 2015, Vol.35
2015, Vol.35



1 引言
祁连圆柏(Sabina przewalskii Kom.)是我国特有的常绿针叶树种,主要分布在青海、 甘肃河西走廊及其南部和四川北部等地区。在祁连山区,祁连圆柏是主要建群树种之一,生长在海拔2800~4300m的阳坡,因其生长年限长,对气候变化响应敏感而被广泛应用于树轮气候研究,对过去气候变化分析具有十分重要的意义,取得了显著的科研成果[1, 2, 3, 4],而且祁连圆柏是国内目前可以用来建立千年年表的唯一树种,运用祁连圆柏年表重建了过去千年尺度的降水、 温度、 干旱指数等[5, 6, 7, 8]。然而这些树轮气候重建工作是建立在树木年轮指标与气象数据之间的统计学基础上的,对树轮气候响应的生理机制并不太清楚,这有时会造成人们对相关分析结果的困惑。
树木径向生长研究对理解树轮-气候响应的生理机制具有重要意义[9]。虽然祁连山地区基于祁连圆柏的树轮气候研究工作已经不少,但对于生长季树木生长动态及其与环境间的关系等方面研究比较缺乏,只有Wang等[10, 11]对祁连山中部祁连圆柏做了一些研究。祁连山东西长800多公里,不同区域生态环境和气候状况均有一定差异,而目前对祁连山东部的研究基本没有,因此该地区祁连圆柏年内生长动态监测十分必要。
树木径向生长测量仪(Dendrometer)可以记录高分辨率的树木茎干变化动态,结合气象站数据,可以对树木年轮形成与环境因子间的关系进行评估,被国内外学者广泛用于树木径向生长监测研究[11, 12, 13, 14, 15, 16, 17]。本研究利用树木径向生长测量仪对祁连山东部3棵祁连圆柏生长季进行连续监测,研究了祁连圆柏生长季径向变化动态,并结合观测点自动气象站数据,探讨环境因子与树木径向增长之间的关系。
2 材料和方法 2.1 研究区概况研究区(36°40′~36°44′N,102°36′~102°45′E; 海拔1998~3165m)位于甘肃省永登县连城自然保护区吐鲁沟国家森林公园内( 图 1),地处祁连山东部,属大通河流域,深居内陆,远离海洋,气候类型为温带大陆性气候。区内日照比较丰富,全年日照时数为2655.2小时,年太阳总辐射为469kJ/cm2,年平均气温7.4℃,≥0℃的活动积温为3204.9℃,无霜期125~135天,年蒸发量1542mm,年降水量419mm,主要集中于6~9月,占全年降水量的60 % [18]。从吐鲁沟沟口(海拔2000m)到山顶(海拔3500m)为逐渐加宽的沟谷型地貌,沟内植被资源丰富且表现出明显的垂直分带特征。分布有祁连圆柏(Sabina przewalskii)、 青海云杉(Picea crassifolia)、 青杄(Picea wilsorii)和油松(Pinus tabulaeformis)等针叶树种和山杨(Populus davidiana)、 白桦(Betula platyphylla)和红桦(Betula albo-sinensis)等阔叶树种; 另外,灌木以及草本植物也比较丰富。

|
图 1 监测点位置 Fig. 1 Location of the monitoring site |
本研究中祁连圆柏监测点位于吐鲁沟圈湾(36°43′21″N, 103°37′59″E; 海拔3100m),坡度 20°~30°,郁闭度0.2,为祁连圆柏纯林,零星伴有金露梅(Potentilla fruticosa)和鲜黄小檗(Berberis diaphana)等灌木以及禾本科和莎草科为主的草本植物。
在2013年用点式树木径向生长测量仪(Point Dendrometer,Ecomatic Germany; 型号:DR; 精度=±2μm)对生长健康、 长势良好的3棵祁连圆柏进行生长季径向生长监测。3棵树分别用YB1、 YB2和YB3表示,具体信息见 表 1。测量仪安装在树木胸径处(1.3m),为了减少树皮的影响,提前剥掉了表层死皮。数据记录时段为2013年4月17日到9月6日,完全包含了祁连圆柏生长季,中间没有数据缺失和异常,数据记录时间间隔为30分钟,一天获取48个数据并储存在数据采集器中。将每天48个数据求平均得到原始数据日均值序列并用于生长模拟,对3棵圆柏原始数据求平均值,然后用于提取日径向增长量(SRI)。
| 表 1 3棵样木基本信息 Table 1 General information of the 3 sample trees |
距离样木约50m处建有自动气象站(Oneset HOBO U30),记录空气温度(T,℃)、 相对湿度(RH,% )、 土壤温度(ST,℃)、 土壤含水量(WC,m3/m3)、 降水量(P,mm)、 太阳辐射(W/m2)、 风速(m/s)、 风向等气象因子。数据每10分钟记录一次,储存在HOBO数据采集器中。根据自动气象站2011~2013年的记录( 图 2),观测点平均最低气温为-18.7℃,平均最高气温20.3℃,年降水量651.6mm,主要集中于7~9月,占全年降水量的59 % ,而5~6月占29 % 。

|
图 2 监测点2013年和2011~2013年温度与降水情况 Fig. 2 Monthly temperature and precipitation recorded by automatic meteorological station in monitoring site in 2013 and 2011~2013 respectively |
通过测量仪数据来确定生长季树木生长开始、 结束和主要生长时期等树木生理信息对生长-气候相关分析十分重要,然而由于以形成层开始活动为标志的生长开始容易和春季复水相混淆[19, 20],只能通过测量仪数据来确定高海拔地区树木生长主要时期[14]。相比其他“S”型函数,Gompertz函数更适合树木生长季径向生长的模拟,也为众多研究者广泛使用[13, 16, 21]。本研究用Gompertz生长模型对祁连圆柏样木原始数据日均值序列和3个样木平均原始数据日均值序列进行模拟,模拟时段为5~9月。然后得到生长速率曲线,认为生长速率大于仪器测量精度(4μm)的时段为祁连圆柏主要生长时期并用于生长-气候相关分析。
模拟公式如下:
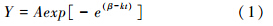
公式(1)中,Y为径向增长原始数据日均值; A为上渐近线; β为与Y初始值有关的参数; k表示内秉生长率,t表示时间。
2.2.4 日径向增长量(SRI)提取树干径向变化由两部分构成,一是由树干液流昼夜变化引起的收缩和膨胀[20, 22, 23],为可逆变化,一是由细胞分裂和增大等引起的不可逆的径向生长[24]。日径向生长量(SRI)可以用最大值法、 平均值法和周期法等多种方法获得[12, 25, 26, 27],本文根据Deslauriers等[13]的周期法提取SRI。将树干径向日变化周期分为先后3个阶段( 图 3):1)收缩阶段,树干直径从早晨最大值收缩到当天最小值的时间区间; 2)膨胀阶段,树干直径由最小值膨胀到第二天早晨最大值的时间区间; 3)径向增长阶段,从树干恢复到当天早晨最大值时起,到第二天早晨最大值的时间区间,是膨胀阶段的一部分。一个完整日周期包括收缩阶段和膨胀阶段。SRI是通过计算连续两周期膨胀阶段最大值的差值得到的,当后一个周期膨胀阶段最大值大于前一个周期的最大值时( 图 3-A),SRI记为正值,若差值为负值( 图 3-C),则认为树木无径向生长,SRI记为零。处理数据为3颗圆柏原始数据平均值,且只对主要生长时期提取SRI( 图 4)。

|
图 3 树干径向日周期变化 A——正常周期持续大约24小时(regular cycle lasting about 24h),B——阴雨天的长周期(long cycle in rainy days),C——后一个周期 没有超过前一周期最大值(previous cycle maximum is not reached) Fig. 3 The stem's circadian cycle divided into three distinct phases |

|
图 4 3个样木平均累积径向变化曲线及SRI变化曲线两条虚线表示 图 5D中的主要生长时期 Fig. 4 Mean time series of cumulative stem radial variation for 3 trees and its extracted daily SRI variation. The two dotted lines represent the main growing period showed in Fig. 5D |
为研究树干日径向生长量(SRI)与不同阶段气象条件的关系, 根据上述提取SRI的时间节点将气象数据也分成对应的3个阶段, 然后与各个阶段降水量(P,mm)、 平均气温(Tmean,℃)、 最高气温(Tmax,℃)、 最低气温(Tmin,℃)、 土壤温度(ST,℃)、 土壤含水量(WC, m3/m3)、 相对湿度(RH,%)和水汽压亏缺(VPD, kPa)等做Pearson相关分析。VPD是由空气湿度和温度计算得到, 此外, 为了确定不同阶段气象要素对树木径向生长的贡献, 利用逐步回归分析建立SRI与不同阶段气象条件之间的多元线性模型, 逐步回归时以F值的概率(probability of F)决定变量的进入(entry)和剔除(removal), 进入值和剔除值分别设置为0.05和0.10。
3 结果 3.1 树干日变化如图 3-A所示, 在晴朗天气下, 树干径向变化表现出规律的昼夜周期波动, 一般从上午07:00~08:00开始, 树木因蒸腾作用和光合活动耗水, 树干径向收缩, 下午16:00~17:00收缩到最小, 然后开始吸水膨胀, 在晚上01:00左右恢复到白天收缩前状态, 到第二天上午07:00~08:00膨胀到最大值, 然后进入下一个周期, 持续时间基本在 24±3个小时之间。而在阴雨天气(图 3-B), 树干持续吸水膨胀, 会出现长时间的持续增长, 周期明显增长, 径向增长量(SRI)也比较大, 天气转晴后又快速下降并恢复正常的日周期波动。
3.2 径向生长模拟用Gompertz模型分别对3棵祁连圆柏径向生长状况和平均生长状况进行模拟并据此得到生长速率曲线(图 5)。模拟结果显示拟合度(R2)均超过96%, 达到了0.01的显著性水平, 说明Gompertz模型能够很好地模拟祁连圆柏生长季径向生长。从图 5可以看出, 3个样木的累积生长曲线和生长速率曲线均有差异, 在最终累积生长量和最大生长速率上, 都是YB2>YB3>YB1, 但最大生长速率出现在5月底到6月初, 3个样木生长速率都在5月初超过了4μm/天, 但再次降低到4μm/天的日期并不相同, YB1和YB2为7月10日前后降到4μm/天, 而YB3在6月底就达到此速率, 这些差异可能是树木个体生理活动不同造成的。根据图 5D, 3个样木平均累积生长量为962μm, 平均最大生长速率为21.8μm/天, 大于速率4μm/天的时期为5月4日到7月8日。

|
图 5 祁连圆柏累积径向变化Gompertz函数模拟和径向生长速率A、 B、 C和D分别表示YB1、 YB2、 YB3以及3个样木平均值的模拟; 虚线表示生长速率为4μm/天 Fig. 5 Gompertz function modeled curves of cumulative radial variation and radial growth rate. A, B, C, and D represent the simulation of YB1, YB2, YB3 and mean value of 3 trees respectively. The dashed line represent the daily growth rate equal to 4μm/d |
日径向增长量(SRI)与各阶段气象参数间的相关分析(n=46)显示(图6), 日径向增长量与收缩阶段所有气象因子无显著相关关系; 与增长阶段、 膨胀阶段和整个周期的一些气象因子具有显著相关关系, 具体是: 与降水和相对湿度呈极显著正相关, 而与最低气温和水汽压亏缺都显著负相关, 与膨胀阶段和整个周期平均气温在0.05水平上负相关, 与最高温和增长阶段平均气温不相关。

|
图 6 日径向增长量(SRI)与不同阶段气象因子相关关系虚线和实线分别表示显著性水平p<0.05和p<0.01 Fig. 6 Pearson correlation coefficients between daily SRI and meteorological variables for each phase of the circadian cycle. Significant level(p<0.01 and p<0.05)is showed with lines |
利用逐步回归分析建立多元线性模型, 如下:


其中, 公式(2)是SRI与整个周期(阶段4)气象条件间的回归模型, 公式(3)表示SRI与膨胀阶段气象条件间的回归模型, 公式(4)表示SRI与膨胀阶段和增长阶段气象条件之间的多元回归模型, 下标中的2、 3和4分别表示膨胀阶段、 增长阶段和完整周期。回归方程中所有参数均通过0.05显著性水平检验, 回归方程相关统计信息见表 2。
| 表 2 回归方程统计信息 Table 2 Statistical information of regression equations |
Dendrometer监测显示研究区祁连圆柏径向增长主要时期为5月初到7月上旬,7月中旬以后树干径向累积增长几乎停止。我们在同一监测点运用微树芯法对祁连圆柏径向生长形态学分析[28]发现5月到7月中旬径向快速生长,且累积生长量超过全年80 % ,但形成层活动直到8月底才结束。而在本研究中,7月中旬以后树干径向累积增长几乎停止,这可能是因为7月中旬以后主要形成尺寸较小的晚材细胞,由细胞增大引起的日径向增长很不明显,容易被茎干水分变化导致的昼夜周期变化掩盖,进而导致累积增长量不再增大,因此本研究确定的主要是树木年轮早材部分的形成时期,与形态学监测的结果一致[28]。Deslauriers等[14]也发现对于高海拔地区,通过Dendrometer数据只能确定树木主要生长时期,且主要生长时期与早材细胞形成时期基本一致。
速率曲线( 图 5)显示祁连圆柏最大生长速率出现在5月底到6月初。我们通过微树芯法发现最大生长速率出现在6月上旬[28],比本研究中Dendrometer确定的时间晚了大约一周,这可能是因为5月底树干直径因温度升高和降雨增多而快速膨胀,从而导致通过记录仪数据得到的生长峰值提前。李兴欢等[29]在利用Dendrometer对小兴安岭红松的研究中也发现最大生长速率期提前这一现象; 而Wang等[11]通过对祁连山中部不同海拔祁连圆柏研究发现最大值都出现在6月,但具体日期仍因不同海拔和不同生长季而有较大差异。
4.2 径向生长与气候的关系相关分析表明日径向增长量(SRI)更多的受到膨胀阶段和增长阶段气象条件的影响,而收缩阶段的气象条件与SRI的相关关系不显著,并且去除收缩阶段气象要素后,多元回归方程的相关系数显著提高。说明下午16:00~17:00到第二天上午07:00~08:00气象条件对径向生长影响很大。Deslauriers等[13]通过相关分析和响应函数分析也发现膨胀阶段和增长阶段环境因子对加拿大冷杉(Abies balsamea)径向生长具有十分重要的意义。在国内,李兴欢等[29]在研究了小兴安岭红松后发现,当去掉收缩阶段气象因子后,回归模型的解释量并没有显著降低,说明径向增长受收缩阶段气象条件影响可能较小。生长季树木年轮生长包括形成层细胞分裂、 细胞增大和细胞壁增厚及木质化3个方面[30],细胞增大是导致每天径向增长的主要因素。Dünisch和Bauch[31]用细胞分析法和Dendrometer两种方法对挪威云衫树苗进行生长监测,发现81 % 的细胞增大发生在下午18:00到第二天早晨06 :00,与水分比较充足的时间段相对应。
祁连圆柏日径向增长量(SRI)与膨胀阶段、 增长阶段的降水和相对湿度显著正相关,说明本地区祁连圆柏生长受到水分胁迫。国内外大量基于Dendrometer的研究[11, 19, 25, 32]发现水分条件对树木年内径向生长具有积极影响,祁连山地区圆柏树轮气候响应分析也表明5~6月水分对树木生长具有重要作用[8, 33, 34]; 另外,与水汽压亏缺(VPD)呈显著负相关更进一步说明水分对祁连圆柏径向生长的重要影响,在生理上,较高的VPD主要会间接影响细胞膨压,进而抑制细胞增大和径向生长[35]。膨胀阶段和增长阶段平均气温与最低气温与SRI显著负相关,这可能是因为较高夜间温度影响树干内水分平衡,进而不利于径向膨胀。Deslauriers等[36]也发现增长阶段平均气温与日径向增长之间存在负相关关系,而且这种负相关是通过温度对增长相持续时间长短的影响来实现的,而不是直接作用于代谢等生理活动。Tardif等[19]认为气温与径向增长间的负相关可能与温度导致的蒸腾作用不利于径向膨胀有关。
5 结论利用树木径向生长测量仪(Dendrometer)对祁连山东部吐鲁沟国家森林公园内3棵祁连圆柏生长季进行生长监测。Gompertz模型能够很好地模拟祁连圆柏生长季径向生长,径向增长主要时期为5月初到7月上旬,最大生长速率出现在5月底到6月初。日径向增长量(SRI)与收缩阶段气象因子没有明显相关,而与膨胀阶段和增长阶段气象因子显著相关,表明下午16:00~17:00到第二天上午07:00~08:00环境条件对径向生长具有重要意义。SRI与降水、 相对湿度和水汽压亏缺的相关结果表明祁连圆柏径向生长受到水分限制,另外,SRI与最低气温和平均气温显著负相关。多元回归模型表明,相比日气象数据,膨胀阶段和增长阶段气象因子能解释更多的径向生长变化。本研究的不足之处是只利用一个生长季监测数据进行分析,无法对分析结果进行进一步验证,另外,作为监测树木径向生长动态的间接手段,树木径向变化测量仪(Dendrometer)还存在不少局限,需要结合微树芯法或针刺创伤法等直接手段进行更深入的研究。
致谢 感谢审稿专家以及编辑部杨美芳老师在文章修改和完善等方面提出的建设性意见和建议。
| 1 | Gou X, Gao L, Deng Y et al. An 850-year tree-ring-based reconstruction of drought history in the western Qilian Mountains of Northwestern China. International Journal of Climatology, 2014, doi:10.1002/joc.4208 |
| 2 | Liang E, Shao X, Liu X. Annual precipitation variation inferred from tree rings since A.D. 1770 for the western Qilian Mts., northern Tibetan Plateau. Tree-Ring Research, 2009,65 (2):95~103 |
| 3 | 杨 保. 树轮记录的小冰期以来青藏高原气候变化的时空特征. 第四纪研究, 2012,32 (1):81~94 Yang Bao. Spatial and temporal patterns of climate variations over the Tibetan Plateau during the period 1300~2010. Quaternary Sciences, 2012,32 (1):81~94 |
| 4 | Gao L, Gou X, Deng Y et al. Climate-growth analysis of Qilian juniper across an altitudinal gradient in the central Qilian Mountains, Northwest China. Trees-Structure and Function, 2012,27 (2):379~388 |
| 5 | Yang B, Qin C, Wang J L et al. A 3500-year tree-ring record of annual precipitation on the northeastern Tibetan Plateau. Proceedings of the National Academy of Sciences of the United States of America, 2014,111 (8):2903~2908 |
| 6 | Shao X M, Xu Y, Yin Z Y et al. Climatic implications of a 3585-year tree-ring width chronology from the northeastern Qinghai-Tibetan Plateau. Quaternary Science Reviews, 2010,29 (17~18):2111~2122 |
| 7 | 刘洪滨, 邵雪梅, 黄 磊等. 青海省海西州德令哈地区近千年来年降水量变化特征分析. 第四纪研究, 2005,25 (2):116~183 Liu Hongbin, Shao Xuemei, Huang Lei et al. Statistical characteristics of annual precipitation variation during the past 1000 years in Delingha, Qinghai Province, China. Quaternary Sciences, 2005,25 (2):116~183 |
| 8 | Gou X, Deng Y, Gao L et al. Millennium tree-ring reconstruction of drought variability in the eastern Qilian Mountains, Northwest China. Climate Dynamics, 2014, doi:10.1007/s00382~014-2431-y |
| 9 | Gričar J, Rathgeber C B K, Fonti P. Monitoring seasonal dynamics of wood formation. Dendrochronologia, 2011,29 (3):123~125 |
| 10 | Wang Z, Yang B, Deslauriers A et al. Two phases of seasonal stem radius variations of Sabina przewalskii Kom. in Northwestern China inferred from sub-diurnal shrinkage and expansion patterns. Trees-Structure and Function, 2012,26 (6):1747~1757 |
| 11 | Wang Z, Yang B, Deslauriers A et al. Intra-annual stem radial increment response of Qilian Juniper to temperature and precipitation along an altitudinal gradient in Northwestern China. Trees-Structure and Function, 2015,29 (1):25~34 |
| 12 | Downes G, Beadle C, Worledge D. Daily stem growth patterns in irrigated Eucalyptus globulus and E.nitens in relation to climate. Trees-Structure and Function, 1999,14 (2):102~111 |
| 13 | Deslauriers A, Morin H, Urbinati C et al. Daily weather response of balsam fir(Abies balsamea(L.)Mill.)stem radius increment from dendrometer analysis in the boreal forests of Québec(Canada). Trees-Structure and Function, 2003,17 (6):477~484 |
| 14 | Deslauriers A, Rossi S, Anfodillo T. Dendrometer and intra-annual tree growth:What kind of information can be inferred?. Dendrochronologia, 2007,25 (2):113~124 |
| 15 | Drew D M, Downes G M, Grzeskowiak V et al. Differences in daily stem size variation and growth in two hybrid eucalypt clones. Trees-Structure and Function, 2009,23 (3):585~595 |
| 16 | Duchesne L, Houle D, D'Orangeville L. Influence of climate on seasonal patterns of stem increment of balsam fir in a boreal forest of Québec, Canada. Agrcultural and Forest Meteorology, 2012, 162~163 :108~114 |
| 17 | Krepkowski J, Bräuning A, Gebrekirstos A et al. Cambial growth dynamics and climatic control of different tree life forms in tropical mountain forest in Ethiopia. Trees-Structure and Function, 2010,25 (1):59~70 |
| 18 | 张军周, 勾晓华, 赵志千等. 树轮生态学研究中微树芯石蜡切片制作的方法探讨. 植物生态学报, 2013,37 (10):972~977 Zhang Junzhou, Gou Xiaohua, Zhao Zhiqian et al. Improved method of obtaining micro-core paraffin sections in dendroecological research. Chinese Journal of Plant Ecology, 2013,37 (10):972~977 |
| 19 | Tardif J, Flannigan M, Bergeron Y. An analysis of the daily radial activity of 7 boreal tree species, northwestern Quebec. Environmental Monitoring and Assessment, 2001,67 (1~2):141~160 |
| 20 | Kozlowski T T, Winget C H. Diurnal and seasonal variation in radii of tree stems. Ecology,1964, 45 :149~155 |
| 21 | Camarero J J, Guerrero-Campo J, Gutierrez E. Tree-ring growth and structure of Pinus uncinata and Pinus sylvestris in the Central Spanish Pyrenees. Arctic and Alpine Research, 1998,30 (1):1~10 |
| 22 | Offenthaler I, Hietz P, Richter H. Wood diameter indicates diurnal and long-term patterns of xylem water potential in Norway spruce. Trees-Structure and Function, 2001,15 (4):215~221 |
| 23 | Zweifel R, Item H, Häsler R. Stem radius changes and their relation to stored water in stems of young Norway spruce trees. Trees-Structure and Function, 2000,15 (1):50~57 |
| 24 | Irvine J, Grace J. Continuous measurements of water tensions in the xylem of trees based on the elastic properties of wood. Planta, 1997,202 (4):455~461 |
| 25 | Bouriaud O, Leban J M, Bert D et al. Intra-annual variations in climate influence growth and wood density of Norway spruce. Tree Physiology, 2005,25 (6):651~660 |
| 26 | Rossi S, Deslauriers A. Intra-annual time scales in tree rings. Dendrochronologia, 2007,25 (2):75~77 |
| 27 | Bräuning A, Volland-Voigt F, Burchardt I et al. Climatic control of radial growth of Cedrela Montana in a humid mountain rainforest in Southern Ecuador. Erdkunde, 2009,63 (4):337~345 |
| 28 | Gou X, Zhou F, Zhang Y et al. Forward modeling analysis of regional scale tree-ring patterns around the northeastern Tibetan Plateau, Northwest China. Biogeosciences, 2013,10 (6):9969~9988 |
| 29 | 李兴欢, 刘瑞鹏, 毛子军等. 小兴安岭红松日径向变化及其对气象因子的响应. 生态学报, 2014,34 (7):1635~1644 Li Xinghuan, Liu Ruipeng, Mao Zijun et al. Daily stem radial variation of Pinus koraiensis and its response to meteorological parameters in Xiaoxing'an Mountain. Acta Ecologica Sinica, 2014,34 (7):1635~1644 |
| 30 | Deslauriers A, Rossi S, Anfodillo T et al. Cambial phenology, wood formation and temperature thresholds in two contrasting years at high altitude in Southern Italy. Tree Physiology, 2008,28 (6):863~871 |
| 31 | Dünisch O, Bauch J. Influence of soil substrate and drought on wood formation of spruce(Picea-abies Karst)under controlled conditions. Holzforschung, 1994,48 (6):447~457 |
| 32 | Zweifel R, Zimmermann L, Newbery D M. Modeling tree water deficit from microclimate:An approach to quantifying drought stress. Tree Physiology, 2005,25 (2):147~156 |
| 33 | Yang B, Qin C, Bräuning A et al. Rainfall history for the Hexi Corridor in the arid Northwest China during the past 620 years derived from tree rings. International Journal of Climatology, 2011,,31 (8):1166~1176 |
| 34 | Liu Y, Sun J Y, Song H M et al. Tree-ring hydrologic reconstructions for the Heihe River watershed, Western China since AD 1430. Water Research, 2010,44 (9):2781~2792 |
| 35 | Major J E, Johnsen K H. Shoot water relations of mature black spruce families displaying a genotypex environment interaction in growth rate. Ⅲ. Diurnal patterns as influenced by vapor pressure deficit and internal water status. Tree Physiology, 2001,21 (9):579~587 |
| 36 | Deslauriers A, Anfodillo T, Rossi S et al. Using simple causal modeling to understand how water and temperature affect daily stem radial variation in trees. Tree Physiology, 2007,27 (8):1125~1136 |
Abstract
Intra-annual stem radial growth monitoring is important to understand the physiological mechanisms of dendroclimatic research. In this study, the radial increment of Qilian Juniper(Sabina przewalskii Kom.)in the eastern Qilian Mountains was monitored in 2013 using automatic point dendrometer to investigate tree-ring formation and its environmental influence. Three well-growing adult trees in the monitoring site(36°43'21"N, 103°37'59"E, with altitude of 3100m)were continuously monitored during the whole season in 2013. All dendrometers were mounted at about 1.3m height of trees and could get a record every 30min. Three continuous raw data series were obtained and no measuring mistake was found. Gompertz function was used to simulate the cumulative stem radial growth, and the homologous growth rate curves were gotten by the first derivative processing. The period that growth rate exceeds the recording accuracy of instrument(4μm/d)was defined as the main growing period and was used to SRI extraction and subsequent analyses. The entire circadian cycle was divided into three distinct phases:(1)Contraction phase, the period from the morning peak to afternoon trough. (2)Expansion phase, the period from afternoon trough to the next morning peak. (3)Stem radial increment phase(SRI), part of expansion phase that from the time when the stem radius exceeds the morning maximum to the subsequent maximum. The relationships between SRI and meteorological variables of different phases were assessed using Pearson correlation and Stepwise regression analysis. Meteorological parameters include precipitation(P, mm), air temperature(T, ℃), relative humidity(RH,%), soil temperature(ST, ℃), water content(WC, m3/m3), and vapor pressure deficit(VPD, kPa). Dendrometer records showed that the maximum of expansion phase appeared at 07 :00~08 :00 and minimum of contraction phase at 16 :00~17 :00. Gompertz function simulated well and the goodness of fit was higher than 96%.It was found that the main growth period, from early May to middle July, corresponded mainly with the growth of early wood. The maximum growth rate occurred in late May-early June. Correlation analyses showed that it is environmental factors of expansion and increment phases, rather than contraction phase, that correlated well with SRI, revealing the great influence of meteorological conditions from 16 :00~17 :00 to 07 :00~08 :00 on stem radial growth. SRI was positively correlated with precipitation and relative humidity and negatively with VPD, indicating the negative effect of drought on tree growth. Negative correlation results were found between SRI and mean air temperature and minimum air temperature. Multiple regression modeling showed that weather conditions of expansion and increment phase, compared with that of entire cycle, could explain more variation of SRI with a high explanation up to 87.1%.