10棵树的样芯来建立树轮稳定同位素序列[48, 49]。
树轮δ13C和 δ18O序列是否存在年龄效应,是否需要去趋势也是树轮稳定同位素气候学研究所关注的问题。对于树轮δ13C,某些树种存在幼龄效应[43, 50, 51, 52]。其幼龄效应主要体现在树木幼龄期树轮δ13C呈现大约1‰~2‰的增加趋势[50, 51]。这可能是由于幼树受到林冠效应影响吸收的光减少[53],且幼树吸收较多的由土壤呼吸释放的CO2[54, 55],导致树木幼年期的δ13C偏高。另外,树木幼龄期的生理特性以及根系系统对水分的输送能力比成年期低[25],也会导致幼龄效应的产生。树轮δ13C的幼龄效应在不同树种持续的时间不同,从20年~200年不等[51, 52, 56, 57, 58]。为避免幼龄效应的影响,树轮δ13C古气候研究中大部分将树轮前20~50年舍弃[48, 51, 59, 60, 61]。
树轮 δ18O幼龄效应的报道相对较少。对法国松树的研究表明[43],该树种存在着30~40年的幼龄效应,主要表现在树轮 δ18O在幼年期呈现一个明显的下降趋势(约1.5‰)。Treydte等[3]在巴基斯坦北部建立的柏树(Juniperus excelsa和Juniperus turkestanica)年轮 δ18O序列也呈现一定幼龄效应,树轮 δ18O值在过去300年下降约3‰。树轮 δ18O的幼龄效应是由于幼树根系较浅,吸收的土壤水分受到蒸发富集的影响较大,导致树轮 δ18O偏高[62]; 老树和幼树叶片形态和功能的差别也会导致幼龄效应的发生[62]; 老幼树树冠的差别会导致其湿度梯度存在差别[63],也会引起幼龄效应。然而,在大多数研究中,树轮 δ18O并没有表现出明显的幼龄效应[44, 45, 64]。因此,利用树轮 δ18O进行气候变化研究时一般不需要去趋势,可较好地保留低频气候变化信号[43, 45]。
确定所需样芯数量后,建立树轮稳定同位素序列时就要考虑采用何种方法(单芯或混合)。树轮稳定同位素分析样品准备比较耗时,稳定同位素比率质谱测定费用相对较高。因此,原木混合后提取纤维素测定稳定同位素比率的方法相对比较经济,缺点是不能评价树木不同个体之间稳定同位素比率的差异(δ13C,1‰~3‰; δ18O,1‰~4‰; δD,5‰~30‰)[65]。通过单芯测定树轮稳定同位素比率(δ13C,δ18O和δD)后求取算术平均值和混合样芯测定结果比较表明,混合测定的结果和单芯测定的结果在平均值和序列变化上没有明显的差异(<1倍标准差)[42, 43, 47, 66]。然而,单芯测定树轮稳定同位素比率的优势是能够针对每一棵树去“诊断”其中可能的非气候信号,也可以将每棵树的稳定同位素序列中的信号进一步“挖掘”[25],例如年龄效应[43]、 气候信号强度[47, 66]等。
另一种树轮稳定同位素序列建立办法是将每棵树的同位素比率单独测定,然后在准备样品时将几年(5年或是任意年)的样品作为一个样品进行分析,最后将稳定同位素比率用线性插值法插值到年分辨率[67, 68]。这样可以有效地减少样品分析数量,保留每棵树低频变化的信息,也可以进行序列的误差估计[25, 67, 69]。Gagen等[70]提出了一种快速有效的方法(join-point)来建立长序列的树轮稳定同位素年表。如果明确活树和半化石木之间的差别及树轮δ13C幼龄效应的差别,就可以利用该方法有效地建立长序列(≥1000年)的树轮稳定同位素年表[70]。将不同分辨率的样品互相重叠的部分单独进行分析,不仅克服了数据产出效率低的缺陷,有效地保留序列的低频信号,同时又能增加序列在长时间尺度上的重复性[71]。如在我国柴达木盆地的研究,首先利用年分辨率的序列来探讨树轮稳定同位素指标的气候意义[72, 73],然后利用3年混合的样品来建立1000年的稳定同位素序列[74],有效地保证了气候低频信息和序列的重复性。
具体研究中,应依据不同研究目的来设计树轮稳定同位素序列建立方案。如进行幼龄效应的研究,就需要将每一棵树的样品进行单独测量; 如研究长期气候变化信息就可以进行混合测量,可高效地提取长时间尺度的气候信息。对于一些早晚材水源存在较大差异的区域,为了研究季节性的气候信号,可以分别测定树轮早晚材纤维素的稳定同位素比率[75, 76]。
3 树轮稳定同位素气候学最近十几年,随着气体稳定同位素质谱分析技术的发展,树轮稳定同位素记录在古气候环境变化方面的研究成果不断涌现,我国树轮稳定同位素气候学研究也取得快速发展(图 2a)。

|
图 2 (a)中国树轮稳定同位素研究开展区域示意图; (b)新疆北部干旱变化重建[82]; (c)藏东南区域云量重建[83]; (d)长白山安图树轮δ13C[84]; (e)长白山安图树轮δD[84]; (f)秦岭黄陵树轮δ13C[81]; (g)福建长汀树轮 δ18O[85] Fig. 2 (a)Tree-ring stable isotopes researches over China; (b)SPEI variability in north part of the Xinjiang Province[82]; (c)variability in cloud cover in southeastern Tibetan Plateau[83]; (d)tree-ring δ18O and(e)δD at Antu County site of the Changbai Mountains[84]; (f)tree-ring δ13C at the site in the Huangling County of Qinling Mountains[81]; (g)tree-ring δ18O from Fokienia hodginsii trees in the Changting County, Fujian Province[85] |
树轮δ13C的分馏主要受气孔导度和光合速率的影响[22, 23]。气孔导度主要受水分条件(相对湿度、 土壤湿度和干旱等)的限制; 光合速率主要受到温度、 辐射强度等影响[25]。因此,树轮δ13C可以用于温度、 辐射、 降水量、 干旱等气象要素的重建。
树轮δ13C重建的温度变化是全球变暖的有力证据之一。欧洲北部,树轮δ13C主要反映出夏季平均温度的变化[58, 77]; 挪威西北部,由于云量较多,温度和辐射强度存在一定差异,树轮δ13C分馏主要受到辐射强度影响[78]。树轮气候学家基于树轮δ13C分别重建了欧洲北部千年以来的夏季平均温度和辐射强度变化[79, 80],为欧洲北部过去千年以来温度变化提供了重要科学数据。基于23个样点的欧洲树轮δ13C网络研究表明,树轮δ13C主要反映温度变化信号。在我国陕西黄陵的油松(Pinus tabulaeformis)树轮δ13C记录了温度信号[81],这也是在我国首次开展的树轮稳定同位素方面的研究工作。秦岭北坡,油松树轮δ13C与5~7月份平均温度相关关系最为密切,利用树轮δ13C揭示了20世纪的温度升高及其变化机制[49](图 2)。
树轮δ13C在一些区域还重建了降水量、 干旱变化等信息[82, 83, 84, 85]。北美树轮δ13C网络主要反映了夏季季风降水的变化[86, 87],成为了研究北美季风降水和干旱变化的重要指标。在欧洲阿尔卑斯山区,Kress 等[64]利用落叶松(Larix gmelinii)树轮δ13C研究了该地区过去350年以来的干旱历史。瑞士亚高山区,云杉(Picea asperata)树轮δ13C主要反映了降水的变化[88]。在热带地区,树轮δ13C与全年降水量存在显著的负相关,成为重建过去降水变化的有效工具[89]。
青藏高原作为气候变化的敏感区,地形复杂,树轮δ13C的气候意义也不尽相同。藏东南地区,冷杉(Abies fabri)和云杉树轮δ13C主要记录了降水变化的信息[90, 91]。在青藏高原东北部,祁连圆柏(Sabina przewalskii Kom.)树轮δ13C主要反映了生长季平均温度的变化[59, 72, 92],而青海云杉树轮δ13C对干旱的响应更为敏感[93]。青藏高原西部,圆柏树轮δ13C也主要记录了夏季平均温度的变化,Treydte等[94]基于树轮δ13C重建了巴基斯坦北部过去1000年以来的温度变化历史。树轮δ13C的研究为认识青藏高原过去千年以上气候变化提供了有利条件。
3.2 树轮稳定氧同位素(δ18O)树轮 δ18O记录研究在最近十几年得到快速发展[3, 83]。源水(降水)δ18O的变化受温度制约[95],因此树轮 δ18O的变化理论上可以反映温度变化[96]。然而,温度信号可能会在叶片水蒸发富集的过程中减弱或丢失。因此,树轮 δ18O在不同的气候区域,既用于温度重建[48],也可用来进行降水量、 相对湿度、 干旱指数以及云量等古气候重建 [3, 73, 75, 97, 98, 99](图 2)。
欧洲和北美地区的树轮 δ18O气候学研究开展较早,已建立了比较完善的树轮 δ18O记录网络。欧洲大陆不同气候区域下23个样点的树轮 δ18O的气候信号在区域大尺度上比较一致[100]。Saurer 等[101]沿经度对欧亚大陆北部从挪威到西伯利亚的断面树轮 δ18O序列的分析表明,树轮 δ18O的值和水汽 δ18O的值在空间上分布一致。以上研究表明,利用树轮 δ18O网络数据可以实现区域大尺度上气候要素的重建。
亚洲地区的树轮 δ18O气候学研究起步较晚,但发展很快。在长时间尺度的树轮 δ18O研究取得了显著成果[3]。在高亚洲西部,树轮 δ18O重建了过去一千年以来的降水变化,指出20世纪是过去1000年以来降水量最多的一个世纪[3]。青藏高原中部,利用树轮 δ18O重建了过去800年夏季降水量变化,表明小冰期气候湿润,而20世纪后半叶为过去800年中最为干旱的时段[102]。在青藏高原东南部树轮 δ18O反映出区域自1860年来干旱化的趋势[98, 103, 104],Liu等[83]基于区域多样点树轮 δ18O网络资料得出夏季风减弱是导致区域1860s以来的干旱化的主要原因(图 2c)。青藏高原东北部,祁连圆柏树轮 δ18O反映了过去千年以来的相对湿度变化,表现出中世纪暖期气候较为干旱,小冰期气候湿润以及20世纪干旱的低频干湿变化型(图 3),为揭示该地区干湿变化历史提供了有效代用资料[74]。在新疆北部地区,树轮 δ18O网络的建立成为了解北疆地区过去300多年干旱变化及其驱动机制的重要资料[99, 105, 106]。亚洲东南部,树轮 δ18O的研究已经大量开展,利用树轮 δ18O分别重建了过去的降水变化[107, 108]、 干旱变化[109, 110]和相对湿度变化[111]。

|
图 3 青藏高原东北部990~2010A.D. 期间3年分辨率祁连圆柏 δ18O表示的相对湿度变化[74]点线代表树轮 δ18O均值±1倍标准差(σ),红色直线代表均值线,蓝色粗线为100年FFT平滑线 Fig. 3 The tree-ring δ18O series(3-year temporal resolution)represent summer relative humidity for northeastern of Qinghai-Tibetan Plateau from 991 A.D. to 2010 A.D.[74]. Horizontal dashed lines denote mean ±σ(one standard deviation). Thick red horizontal lines denote long-term mean values while thick blue lines denote the smoothed results using a 100-year fast Fourier transform (FFT) filter to emphasize century-scale variation |
此外,可利用树轮 δ18O来分析大气环流在长时间尺度上的变化,并分析这种变化产生的机制和原因。亚洲季风区北边缘区的贺兰山树轮 δ18O可以指示东亚夏季风降水的变化[112, 113]; 藏东南地区的树轮 δ18O为了解南亚季风变化及ENSO变化提供了可能[103, 110]。在美国东南沿海,树轮 δ18O记录能够很好的反映热带气旋强度及其变化[114]。
3.3 树轮稳定氢同位素(δD)与树轮稳定碳氧同位素研究相比,树轮δD气候学的研究结果较少[47, 111, 115]。一方面与树轮δD的测定难度和硝化纤维准备过程较繁琐有关; 另一方面,树轮δD分馏过程的复杂性也限制了其在气候变化研究中的应用[26, 32]。树轮稳定氢氧同位素虽然在源水和分馏机制上类似,但是两者在生物化学分馏过程中的差异导致所反映的气候信息不尽相同[26, 77, 116]。早期,树轮δD主要用于表示古气温的变化[84, 115, 117, 118]。我国长白山的树轮δD反映了前一年的温度变化信息,并能在一定程度上揭示季风强弱和区域温度变化的信息[84](图 2e)。我国西部川西高原地区的研究表明,树轮δD的变化主要受到生长季相对湿度的影响[111],但中国西北干旱区的树轮δD反映冬季温度信号更强,并且与北极涛动存在着一定的联系[47/a>]。因此,树轮δD不仅能反映温度变化,还与湿度变化存在一定的关系。然而,干旱区树轮δD与气候要素相关关系较弱,现阶段很难准确利用其重建过去气候变化历史。近年来,树轮δD的研究大多与树轮δ13C和 δ18O综合[111],多个代用指标综合应用能够更为准确的分析局地和区域气候以及大气环流在不同时空尺度上的特征和变化[77, 105, 119],这也是树轮稳定同位素气候学发展的趋势。
3.4 树轮稳定同位素年内高分辨率记录树轮稳定同位素年内变化型为热带地区树木定年提供了一个有效的方法(稳定同位素比率的年内循环周期可作为定年的依据)[10],对揭示树轮稳定同位素的生理机制、 完善同位素分馏模型以及解释同位素记录的气候信号有极大帮助。近年来树轮稳定同位素年内变化的研究有以下几个特点:1)主要集中于热带地区,欧洲和美国一些温带和亚热带地区; 2)研究的树种有限,对阔叶树(如柚木(Tectona grandis)、 云雾林(Cloud forest)、 栎树(Quercus Linn)和栖槐(Sequoias empervirens)等)的研究较多,针叶树研究相对较少; 3)研究主要集中在 δ18O和δ13C方面,对δD的研究较少[120]。
全木和纤维素年内的δ13C值呈先升后降的变化特征,且δ13C逐年变化并非连续[2]。阔叶树树轮δ13C存在明显的年内变化型: 在生长初期呈现急剧上升趋势,生长中期呈现显著下降趋势,而在生长季后期存在一个短暂的上升(图 4)[121]。这3个阶段的生理意义在于: 第一阶段δ13C值的上升是由于树木利用前一年积累的光合产物。树木优先消耗12C,使得储存物质中13C富集,从而出现上升趋势; 第二阶段,树木开始利用当年生产的糖类物质(13C值相对亏损)。根据同位素混合原理,随着利用当年生产的糖类物质增加,树轮δ13C呈现下降趋势; 第三阶段中,由于树木叶片的凋落,树木又开始利用夏季储存下来的光合产物,导致树轮δ13C上升[122, 123, 124]。树轮δ13C年内显著的变化也可能由于局地小环境的气象条件改变[125, 126],或利用前一年光合产物引起[127, 128, 129]。13CO2示踪法[122, 130] 和树轮δ13C数理模型的方法[124]证明树轮δ13C年内变化在一定程度上受前期积累的碳水化合物的影响。但是树种间也存在差异,如罗汉松(Podocarpus macrophyllus)受前3年储存物质的影响,长穗巴豆木(Croton macrostachyus)仅受前一年储存物质的影响[130]。大多研究认为常绿树种树轮δ13C年内变化可能反映气候要素的变化[2, 127, 131]。常绿树种树轮δ13C年内最大值与最大水汽压差、 最高温、 最低土壤水汽、 干旱变化和最大δ13CO2等环境因子密切相关[1, 60, 82, 125, 126, 132, 133]。Schubert 和Jahren[131]对全球15个地点的常绿树种树轮δ13C年内变化研究发现阔叶树种和针叶树种存在不同的年内变化型(图 4a和4b),常绿树种树轮δ13C年内变化主要由大气中CO2的δ13C以及降水的季节变化引起。在藏东南色季拉山的研究也发现长苞冷杉(Abies georgei)树轮δ13C存在明显的年内变化型(数据未发表)(图 4c)。

|
图 4 阔叶树(a)和针叶树(b)树轮δ13C年内变化型[2, 121]以及藏东南色季拉山长苞冷杉(c)树轮δ13C年内变化(数据未发表) Fig. 4 Inner-annual variability patterns of tree-ring δ13C for (a) broad-leaved tree and (b) needle ever-green trees[2, 121],(c)inner-annual variations of tree-ring δ13C at Sygera Mountains in southeastern Tibetan Plateau |
不论是针叶树还是阔叶树的树轮 δ18O,受前一年光合产物的影响都不明显。因此,树轮 δ18O在气候环境季节变化的研究中具有很大的优势。树轮 δ18O的年内变化可以揭示降水 δ18O[10, 134, 135, 136, 137]、 季风降水[138, 139, 140]、 厄尔尼诺和南方涛动[135]、 雾发生的频率[9, 141, 142]、 相对湿度[139, 143, 144, 145]、 气温[143]、 干旱事件[146]以及热带气旋[147]。树轮 δ18O年内变化在研究特殊环境气候事件具有独特的优势。Evans和Schrag[10]通过研究美洲不同气候区的树轮 δ18O的年内变化发现,树轮 δ18O 在1998年内出现的低值能够反映当年的ENSO事件。Roden等[148]在美国加州地区发现栖槐(Sequoia sempervirens)树轮 δ18O的年内变化与当地雾的频率的季节变化存在很好的一致性。Li等[147]在美国田纳西州研究中发现树轮 δ18O年内变化记录了飓风事件。
近年来,关于树轮 δ18O年内变化的研究日益增多,其中热带地区的研究成果最为丰富。Evans[149]在哥斯达黎加运用模型的方法发现树轮 δ18O年内变化和降水有很好的一致性,树轮 δ18O很好地反映ENSO事件,并能解释蒙特金蟾灭绝的原因[141]。Managave等[140]报道在印度南部的树轮 δ18O年内变化主要受季风降水影响,并存在特殊的年内变化型,这种变化型可能由两种季风系统(西南季风和东北季风)交替影响引起的。在印度中部柚木树轮 δ18O的年内变化受降水和相对湿度的控制[140, 150]。Zhu等[136]利用年内变化的树轮 δ18O重建了泰国北部100多年以来的降水量年内变化。柬埔寨树轮 δ18O的年内变化与降水量存在很好的相关性,能很好地表征印度-太平洋暖槽(IPWP)的移动以及ENSO活动[135]。在藏东南色季拉山冷杉树轮 δ18O的年内变化很好地揭示了印度季风降水 δ18O的季节变化(数据未发表)。在印度尼西亚,树轮 δ18O季节变化主要受降水控制,在干季和湿季交替影响下发生周期性变化[151]。亚洲热带和亚热带地区的树木生长较快,有利于树轮稳定同位素年内变化型的分析。同时,季风期和非季风期气候差异较大,树轮 δ18O年内变化序列中可能包含更多的季节上的气候变化信息。
4 树轮稳定同位素记录与碳循环在气候模式中需要考虑到植物与大气之间碳-水循环的相互作用,树轮稳定碳氧同位素作为生物体和大气CO2浓度变化相互作用的良好记录载体。因此,树轮稳定碳氧同位素记录相结合能够为气候模式中植物-大气的相互作用提供基础认识并对大气模式模拟结果进行检验[16, 20]。Saurer等[20]利用区域的树轮δ13C资料计算出区域水分利用效率(Intrinsic Water-use Efficiency,简称iWUE)的变化和植被-大气模式(LPX-bern model)得到的结果进行比较,揭示出由于CO2浓度升高和气候变化导致土壤水分减少,植被水分利用效率在20世纪提高。树轮δ13C记录为气候变化中森林碳-水-能量循环提供了验证数据[16],对于气候模式中CO2的贡献提供了科学认识。
植物叶片气孔的气体交换以及生长量的改变是树木对于CO2浓度增加最主要的响应方式[152]。过去100年大气CO2浓度急剧的增加,从1920年的303 ppm增加到2011年的391 ppm[153]。FACE(Free-air Carbon Dioxide Enrichment Experiment)实验结果表明,CO2浓度的增加会导致树木iWUE的增加,进而对于树木生长产生一定的促进作用[154, 155]。相比于短期实验,树轮δ13C表征了树木长期iWUE的变化,更能反映自然生长条件下树木对CO2浓度升高的响应[156, 157]。树轮δ13C计算的iWUE在过去40年中增长了20.5%,并且在亚热带到寒带不同类型的森林区没有显著区别[156]。北半球中高纬度的83条树轮δ13C表征的树木iWUE在过去250年平均增长了30%,显著增加是从1950年开始[13],反映了气候变化背景下碳循环变化的一个重要方面。
树木对于CO2浓度增加的响应方式会随时间发生改变,这种响应将与其他的环境要素(气候变暖)产生交互作用[158]。目前,多数研究对自然生长条件下树木生长对于CO2浓度升高以及气候变暖如何响应进行了分析[11, 14, 156, 157, 159, 160, 161, 162]。部分研究发现,20世纪树木iWUE呈现出明显的增长趋势,该增长趋势可能是由于CO2浓度急剧增加导致[160, 161, 162]。
然而,iWUE的增加是否会导致树木生长的增加则在不同区域和不同树种间存在差异。在相对湿润的温带森林区、 高山林线处树木生长则呈现出显著增加[160, 163, 164],而在热带、 相对干旱的温带森林以及海拔下限处树木生长则表现出了下降[11, 157, 165, 166]。但是,位于地中海地区的西班牙刺柏(Juniperus formosana)即使处于较为干旱的气候环境之中,iWUE的增加显著地促进了树木生长[167]; 位于黑河流域的河岸胡杨林(Populus euphratica)也表现出了显著的CO2浓度升高产生的促进作用[14]。由此可见,较大范围内iWUE的增加并不会等同的完全促进树木的增长,多样点的树木生长与iWUE的综合分析也表明CO2浓度升高产生的肥化效应可能不足以抵消气候变化带来的其它负面因素的影响,例如: 干旱,营养缺乏以及光合适应现象[156, 165]。
利用树轮资料研究树木生长对CO2浓度升高如何响应时,分析方法和研究手段均趋于定量与定性相结合。在分析方法上,RC(Response Contrast)值的提出能够较好的定量化评价树木断面积指数(BAI)与iWUE在某一时间段内相对累积变化,便于比较不同样点以及不同时段的树木生长状况[14, 165]。除利用BAI值与iWUE的一般线性回归来分析CO2浓度升高是否促进树木的生长外[11, 14, 157],还可以利用一般线性混合模型来建立树木生长与iWUE、 气候变量以及立地因子(如: 树木密度、 树木胸径)关系,从而定量评估iWUE、 气候变量对树木生长变化产生的贡献,分析立地因素对树木生长变化存在的影响是否显著。将树木对CO2浓度升高响应的理论情景纳入分析,可以定性解释树木iWUE的增加方式是属于主动响应、 被动响应以及最强响应的哪一种,为分析当前气候背景下树木生理过程变化具有指导作用[14, 159, 168]。从研究手段来看,树轮δ13C并不能说明iWUE变化是否来自于水分状况的改变,因为光合速率(A)和气孔导度(g)的改变都会导致树轮δ13C以及iWUE的改变[169],而树轮 δ18O只记录g的变化。因此,多种同位素分析手段的结合可以较好的探讨树木生长对于CO2浓度升高与气候变暖的响应,分析导致树木生长变化的主要原因(图 5)[11, 14, 165, 167, 170]。

|
图 5 利用树轮代用资料分析树木生长动态变化 Fig. 5 Tree growth dynamics investigation based on tree-ring proxies |
利用树轮同位素资料进行森林生长动态变化及碳循环研究存在着一定的优越性以及不确定性。主要有:1)树木生长变化区域性比较明显,树木的生长没有呈现出预期的增长趋势[157, 170],这与一些生态系统模型的假设和输出结果相悖,模型模拟认为由于CO2浓度增加陆地生态系统碳储量在未来半个世纪内会显著增加[171, 172],而树轮资料则成为可靠的代用资料来揭示树木对于长期环境变化的响应方式。2)iWUE的增加速率随着时间减小,树木对CO2浓度升高响应的敏感性降低,表现出了一定的饱和效应[157, 173]。这种饱和效应被认为是由于极端干旱所引起,但这种饱和现象并没有纳入生态系统模型之中。现有模型普遍认为iWUE会随着CO2浓度的增加持续增长[174]。如果饱和效应的出现导致较大区域尺度上树木生长持续出现降低趋势,那么陆地生态系统的碳储量可能会高估[157],而树轮稳定同位素资料能够较好地补充完善模型的一些假设条件以及参数设定。3)树木生长也可能由营养元素的丢失而引起,在CO2增加的背景下,森林生产力也会受到营养缺失的限制,尤其是氮(N)元素[175]。N沉降能促进森林生产力[176],但是也会引起土壤酸化,导致其他营养元素的丢失,进而限制森林生产力的增加[177]。树轮氮同位素(δ15N)的引入可以用来分析这一问题[178]。因此,未来研究中,需要利用多指标记录,来区分树木生长对CO2浓度增加响应过程中资源限制的作用(例如: 水分、 营养元素等)。
5 结论与展望树轮稳定同位素代用资料在古气候和古环境研究中具有重大潜力,能够较好地保留低频信号。在古气候学研究中,树轮稳定同位素资料成为一项独特且重要的代用资料。在中国西部干旱、 半干旱区,树轮稳定碳同位素能较好地反映区域温度的变化,树轮稳定氧同位素则能够较好解释区域干湿变化。因此,在未来的工作中,在干旱、 半干旱区可以利用树轮稳定碳同位素记录进行区域温度变化重建,利用树轮稳定氧同位素记录进行区域干湿变化的研究。在湿润区树轮稳定碳氧同位素相结合可开展树木生理响应和气候变化的相关研究; 同时,可以在亚热带区域开展树轮稳定氧同位素高分辨率的研究,来分析极端气候事件的发生及其影响。
在欧美,树轮稳定同位素气候学研究从点到面的工作已经开展,中国现有的树轮气候学的工作大多集中在样点或小区域气候重建方面,应尽快开展区域网格资料的综合研究。另外,多个代用指标的综合研究也已成为一个趋势,可完善和补充单个指标在特殊时段代用指标气候意义发生转变或不明确的情形。现有成果表明,树轮稳定碳氧同位素能够在较大空间范围内反映较为一致的气候信息。树轮稳定同位素网格资料的建立有利于分析长时间尺度上控制我国气候的多个季风环流子系统的影响程度及其空间变化。相对于宽度资料来说,树轮稳定同位素长序列资料相对短缺,应重点在我国具有建立长序列潜力的典型地区开展工作,完善树轮稳定同位素网络的建立,进行区域气候变化的综合性研究工作,同时兼顾多个科学问题设计研究方案。
稳定同位素分馏机理的明确为多同位素指标的应用提供理论基础。树轮稳定同位素分馏机理研究亟须加强,特别是稳定氢同位素分馏过程。此方面工作开展可解决树轮长序列氢氧同位素序列气候意义的解释问题,如为何湿润区树轮稳定氢氧同位素反映相对一致的气候信息,但在干旱区和半干旱区树轮稳定氢氧同位素序列反映气候信息则存在较大差异,导致这种差异的原因也需要明确。新方法的应用,如碳链不同位置氢氧同位素信息(position stable isotope),细胞尺度同位素信号的识别等均有助于树轮稳定同位素分馏机理的完善及其记录信号的分离。树轮稳定同位素记录在生态学方面的研究也应该进一步加强。结合全球碳循环研究、 树轮记录反映的生长量变化以及树轮稳定同位素记录,可为全球变化背景下森林动态变化提供可靠的科学依据。
| 1 | Wilson A T, Grinsted M J. 12 C/13 C in cellulose and lignin as palaeothermometers. Nature, 1977,265 (5590):133~135 |
| 2 | Leavitt S W, Lone A. Seasonal stable-carbon isotope variability in tree rings:Possible paleoenvironmental signals. Chemical Geology, 1991,87 (1):59~70 |
| 3 | Treydte K S, Schleser G H, Helle G et al. The twentieth century was the wettest period in Northern Pakistan over the past millennium. Nature, 2006,440 (7088):1179~1182 |
| 4 | Gagen M, McCarroll D, Loader N J et al. Stable isotopes in dendroclimatology:Moving beyond 'potential’ dendroclimatology. In:Hughes M K, Swetnam T W, Diaz H F eds. Dendroclimatology Progress and Prospects. Netherlands:Springer, 2011. 147~172 |
| 5 | Young G H F, McCarroll D, Loader N J et al. A 500-year record of summer near-ground solar radiation from tree-ring stable carbon isotopes. The Holocene, 2010,20 (3):315~324 |
| 6 | van Der Sleen P, Groenendijk P, Vlam M et al. No growth stimulation of tropical trees by 150 years of CO2 fertilization but water-use efficiency increased. Nature Geoscience, 2015,8 (1):24~28 |
| 7 | Loader N J, Young G H F, Grudd H et al. Stable carbon isotopes from Torneträsk, Northern Sweden provide a millennial length reconstruction of summer sunshine and its relationship to Arctic circulation. Quaternary Science Reviews, 2013, 62 :97~113 |
| 8 | Loader N J, Walsh R P D, Robertson I et al. Recent trends in the intrinsic water-use efficiency of ringless rainforest trees in Borneo. Philosophical Transactions of the Royal SocietyB: Biological Sciences, 2011,366 (1582):3330~3339 |
| 9 | Anchukaitis K J, Evans M N, Wheelwright N T et al. Stable isotope chronology and climate signal calibration in neotropical montane cloud forest trees. Journal of Geophysical Research:Biogeosciences, 2008,113 (G3):G03030 |
| 10 | Evans M N, Schrag D P. A stable isotope-based approach to tropical dendroclimatology. Geochimica et Cosmochimica Acta, 2004,68 (16):3295~3305 |
| 11 | Lévesque M, Siegwolf R, Saurer M et al. Increased water-use efficiency does not lead to enhanced tree growth under xeric and mesic conditions. New Phytologist, 2014,203 (1):94~109 |
| 12 | Keenan T F, Hollinger D Y, Bohrer G et al. Increase in forest water-use efficiency as atmospheric carbon dioxide concentrations rise. Nature, 2013,499 (7458):324~327 |
| 13 | Wang Guoan, Feng Xiahong. Response of plants' water use efficiency to increasing atmospheric CO2 concentration. Environmental Science and Technology, 2012,46 (16):8610~8620 |
| 14 | Liu Xiaohong, Wang Wenzhi, Xu Guobao et al. Tree growth and intrinsic water-use efficiency of inland riparian forests in Northwestern China:Evaluation via δ 13 C and δ18O analysis of tree rings. Tree Physiology, 2014,34 (9):966~980 |
| 15 | Walker X J, Mack M C, Johnstone J F. Stable carbon isotope analysis reveals widespread drought stress in boreal black spruce forests. Global Change Biology, 2015, doi:10.1111/gcb.12893 |
| 16 | De Kauwe M G, Medlyn B E, Zaehle S et al. Forest water use and water use efficiency at elevated CO2:A model-data intercomparison at two contrasting temperate forest FACE sites. Global Change Biology, 2013,19 (6):1759~1779 |
| 17 | Treydte K, Boda S, Graf Pannatier E et al. Seasonal transfer of oxygen isotopes from precipitation and soil to the tree ring:Source water versus needle water enrichment. New Phytologist, 2014,202 (3):772~783 |
| 18 | Larcher L, Hara-Nishimura I, Sternberg L. Effects of stomatal density and leaf water content on the 18 O enrichment of leaf water. New Phytologist, 2015,206 (1):141~151 |
| 19 | Hommel R, Siegwolf R, Saurer M et al. Drought response of mesophyll conductance in forest understory species-impacts on water-use efficiency and interactions with leaf water movement. Physiologia Plantarum, 2014,152 (1):98~114 |
| 20 | Saurer M, Spahni R, Frank D C et al. Spatial variability and temporal trends in water-use efficiency of European forests. Global Change Biology, 2014,20 (12):3700~3712 |
| 21 | Boucher, Guiot J, Hatté C et al. An inverse modeling approach for tree-ring-based climate reconstructions under changing atmospheric CO2 concentrations. Biogeosciences, 2014,11 (12):3245~3258 |
| 22 | Farquhar G D, Ehleringer J R, Hubick K T. Carbon isotope discrimination and photosynthesis. Annual Review of Plant Biology 1989,40 (1):503~537 |
| 23 | Farquhar G D, O'leary M H, Berry J A. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Australian Journal of Plant Physiology, 1982,9 (2):121~137 |
| 24 | Deniro M J D, Epstein S. Relationship between oxygen isotope ratios of terrestrial plant cellulose, carbon dioxide and water. Science, 1979,204 (4388):51~53 |
| 25 | McCarroll D, Loader N J. Stable isotopes in tree rings. Quaternary Science Reviews, 2004,23 (7~8):771~801 |
| 26 | Waterhouse J S, Switsur V R, Barker A C et al. Oxygen and hydrogen isotope ratios in tree rings: How well do models predict observed values?Earth and Planetary Science Letters, 2002,201 (2):421~430 |
| 27 | Saurer M, Robertson I, Siegwolf R et al. Oxygen isotope analysis of cellulose:An interlaboratory comparison. Analytical Chemistry, 1998,70 (10):2074~2080 |
| 28 | Saurer M, Siegwolf R, Borella S et al. Environmental information from stable isotopes in tree rings of Fagus sylvatica. In:Beniston M, Innes J L eds. The Impacts of Climate Variability on Forests. Berlin Heidelberg:Springer, 1998.241~253 |
| 29 | Ehleringer J R, Dawson T E. Water uptake by plants: Perspectives from stable isotope composition. Plant Cell Enviroment, 1992,15 (9):1073~1082 |
| 30 | Sternberg L, Deniro M J D. Biogeochemical implications of the isotopic equilibrium fractionation factor between the oxygen atoms of acetone and water. Geochimica et Cosmochimica Acta, 1983,47 (12):2271~2274 |
| 31 | Anderson W T, Bernasconi S M, Mckenzie J A et al. Model evaluation for reconstructing the oxygen isotopic composition in precipitation from tree ring cellulose over the last century. Chemical Geology, 2002,182 (2~4):121~137 |
| 32 | Roden J S, Lin G, Ehleringer J R. A mechanistic model for interpretation of hydrogen and oxygen isotope ratios in tree-ring cellulose. Geochimica et Cosmochimica Acta, 2000,64 (1):21~35 |
| 33 | Sternberg L, Pinzon M C, Anderson W T et al. Variation in oxygen isotope fractionation during cellulose synthesis:Intramolecular and biosynthetic effects. Plant, Cell and Environment, 2006,29 (10):1881~1889 |
| 34 | Craig H, Gordon L I. Deuterium and oxygen 18 variations in the ocean and marine atmosphere. In:Tongiogi E ed. Proceed of Stable Isotopes in Oceanographic Studies and Paleotemperatures.Italy:Pisa, 1965. 9~130 |
| 35 | Sternberg L. Oxygen stable isotope ratios of tree-ring cellulose:The next phase of understanding. New Phytologist, 2009,181 (3):553~562 |
| 36 | Deniro M J, Epstein S. Isotopic composition of cellulose from aquatic organisms. Geochimica et Cosmochimica Acta, 1981,45 (10):1885~1894 |
| 37 | Leavitt S W, Danzer S R. Method for batch processing small wood samples to holocellulose for stable-carbon isotope analysis. Analytical Chemistry, 1993,65 (1):87~89 |
| 38 | Leavitt S W, Long A. Sampling strategy for stable carbon isotope ananlysis of tree in pine. Nature, 1984,311 (5982):145~147 |
| 39 | McCarroll D, Pawellek F. Stable carbon isotope ratios of latewood cellulose in Pinus sylvestris from Northern Finland:Variability and signal-strength. The Holocene, 1998,8 (6):675~684 |
| 40 | Daux V, Edouard J L, Masson-Delmotte V et al. Can climate variations be inferred from tree-ring parameters and stable isotopes from Larix decidua?Juvenile effects, budmoth outbreaks, and divergence issue. Earth and Planetary Science Letters, 2011,309 (3~4):221~233 |
| 41 | Liu Yu, Ma Limin, Leavitt S W et al. A preliminary seasonal precipitation reconstruction from tree-ring stable carbon isotopes at Mt. Helan, China, since AD 1804. Global and Planetary Change, 2004,41 (3~4):229~239 |
| 42 | Liñn I D, Gutiérrez E, Helle G et al. Pooled versus separate measurements of tree-ring stable isotopes. Science of the Total Environment, 2011,409 (11):2244~2251 |
| 43 | Szymczak S, Joachimski M, Bräuning A et al. Are pooled tree ring δ 13 C and δ18O series reliable climate archives?——A case study of Pinus nigra spp. laricio(Corsica/France). Chemical Geology, 2012, 308~309 :40~49 |
| 44 | Shi Chunming, Masson-Delmotte V, Risi C et al. Sampling strategy and climatic implications of tree-ring stable isotopes on the southeast Tibetan Plateau. Earth and Planetary Science Letters, 2011,301 (1~2):307~316 |
| 45 | Young G H F, Demmler J C, Gunnarson B E et al. Age trends in tree ring growth and isotopic archives:A case study of Pinus sylvestris L. from Northwestern Norway. Global Biogeochemical Cycles, 2011,25 (2):GB2020 |
| 46 | Dodd J P, Patterson W P, Holmden C et al. Robotic micromilling of tree-rings:A new tool for obtaining subseasonal environmental isotope records. Chemical Geology, 2008,252 (1~2):21~30 |
| 47 | Liu Xiaohong, An Wenling, Treydte K et al. Pooled versus separate tree-ring δD measurements, and implications for reconstruction of the Arctic Oscillation in Northwestern China. Science of the Total Environment, 2015, 511 :584~594 |
| 48 | Liu Xiaohong, Shao Xuemei, Liang Eryuan et al. Climatic significance of tree-ring δ18O in the Qilian Mountains, Northwestern China and its relationship to atmospheric circulation patterns. Chemical Geology, 2009,268 (1~2):147~154 |
| 49 | Liu Yu, Wang Yanchao, Li Qiang et al. Tree-ring stable carbon isotope-based May-July temperature reconstruction over Nanwutai, China, for the past century and its record of 20th century warming. Quaternary Science Reviews, 2014, 93 :67~76 |
| 50 | Leavitt S W, Long A. The global biosphere as net CO2 source or sink: Evidence from carbon isotopes in tree rings. In:Caldwell D E, Brierley J A, Brierley C L eds. Planetary Ecology. New York:Van Nostrand Reinhold, 1985. 89~99 |
| 51 | Freyer H. On the 13 C record in tree rings. Part Ⅰ. 13 C variations in northern hemispheric trees during the last 150 years. Tellus, 1979,,31 (2):124~137 |
| 52 | Gagen M, McCarroll D, Robertson I et al. Do tree ring δ 13 C series from Pinus sylvestris in Northern Fennoscandia contain long-term non-climatic trends?Chemical Geology, 2008,252 (1~2):42~51 |
| 53 | Francey R J, Farquhar G D. An explanation of 13 C/12 C variations in tree rings. Nature, 1982,297 (5861):28~31 |
| 54 | Saurer M, Cherubini P, Bonani G et al. Tracing carbon uptake from a natural CO2 spring into tree rings:An isotope approach. Tree Physiology, 2003,23 (14):997~1004 |
| 55 | Schleser G H, Jayasekera R. δ 13 C-variations of leaves in forests as an indication of reassimilated CO2 from the soil. Oecologia, 1985,65 (4):536~542 |
| 56 | Craig H. Carbon-13 variations in Sequoia rings and the atmosphere. Science, 1954,119 (3083):141~143 |
| 57 | Stuiver M, Burk R L, Quay P D. 13 C/12 C ratios in tree rings and the transfer of biospheric carbon to the atmosphere. Journal of Geophysical Research, 1984,89 (D7):11731~11748 |
| 58 | Gagen M, McCarroll D, Loader N J et al. Exorcising the 'segment length curse’: Summer temperature reconstruction since AD 1640 using non-detrended stable carbon isotope ratios from pine trees in Northern Finland. The Holocene, 2007,17 (4):435~446 |
| 59 | Xu Guobao, Chen Tuo, Liu Xiaohong et al. Summer temperature variations recorded in tree-ring δ 13 C values on the northeastern Tibetan Plateau. Theoretical and Applied Climatology, 2011,105 (1~2):51~63 |
| 60 | Leavitt S W, Long A. Stable carbon isotope chronologies from trees in the Southwestern United States. Global Biogeochemical Cycles, 1988,2 (3):189~198 |
| 61 | Saurer M, Siegenthaler U, Schweingruber F. The climate-carbon isotope relationship in tree rings and the significance of site conditions. TellusB, 1995,47 (3):320~330 |
| 62 | Marshall J D, Monserud R A. Co-occurring species differ in tree-ring δ18O trends. Tree Physiology, 2006,26 (8):1055~1066 |
| 63 | Labuhn I, Daux V, Pierre M et al. Tree age, site and climate controls on tree ring cellulose δ18O:A case study on oak trees from south-western France. Dendrochronologia, 2014,32 (1):78~89 |
| 64 | Kress A, Saurer M, Siegwolf R T W et al. A 350 year drought reconstruction from Alpine tree ring stable isotopes. Global Biogeochemical Cycles, 2010,24 (2):GB2011 |
| 65 | Leavitt S W. Tree-ring C-H-O isotope variability and sampling. Science of the Total Environment, 2010,408 (22):5244~5253 |
| 66 | Liu Yu, Wang Ruiyuan, Leavitt S W et al. Individual and pooled tree-ring stable-carbon isotope series in Chinese pine from the Nan Wutai region, China:Common signal and climate relationships. Chemical Geology, 2012, 330~331 :17~26 |
| 67 | Woodley E J, Loader N J, McCarroll D et al. Estimating uncertainty in pooled stable isotope time-series from tree-rings. Chemical Geology, 2012, 294 :243~248 |
| 68 | Boettger T, Friedrich M. A new serial pooling method of shifted tree ring blocks to construct millennia long tree ring isotope chronologies with annual resolution. Isotopes in Environmental and Health Studies, 2009,45 (1):68~80 |
| 69 | Mayr C, Frenzel B, Friedrich M et al. Stable carbon-and hydrogen-isotope ratios of subfossil oaks in Southern Germany:Methodology and application to a composite record for the Holocene. The Holocene, 2003,13 (3):393~402 |
| 70 | Gagen M, McCarroll D, Jalkanen R et al. A rapid method for the production of robust millennial length stable isotope tree ring series for climate reconstruction. Global and Planetary Change, 2012, 82~83 :96~103 |
| 71 | Hangartner S, Kress A, Saurer M et al. Methods to merge overlapping tree-ring isotope series to generate multi-centennial chronologies. Chemical Geology, 2012, 294~295 :127~134 |
| 72 | Wang Wenzhi, Liu Xiaohong, Shao Xuemei et al. A 200-yr temperature record from tree-ring δ13C at the Qaidam Basin of the Tibetan Plateau after identifying the optimum method to correct for changing atmospheric CO2 and δ 13 C. Journal of Geophysical Research, 2011,116 (G4):G04022 |
| 73 | Xu Guobao, Chen Tuo, Liu Xiaohong et al. Potential linkages between the moisture variability in the northeastern Qaidam Basin, China, since 1800 and the East Asian summer monsoon, as reflected by tree-ring δ18O. Journal of Geophysical Research, 2011, 116 :D09111 |
| 74 | Wang Wenzhi, Liu Xiaohong, Xu Guobao et al. Moisture variations over the past millennium characterized by Qaidam Basin tree-ring δ18O. Chinese Science Bulletin, 2013,58 (32):3956~3961 |
| 75 | An Wenling, Liu Xiaohong, Leavitt S W et al. Specific climatic signals recorded in earlywood and latewood δ18O of tree rings in Southwestern China. TellusB, 2012, 64 :18703 |
| 76 | Kern Z, Patkó M, Kzmér M et al. Multiple tree-ring proxies(earlywood width, latewood width and δ 13 C)from pedunculate oak(Quercus robur L.), Hungary. Quaternary International, 2013, 293 :257~267 |
| 77 | Loader N J, Santillo P M, Woodman-Ralph J P et al. Multiple stable isotopes from oak trees in Southwestern Scotland and the potential for stable isotope dendroclimatology in maritime climatic regions. Chemical Geology, 2008,252 (1~2):62~71 |
| 78 | Young G H F, McCarroll D, Loader N J et al. A 500-year record of summer near-ground solar radiation from tree-ring stable carbon isotopes. The Holocene, 2010,20 (3):315~324 |
| 79 | Loader N J, Young G H F, Grudd H et al. Stable carbon isotopes from Torneträsk,Northern Sweden provide a millennial length reconstruction of summer sunshine and its relationship to Arctic circulation. Quaternary Science Reviews, 2013, 62 :97~113 |
| 80 | McCarroll D, Loader N J, Jalkanen R et al. A 1200-year multiproxy record of tree growth and summer temperature at the northern pine forest limit of Europe. The Holocene, 2013,23 (4):471~484 |
| 81 | Liu Yu, Wu Xiangding, Leavitt S W et al. Stable carbon isotope in tree rings from Huangling, China and climatic variation. Science in China, 1996,39 (2):152~161 |
| 82 | 徐国保. 北疆地区树轮稳定氧同位素比率(δ18O)气候意义. 北京: 中国科学院大学博士学位论文, 2014. 83~92 Xu Guobao. Climatic Significance of Stable Oxygen(δ18O)in Tree-ring in North Part of Xinjiang Uygur Autonomous Region. Beijing:The Doctoral Dissertation of University of Chinese Academy of Sciences, 2014. 83~92 |
| 83 | Liu Xiaohong, Xu Guobao, Grieinger J et al. A shift in cloud cover over the southeastern Tibetan Plateau since 1600:Evidence from regional tree-ring δ18O and its linkages to tropical oceans. Quaternary Science Reviews, 2014, 88 :55~68 |
| 84 | Aucour A M, Tao F X, Sheppard S M F et al. Climatic and monsoon isotopic signals(δD, δ 13 C)of Northeastern China tree rings. Journal of Geophysical Research, 2002,107 (D9):4074 |
| 85 | Xu Chenxi, Zheng Huaizhou, Nakatsuka T et al. Oxygen isotope signatures preserved in tree ring cellulose as a proxy for April-September precipitation in Fujian, the subtropical region of Southeast China. Journal of Geophysical Research, 2013,118 (23):2013JD019803 |
| 86 | Leavitt S W. Regional expression of the 1988 U.S. midwest drought in seasonal δ 13 C of tree rings. Journal of Geophysical Research, 2007,112 (D6):D06107 |
| 87 | Leavitt S W, Woodhouse C A, Castro C L et al. The North American monsoon in the U. S. Southwest:Potential for investigation with tree-ring carbon isotopes. Quaternary International, 2011,235 (1~2):101~107 |
| 88 | Treydte K, Schleser G H, Schweingruber F H et al. The climatic significance of δ 13 C in subalpine spruces(Ltschental, Swiss Alps). TellusB, 2001,53 (5):593~611 |
| 89 | Fichtler E, Helle G, Worbes M. Stable-carbon isotope time series from tropical tree rings indicate a precipitation signal. Tree-Ring Research, 2010,66 (1):35~49 |
| 90 | 刘晓宏, 秦大河, 邵雪梅等. 西藏喜马拉雅冷杉年轮δ 13 C与气候意义. 科学通报, 2003,48 (11):1209~1213 Liu Xiaohong, Qin Dahe, Shao Xuemei et al. Climatic significance of stable carbon isotope in tree rings of Abies spectabibis in southeastern Tibet. Chinese Science Bulletin, 2003,48 (11):1209~1213 |
| 91 | 刘晓宏, 秦大河, 邵雪梅等. 西藏林芝冷杉树轮稳定碳同位素对气候的响应. 冰川冻土, 2002,24 (5):574~578 Liu Xiaohong, Qin Dahe, Shao Xuemei et al. Stable carbon isotope of Abies spectabibis from Nyingchi County of Tibet Autonomous Region and its response to climate change. Journal of Glaciology and Geocryology, 2002,24 (5):574~578 |
| 92 | Liu Xiaohong, Shao Xuemei, Zhao Liangju et al. Dendroclimatic temperature record derived from tree-ring width and stable carbon isotope chronologies in the middle Qilian Mountains, China. Arctic, Antarctic, and Alpine Research, 2007,39 (4):651~657 |
| 93 | Liu Xiaohong, Shao Xuemei, Liang Eryuan et al. Species-dependent responses of juniper and spruce to increasing CO2 concentration and to climate in semi-arid and arid areas of Northwestern China. Plant Ecology, 2007,193 (2):195~209 |
| 94 | Treydte K, Frank D C, Saurer M et al. Impact of climate and CO2 on a millennium-long tree-ring carbon isotope record. Geochimica et Cosmochimica Acta, 2009,73 (16):4635~4647 |
| 95 | Dansgaard W. Stable isotopes in precipitation. Tellus, 1964,16 (4):436~468 |
| 96 | Burk R L, Stuiver M. Oxygen isotope ratios in trees reflect mean annual temperature and humidity. Science, 1981,211 (4489):1417~1419 |
| 97 | Liu Xiaohong, An Wenling, Treydte K et al. Tree-ring δ18O in Southwestern China linked to variations in regional cloud cover and tropical sea surface temperature. Chemical Geology, 2012, 291 :104~115 |
| 98 | Liu Xiaohong, Zeng Xiaomin, Leavitt S W et al. A 400-year tree-ring δ18O chronology for the southeastern Tibetan Plateau:Implications for inferring variations of the regional hydroclimate. Global and Planetary Change, 2013, 104 :23~33 |
| 99 | Xu Guobao, Liu Xiaohong, Qin Dahe et al. Tree-ring δ18O evidence for the drought history of eastern Tianshan Mountains, Northwest China since 1700 AD. International Journal of Climatology, 2014,34 (12):3336~3347 |
| 100 | Treydte K, Frank D, Esper J et al. Signal strength and climate calibration of a European tree-ring isotope network. Geophysical Research Letters, 2007,34 (24):L24302 |
| 101 | Saurer M, Schweingruber F, Vaganov E A et al. Spatial and temporal oxygen isotope trends at the northern tree-line in Eurasia. Geophysical Research Letters, 2002,29 (15):10~11 |
| 102 | Grieinger J, Bräuning A, Helle G et al. Late Holocene Asian summer monsoon variability reflected by δ18O in tree-rings from Tibetan junipers. Geophysical Research Letters, 2011,38 (3):L03701 |
| 103 | Xu Hai, Hong Yetang, Hong Bin. Decreasing Asian summer monsoon intensity after 1860 AD in the global warming epoch. Climate Dynamics, 2012,39 (7~8):2079~2088 |
| 104 | Shi Chunming, Daux V, Zhang Qibing et al. Reconstruction of southeast Tibetan Plateau summer climate using tree ring δ18O:Moisture variability over the past two centuries. Climate of the Past, 2012,8 (1):205~213 |
| 105 | Xu Guobao, Liu Xiaohong, Qin Dahe et al. Drought history inferred from tree ring δ 13 C and δ18O in the central Tianshan Mountains of China and linkage with the North Atlantic Oscillation. Theoretical and Applied Climatology, 2014,116 (3~4):385~401 |
| 106 | Xu Guobao, Liu Xiaohong, Qin Dahe et al. Relative humidity reconstruction for northwestern China's Altay Mountains using tree-ring δ18O. Chinese Science Bulletin, 2014,59 (2):190~200 |
| 107 | Xu Chenxi, Sano M, Nakatsuka T. A 400-year record of hydroclimate variability and local ENSO history in northern Southeast Asia inferred from tree-ring δ18O. Palaeogeography, Palaeoclimatology, Palaeoecology, 2013, 386 :588~598 |
| 108 | Sano M, Tshering P, Komori J et al. May-September precipitation in the Bhutan Himalaya since 1743 as reconstructed from tree ring cellulose δ18O. Journal of Geophysical Research, 2013,118 (15):8399~8410 |
| 109 | Sano M, Ramesh R, Sheshshayee M et al. Increasing aridity over the past 223 years in the Nepal Himalaya inferred from a tree-ring δ18O chronology. The Holocene, 2011,22 (7):809~817 |
| 110 | Sano M, Xu C, Nakatsuka T. A 300-year Vietnam hydroclimate and ENSO variability record reconstructed from tree ring δ18O. Journal of Geophysical Research, 2012,117 (D12):D12115 |
| 111 | An Wenling, Liu Xiaohong, Leavitt S W et al. Relative humidity history on the Batang-Litang Plateau of Western China since 1755 reconstructed from tree-ring δ18O and δD. Climate Dynamics, 2014,42 (9~10):2639~2654 |
| 112 | Liu Weiguo, Feng Xiahong, Liu Yu et al.δ18O values of tree rings as a proxy of monsoon precipitation in arid Northwest China. Chemical Geology, 2004,206 (1~2):73~80 |
| 113 | Liu Yu, Cai Qiufang, Liu Weiguo et al. Monsoon precipitation variation recorded by tree-ring δ18O in arid Northwest China since AD 1878. Chemical Geology, 2008,252 (1~2):56~61 |
| 114 | Miller D L, Mora C I, Grissino-Mayer H D et al. Tree-ring isotope records of tropical cyclone activity.Proceedings of the National Academy of Sciences of the United States of America, 2006,103 (39):14294~14297 |
| 115 | Epstein S, Yapp C J. Climatic implications of the D/H ratio of hydrogen in CH groups in tree cellulose. Earth and Planetary Science Letters, 1976,30 (2):252~261 |
| 116 | Gori Y, Wehrens R, Greule M et al. Carbon, hydrogen and oxygen stable isotope ratios of whole wood, cellulose and lignin methoxyl groups of Picea abies as climate proxies. Rapid Communications in Mass Spectrometry, 2013,27 (1):265~275 |
| 117 | Yapp C J, Epstein S. Climatic significance of the hydrogen isotope ratios in tree cellulose. Nature, 1982,297 (5868):636~639 |
| 118 | Feng Xiahong, Epstein S. Climatic temperature records in δD data from tree rings. Geochimica et Cosmochimica Acta, 1995,59 (14):3029~3037 |
| 119 | Liu Xiaohong, An Wenling, Leavitt S W et al. Recent strengthening of correlations between tree-ring δ 13 C and δ18O in mesic Western China:Implications to climatic reconstruction and physiological responses. Global and Planetary Change, 2014, 113 :23~33 |
| 120 | Jahren A H, Sternberg L S L O. Annual patterns within tree rings of the Arctic middle Eocene(ca .45Ma):Isotopic signatures of precipitation, relative humidity, and deciduousness. Geology, 2008,36 (2):99~102 |
| 121 | Helle G, Schleser G H. Beyond CO2-fixation by Rubisco——An interpretation of 13 C/12 C variations in tree rings from novel intra-seasonal studies on broad-leaf trees. Plant, Cell and Environment, 2004,27 (3):367~380 |
| 122 | Kagawa A, Sugimoto A, Maximov T C. 13 CO2 pulse-labelling of photoassimilates reveals carbon allocation within and between tree rings. Plant, Cell and Environment, 2006,29 (8):1571~1584 |
| 123 | Kagawa A, Sugimoto A, Maximov T C. Seasonal course of translocation, storage and remobilization of 13 C pulse-labeled photoassimilate in naturally growing Larix gmelinii saplings. New Phytologist, 2006,171 (4):793~804 |
| 124 | Eglin T, Francois C, Michelot A et al. Linking intra-seasonal variations in climate and tree-ring δ 13 C:A functional modelling approach. Ecological Modelling, 2010,221 (15):1779~1797 |
| 125 | Leavitt S W. Prospects for reconstruction of seasonal environment from tree-ring δ 13 C:Baseline findings from the Great Lakes area, USA. Chemical Geology, 2002,192 (1~2):47~58 |
| 126 | Eilmann B, Buchmann N, Siegwolf R et al. Fast response of Scots pine to improved water availability reflected in tree-ring width and δ 13 C. Plant, Cell and Environment, 2010,33 (8):1351~1360 |
| 127 | Barbour M M, Walcroft A S, Farquhar G D. Seasonal variation in δ 13 C and δ18O of cellulose from growth rings of Pinus radiata. Plant, Cell and Environment, 2002,25 (11):1483~1499 |
| 128 | Livingston N J, Spittlehouse D L. Carbon isotope fractionation in tree ring early and late wood in relation to intra-growing season water balance. Plant, Cell and Environment, 1996,19 (6):768~774 |
| 129 | Schulze B, Wirth C, Linke P et al. Laser ablation-combustion-GC-IRMS—A new method for online analysis of intra-annual variation of δ 13 C in tree rings. Tree Physiology, 2004,24 (11):1193~1201 |
| 130 | Krepkowski J, Gebrekirstos A, Shibistova O et al. Stable carbon isotope labeling reveals different carry-over effects between functional types of tropical trees in an Ethiopian mountain forest. New Phytologist, 2013,199 (2):441~451 |
| 131 | Schubert B A, Jahren A H. Quantifying seasonal precipitation using high-resolution carbon isotope analyses in evergreen wood. Geochimica et Cosmochimica Acta, 2011,75 (22):7291~7303 |
| 132 | Sheu D D, Kou P, Chiu C H et al. Variability of tree-ring δ 13 C in Taiwan fir:Growth effect and response to May-October temperatures. Geochimica et Cosmochimica Acta, 1996,60 (1):171~177 |
| 133 | Klein T, Hemming D, Lin T et al. Association between tree-ring and needle δ 13 C and leaf gas exchange in Pinus halepensis under semi-arid conditions. Oecologia, 2005,144 (1):45~54 |
| 134 | Managave S R. Model evaluation of the coherence of a common source water oxygen isotopic signal recorded by tree-ring cellulose and speleothem calcite. Geochemistry, Geophysics, Geosystems, 2014,15 (4):905~922 |
| 135 | Zhu Mengfang, Stott L, Buckley B et al. Indo-Pacific Warm Pool convection and ENSO since 1867 derived from Cambodian pine tree cellulose oxygen isotopes. Journal of Geophysical Research, 2012, 117 :D11307 |
| 136 | Zhu Mengfang, Stott L, Buckley B et al.20th century seasonal moisture balance in Southeast Asian montane forests from tree celluloseδ18O. Climatic Change, 2012,115 (3~4):505~517 |
| 137 | Xu Chenxi, Sano M, Yoshimura K E I et al. Oxygen isotopes as a valuable tool for measuring annual growth in tropical trees that lack distinct annual rings. Geochemical Journal, 2014,48 (4):371~378 |
| 138 | Managave S R, Ramesh R. Isotope dendroclimatology: A review with a special emphasis on tropics handbook of environmental isotope geochemistry. In: Baskaran M ed. Handbook of Environmental Isotope Geochemistry, Advances in Isotope Geochemistry. Berlin Heidelberg:Springer, 2011. 811~833 |
| 139 | Managave S R, Sheshshayee M S, Borgaonkar H P et al. Past break-monsoon conditions detectable by high resolution intra-annual δ18O analysis of teak rings. Geophysical Research Letters, 2010,37 (5):L05702 |
| 140 | Managave S, Sheshshayee M, Bhattacharyya A et al. Intra-annual variations of teak cellulose δ18O in Kerala, India:Implications to the reconstruction of past summer and winter monsoon rains. Climate Dynamics, 2010,37 (3~4):555~567 |
| 141 | Anchukaitis K J, Evans M N. Tropical cloud forest climate variability and the demise of the Monteverde golden toad.Proceedings of the National Academy of Sciences of the United States of America, 2010,107 (11):5036~5040 |
| 142 | Johnstone J A, Roden J S, Dawson T E. Oxygen and carbon stable isotopes in coast redwood tree rings respond to spring and summer climate signals. Journal of Geophysical Research, 2013,118 (4):1438~1450 |
| 143 | Berkelhammer M, Stott L D. Modeled and observed intra-ring δ18O cycles within Late Holocene Bristlecone pine tree samples. Chemical Geology, 2009,264 (1~4):13~23 |
| 144 | Poussart P F, Schrag D P. Seasonally resolved stable isotope chronologies from Northern Thailand deciduous trees. Earth and Planetary Science Letters, 2005,235 (3):752~765 |
| 145 | Poussart P F, Evans M N, Schrag D P. Resolving seasonality in tropical trees:Multi-decade, high-resolution oxygen and carbon isotope records from Indonesia and Thailand. Earth and Planetary Science Letters, 2004,218 (3~4):301~316A |
| 146 | Sarris D, Siegwolf R, Körner C. Inter-and intra-annual stable carbon and oxygen isotope signals in response to drought in Mediterranean pines. Agricultural and Forest Meteorology, 2013,168:59~68 |
| 147 | Li Zhenghua, Labbé N, Driese S G et al. Micro-scale analysis of tree-ring δ18O and δ13C on α-cellulose spline reveals high-resolution intra-annual climate variability and tropical cyclone activity. Chemical Geology, 2011,284(1~2):138~147 |
| 148 | Roden J S, Johnstone J A, Dawson T E. Intra-annual variation in the stable oxygen and carbon isotope ratios of cellulose in tree rings of coast redwood(Sequoia sempervirens). The Holocene, 2009,19 (2):189~197 |
| 149 | Evans M N. Toward forward modeling for paleoclimatic proxy signal calibration:A case study with oxygen isotopic composition of tropical woods. Geochemistry, Geophysics, Geosystems, 2007,8 (7):Q07008 |
| 150 | Managave S R, Sheshshayee M S, Ramesh R et al. Response of cellulose oxygen isotope values of teak trees in differing monsoon environments to monsoon rainfall. Dendrochronologia, 2011,29 (2):89~97 |
| 151 | Schollaen K, Heinrich I, Neuwirth B et al. Multiple tree-ring chronologies(ring width, δ13C and δ18O)reveal dry and rainy season signals of rainfall in Indonesia. Quaternary Science Reviews, 2013,73:170~181 |
| 152 | Huang J-G, Bergeron Y, Denneler B et al. Response of forest trees to increased atmospheric CO2. Critical Reviews in Plant Sciences, 2007,26 (5~6):265~283 |
| 153 | IPCC. Climate change 2013:The physical science basis. In:Stocker T F, Qin D, Plattner G-K et al.eds. Contribution of Working Group Ⅰ to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, United Kingdom and New York:Cambridge University Press, 2013.1~1535 |
| 154 | Morison J. Response of plants to CO2 under water limited conditions. Vegetatio, 1993,104/105:193~209 |
| 155 | Morgan J, Pataki D, Körner C et al. Water relations in grassland and desert ecosystems exposed to elevated atmospheric CO2. Oecologia, 2004,140 (1):11~25 |
| 156 | Peñuelas J, Canadell J G, Ogaya R. Increased water-use efficiency during the 20th century did not translate into enhanced tree growth. Global Ecology and Biogeography, 2011,20 (4):597~608 |
| 157 | Peñuelas J, Hunt J M, Ogaya R et al. Twentieth century changes of tree-ring δ13C at the southern range-edge of Fagus sylvatica: Increasing water-use efficiency does not avoid the growth decline induced by warming at low altitudes. Global Change Biology, 2008,14 (5):1076~1088 |
| 158 | Fischlin A, Midgley G F, Price J T R et al. Ecosystems, their properties, goods, and services. In:Parry M L, Canziani O F, Palutikof J P et al. eds. Climate Change 2007:Impacts, Adaptation and Vulnerability. Contribution of Working Group Ⅱ to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge:Cambridge University Press, 2007. 211~272 |
| 159 | Linares J C, Camarero J J. From pattern to process: Linking intrinsic water-use efficiency to drought-induced forest decline. Global Change Biology, 2012,18 (3):1000~1015 |
| 160 | Linares J C, Delgado-Huertas A, Camarero J J et al. Competition and drought limit the response of water-use efficiency to rising atmospheric carbon dioxide in the Mediterranean fir Abies pinsapo. Oecologia, 2009,161 (3):611~624 |
| 161 | Andreu-Hayles L, Planells O, Guti rrez E et al. Long tree-ring chronologies reveal 20th century increases in water-use efficiency but no enhancement of tree growth at five Iberian pine forests. Global Change Biology, 2011,17 (6):2095~2112 |
| 162 | Silva L C R, Horwath W R. Explaining global increases in water use efficiency:Why have we overestimated responses to rising atmospheric CO2 in natural forest ecosystems?PloS One, 2013,8 (1):e53089 |
| 163 | Cole C T, Anderson J E, Lindroth R L et al. Rising concentrations of atmospheric CO2 have increased growth in natural stands of quaking aspen(Populus tremuloides). Global Change Biology, 2010,16 (8):2186~2197 |
| 164 | Mcmahon S M, Parker G G, Miller D R. Evidence for a recent increase in forest growth. Proceedings of the National Academy of Sciences of the United States of America, 2010,107 (8):3611~3615 |
| 165 | Silva L C, Anand M. Probing for the influence of atmospheric CO2 and climate change on forest ecosystems across biomes. Global Ecology and Biogeography, 2013,22 (1):83~92 |
| 166 | Nock C A, Baker P J, Wanek W et al. Long-term increases in intrinsic water-use efficiency do not lead to increased stem growth in a tropical monsoon forest in Western Thailand. Global Change Biology, 2011,17 (2):1049~1063 |
| 167 | Granda E, Rossatto D, Camarero J J et al. Growth and carbon isotopes of Mediterranean trees reveal contrasting responses to increased carbon dioxide and drought. Oecologia, 2014,174 (1):307~317 |
| 168 | Saurer M, Siegwolf R T W, Schweingruber F H. Carbon isotope discrimination indicates improving water-use efficiency of trees in Northern Eurasia over the last 100 years. Global Change Biology, 2004,10 (12):2109~2120 |
| 169 | Scheidegger Y, Saurer M, Bahn M et al. Linking stable oxygen and carbon isotopes with stomatal conductance and photosynthetic capacity:A conceptual model. Oecologia, 2000,125 (3):350~357 |
| 170 | Gómez-Guerrero A, Silva L C R, Barrera-Reyes M et al. Growth decline and divergent tree ring isotopic composition(δ13C and δ18O)contradict predictions of CO2 stimulation in high altitudinal forests. Global Change Biology, 2013,19 (6):1748~1758 |
| 171 | Sitch S, Huntingford C, Gedney N et al. Evaluation of the terrestrial carbon cycle, future plant geography and climate-carbon cycle feedbacks using five Dynamic Global Vegetation Models(DGVMs). Global Change Biology, 2008,14 (9):2015~2039 |
| 172 | Friedlingstein P, Joel G, Field C B et al. Toward an allocation scheme for global terrestrial carbon models. Global Change Biology, 1999,5 (7):755~770 |
| 173 | Wang Wenzhi, Liu Xiaohong, An Wenling et al. Increased intrinsic water-use efficiency during a period with persistent decreased tree radial growth in Northwestern China:Causes and implications. Forest Ecology and Management, 2012,275:14~22 |
| 174 | Leuzinger S, Koerner C. Water savings in mature deciduous forest trees under elevated CO2. Global Change Biology, 2007,13 (12):2498~2508 |
| 175 | Fisher J B, Badgley G, Blyth E. Global nutrient limitation in terrestrial vegetation. Global Biogeochemical Cycles, 2012,26 (3), doi:10.1029/2011GB004252 |
| 176 | Wu Chaoyang, Hember R A, Chen J M et al. Accelerating forest growth enhancement due to climate and atmospheric changes in British Colombia, Canada over 1956~2001. Scientific Reports, 2014,4:4461 |
| 177 | Gradowski T, Thomas S C. Phosphorus limitation of sugar maple growth in central Ontario. Forest Ecology and Management, 2006,226 (1~3):104~109 |
| 178 | Marshall J D, Brooks J R, Lajtha K. Sources of variation in the stable isotopic composition of plants. Stable Isotopes in Ecology and Environmental Science, 2007,2:22~60" |
 , Xu Guobao①②③, Wang Wenzhi①③, Wu Guoju①③, Zeng Xiaomin④, An Wenling①③, Zhang Xuanwen①③
, Xu Guobao①②③, Wang Wenzhi①③, Wu Guoju①③, Zeng Xiaomin④, An Wenling①③, Zhang Xuanwen①③
Abstract
Stable isotope ratios (carbon, hydrogen and oxygen) in tree rings are closely related to the climate and environment conditions of trees growing period. Controlled experiments suggested that stable isotopes ratios recorded the information of water source, drought, intrinsic water-use efficiency and others. Tree-ring stable isotopes have become one of the important proxies in paleoclimate and global change ecology investigations. Tree-ring stable isotopes could retain low-frequency climate change signals, and it can be used to reconstruct paleo-climate and to study in term of global change in ecology by combing with tree-ring width proxy. Herein, we reviewed the theory of the tree-ring stable isotope fractionation, and assessed the results of the paleoclimate and environment using the tree-ring stable isotopes. We paid more attention on the progresses on the tree-ring stable isotopes over the different climatic regimes of China. We concluded that the tree-ring carbon stable isotope is a good proxy for the temperature variability, and the tree-ring oxygen stable isotope is a good indicator for the moisture condition in arid and semi-arid China. Lastly, we suggested the future developing terms of the tree-ring stable isotopes investigations in China.