 2015, Vol.35
2015, Vol.35



1 引言
树木年轮作为气候记录的代用指标,具有定年准确、 连续性强、 分辨率高、 易于获取和复本量大等优点[1],被广泛用于气候学研究以及古气候重建[2, 3, 4, 5, 6]。水曲柳(Fraxinus mandshurica Rupr.)是东北典型阔叶红松林内最主要阔叶树种,因年轮清晰且与气候因子相关性好,近年来已被用于树轮气候学研究[7, 8, 9, 10, 11]。已有学者利用水曲柳轮宽年表分别重建了长白山地区1765年以来的秋季(9~10)平均温度[9]和年降水变化[11],用水曲柳与红松混合年表重建了长白山敦化地区1854年以来的生长季(4~9)低温变化[8]。
小兴安岭是东北典型阔叶红松(Pinus koraiensis Sieb. et Zucc.)林的主要分布区。该区地处我国高纬度区域,是森林受气候变化影响最显著的区域之一[12]。了解该区过去气候变化历史及变异规律,对阔叶红松林顶极群落保护与恢复,以及探讨气候变化下现存阔叶红松林的稳定性和适应性、 碳源汇变化等都具有重要意义。但已有气候器测记录时间较短,因此获取长时间的气候代理数据成为目前亟待解决的问题。尹红等[10]虽然已利用小兴安岭五营地区红松树轮宽度重建了1796年以来10月温度变化,但仅有该气候重建远远不能满足当今的科研需求,还需要更多地点、 更多月份及更多气候指标的重建,来彼此验证及获得区域气候记录。
本文利用小兴安岭地区伊春市凉水国家级自然保护区内水曲柳树木年轮宽度资料,重建了1790年以来2~3月的低温变化,为伊春乃至小兴安岭地区近几百年来气候变化研究提供高分辨率的资料,同时也可为该地区的树木年轮气候学研究奠定基础。
2 材料与方法 2.1 研究地区概况本研究采样点(47.19°N,128.87°E; 海拔475m)位于凉水国家级自然保护区内(图 1),行政区划隶属于黑龙江省伊春市带岭区。地理上属我国小兴安岭的东南段,为典型的低山丘陵地貌,海拔280~707m,森林覆被率为96%,是我国现有保存下来的较大片原始阔叶红松林之一。地带性土壤为暗棕壤。

|
图 1 采样点和气象站点位置图 Fig. 1 Locating of sampling sites and meteorological station |
该区位于欧亚大陆的东侧,深受海洋气候的影响,具有明显的温带大陆性季风气候特征。冬季气候严寒、 干燥而多风雪。夏季降水集中、 气温较高。春秋降水量小,易发生干旱。伊春气象站(47.73°N,128.92°E; 海拔241m)气候资料表明,该区年平均气温-0.3℃,极端最高温度38.2℃,极端最低温度-43.1℃,1月平均最低温度-29.1℃,7月平均最高温度26.8℃; 年降水量676mm,主要集中在6~9月(占76%),雨热同季(图 2)。年平均相对湿度69.58%,年蒸发量805.4mm; 年日照时数2353小时,无霜期100~120天(http://www.lsbhq. com/ls-jj.asp?ClassID=30)。

|
图 2 伊春气象站温度和降水量的月变化(1957~2009年) Fig. 2 Monthly mean,minimum and maximum temperature and total precipitation of Yichun Meteorological Station(1957~2009) |
研究区植被类型为典型阔叶红松林。林内有水曲柳、 臭冷杉(Abies nephrolepis(Trautv.)Maxim.)、 鱼鳞云杉(Picea jezoensis var. microsperma (Lindl.)Cheng et L. K.Fu)、 紫椴(Tilia amurensis Rupr.)、 枫桦(Betula costata Trautv.)、 大青杨(Populus ussuriensis Kom.)、 青楷槭(Acer tegmentosum Maxim.)和色木槭(A. mono Maxim.)等针、 阔叶乔木; 灌木主要有刺五加(Acanthopanax senticosus (Rupr. et Maxim.)Harms)、 毛榛子(Corylus mandshurica Maxim.)、 东北山梅花(Philadelphus schrenkii Rupr.)和暴马丁香(Syringa reticulata(Blume)H. Hara var. amurensis(Ruprecht)P. S.Green & M. C.Chang)等; 藤本植物有五味子(Schisandra chinensis (Turcz.)Baill.)和狗枣猕猴桃(Actinidia kolomikta (Maxim. et Rupr.)Maxim.)等[13]。
2.2 研究方法 2.2.1 采样及预处理2011年6月,在凉水国家级自然保护区内选取树龄较大且生长正常的水曲柳,在胸高处(H=1.3m)进行生长锥取样。在每株树木相对方向上钻取2个样芯以保证交叉定年的准确性,然后将钻取的树芯装入塑料管并进行编号,同时记录样本的特征及周围的生境条件,共采集23棵树46个芯。将野外采集的所有样芯带回实验室,进行固定、 干燥、 打磨等预处理[14]。
2.2.2 交叉定年及年表建立首先,采用骨架图法对经预处理的样芯,在显微镜下交叉定年,接着用年轮分析仪进行宽度测量,精度为0.001mm。其次,用COFECHA程序[15]来检验定年结果的准确性,根据软件提示检查并修订交叉定年与宽度测量过程中出现的误差,直到能确保年轮的日历年代准确为止。最后,利用ARSTAN程序[16]来完成对最终年轮宽度序列的去趋势和标准化。去趋势方法采用保守的负指数或线性函数法,用双权重平均法进行年轮曲线的标准化,最终建立了水曲柳的标准年表(STD)、 差值年表(RES)和自回归年表(ARS)。
年表统计特征表明(表 1),水曲柳树轮序列最长为277年(1734~2010年),3种年表的平均敏感度均较低(0.11~0.13),说明树轮资料具有较弱的高频波动信号,这与其他学者的研究结果一致[7, 9, 11],标准年表的一阶自相关系数较高,说明前一年气候也可能对当年的树木生长产生影响[2]。此外,年表的信噪比、 第一主成分方差解释量以及树木间平均相关系数均较高,表明年轮样本间具有较好的一致性且包含了较多环境信息[1, 4]。以上年表统计分析表明水曲柳年表适合进行年轮-气候分析。综合年表特征,最后选择变异较小、 离散度较低、 逐年变化具有共性较高的自回归年表(图 3),用于随后的分析与重建。可靠年表序列长度(EPS>0.85)为221年,即1790~2010年。
| 表 1 凉水国家自然保护区内水曲柳年表统计特征 Table 1 Major statistic characteristics of chronologies of Fraxinus mandshurica at Liangshui National Nature Reserve |

|
图 3 凉水自然保护区水曲柳自回归年表 实线、 虚线和黑色粗线分别表示年轮指数、 复本量和11年滑动平均序列 Fig. 3 The tree-ring ARS chronologies of Fraxinus mandshurica at Liangshui National Natural Reserve. The solid,dashed and black bold line represents the tree-ring index,sample depth,and an 11-year moving average series,respectively |
用距离采样点最近的(61km)伊春气象站1957~2009年的气候数据进行相关性分析,气候数据由中国气象科学数据共享服务网(http://cdc.cma.gov.cn/)免费提供。用Mann-Kendall方法[17]对该站气候记录进行检验,气温和降水数据变化相对均一,可用于进一步分析。考虑到前一年气候状况对当年水曲柳径向生长的影响[1, 4],分析时选用前一年和当年月平均气温、 月最低气温、 月最高气温和月降水量数据与年轮指数进行相关分析。
2.2.4 统计分析用SPSS19.5软件相关分析来确定气候因子对水曲柳径向生长的影响。基于相关分析结果,同时结合水曲柳年表和气温序列对比变化趋势,确定对2~3月平均最低气温进行重建。通过SPSS软件中的线性回归来获得重建方程,用符号检验、 误差缩减值以及乘积平均值等来分段检验方程的稳定性[1, 18]; 此外,还采用自助法(Bootstrap)和刀切法(Jackknife)[19]来进一步检验重建方程的可靠性。为确定伊春过去221年的偏冷和偏暖期,本文将11年滑动平均温度变化超出均值0.5倍标准差的时段,定义为偏冷期或偏暖期。采用MTM程序对重建温度序列进行多带谱分析[20],以检验2~3月最低温度变化的周期性。
3 结果与讨论 3.1 径向生长与气候要素的关系水曲柳年轮指数与前一年和当年各气候因子相关分析结果表明(图 4),上一年和当年的温度与降水均对径向生长均有很大影响,且上一年的影响相对较大。水曲柳径向生长与前一年1~3月、 当年3月和6月的降水呈显著正相关(p<0.05); 与前一年1~6月、 10月、 12月和当年1~4月、 6月、 10月、 12月的月平均最低温度均呈显著正相关(p<0.05),与前一年2月相关系数最高(r=0.68)。年轮指数与月平均温度和月平均最高温度的相关性分析结果与月平均最低气温类似,均是前一年要高于当年。年轮指数与年平均温度偏相关(控制年降水)分析结果(r =0.68,p<0.01)也表明水曲柳与温度的相关性要明显高于降水,温度是限制该地区水曲柳径向生长的主要因子,这与其他学者在长白山的研究结果[8, 9]基本一致; 但不同的是李牧和王晓春[8]研究发现敦化地区水曲柳径向生长与生长季(4~9月)温度相关性较高,而王蔚蔚等[9]则发现长白山地区水曲柳径向生长主要受秋季(9~10月)温度限制。小兴安岭与长白山地区气候变化特征自身存在差异,可能是导致两地区水曲柳对月气候因子(温度)响应窗口存在不确定性的原因。 由南到北(长白山到小兴安岭)生长季、 秋季温度对水曲柳的限制作用逐渐减弱,而生长季前(2~4月)温度对水曲柳径向生长的限制作用逐渐增强[5],可能是越往北生长季水分限制作用逐渐加强的结果,温度升高所引起的“暖干旱”对径向生长的负效应远远大于温度升高本身所引起的径向生长的正效应。

|
图 4 水曲柳年轮指数与月降水、 均温(a)以及月平均最低、 最高温(b)的相关性分析 短划线表示显著相关(p=0.05),横坐标中的“P”代表前一年 Fig. 4 Pearson correlation coefficients between tree-ring index of Fraxinus mandshurica and monthly total precipitation, mean(a),minimum and maximum temperature(b). Significant correlations(p=0.05)are indicated by dash lines. The letter “P” in abscissa represents the previous year |
本研究发现小兴安岭地区水曲柳径向生长主要受冬半年(P10~3月)的温度限制,尤其是2~3月的最低气温。水曲柳为耐寒树种,但特别容易受到是霜冻灾害的影响,轻者使当年生长量降低,重者使整株林木死亡[21],故水曲柳的径向生长与月最低温度关系密切。尽管冬季温度对树木生长的正反馈效应及其反馈机制目前尚不完全清楚,但冬季(休眠期)温度与夏季(生长季)温度对于树木生长来说具有同等重要的作用在北半球高纬度寒冷地区已被证实[22]。虽然东北地区2~3月气温很低,树木处于休眠状态,但是植物体内(芽、 茎和根等部位)的温度要比气温高的多,树体内的生理活动并没有完全停止,树体内仍有树液流动,组织的代谢(如合成抗寒或叶/花芽分化的底物)仍在进行,树体仍然有分化、 发育过程。较高的冬季(2~3月)最低气温,一方面,促使植物体内的温度相对较高,这将大大提高树体内各种生物酶的活性,进而加快树体内各项物理、 生物及化学活动进程,为树体内各项生理、 生化活动提供环境条件; 另一方面,导致冰雪覆盖减少、 水土流失加剧、 土壤温度升高,进而加速生物地球化学循环和凋落物的分解,促进土壤养分的利用(矿化与释放)[23, 24, 25, 26],为树体内各项生理生化活动提供物质和能量基础。这都将为水曲柳日后生长打下坚实的基础,并且是水曲柳在生长季内形成宽轮的必备条件。此外,冬季(2~3月)最低气温可大大降低水曲柳地下根系、 地上休眠芽等器官以及其他组织或细胞发生冻害可能性,一定程度上确保了生长季开始后水曲柳具有足够的根叶量,以保证其正常的生长。暖冬往往会间接的影响夏季的温度,即暖冬后往往会紧跟着暖夏(某种意义上延长生长季长度),暖冬对夏季温度的间接影响也可能是冬季温度对水曲柳径向生长的正效应作用方式之一[22]。
若水曲柳积累了较多的营养物质(形成宽轮),在休眠期来临前往往会形成较多且饱满的休眠芽,为来年形成宽轮打下基础; 此外,植物根部及主干贮存的营养物质,也能为来年生长季初期的抽枝、 萌叶等生理过程提供较好物质及能量基础[27, 28]。因此,前一年积累的营养物质对展叶较晚、 落叶较早的水曲柳[29]来年的径向生长至关重要且贡献量较大; 反之,较低的2~3月最低温度,往往会造成当年和下一年形成窄轮。
3.2 2~3月最低温度重建年轮-气候关系分析表明,2~3月最低温度是伊春地区水曲柳径向生长的主要限制因素; 因此,选择伊春市气象站2~3月最低温度为重建因子,以树轮宽度自回归年表为变量,以1958~2010年为建模期,用一元线性回归模型建立树轮宽度指数与前一年2~3月平均最低温度之间的转换方程:
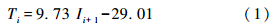
公式(1)中,Ti为第i年的2~3月平均最低温度,Ii+1为第i+1年水曲柳自回归年表的年轮指数。该重建方程的相关系数为0.71,方差解释量为51.0%,调整后方差解释量为50.1%,F检验值为53.2,远超过99.99%的显著水平,说明重建方程有较高的可靠性(图 5)。

|
图 5 年轮指数与伊春2~3月最低温度的 散点图(1958~2010年) Fig. 5 Scatter plot of February-March minimum temperature of Yichun and tree-ring indices with linear fitting curve(1958~2010) |
分别以1957~1982年为校准期(检验期)、 1983~2009年检验期(校准期),用分段法来检验重建方程的稳定性。独立统计检验的参数有方差解释量(R2)、 误差缩减值(RE)、 符号检验(ST)和乘积平均值检验(PMT)等几个统计量。检验结果表明(表 2),在两个独立检验期,除1957~1982年为校准期时的符号检验外,R2、 ST、 和PMT均达到了95%或99%的显著水平,表明重建方程稳定可靠。一般认为误差缩减值(RE)大于0时,重建方程具有有效性[1, 4, 18],本研究分段检验的RE值分别为0.74和0.77,均较好的通过了检验。
| 表 2 2~3月最低温度重建回归方程的分段校准和检验统计特征 Table 2 Calibration and verification status of the February-March minimum temperature regression equation for two split periods |
此外,本研究还采用了自助法和刀切法对重建方程进行了检验,进一步确保重建方程的可靠性和稳定性。检验结果表明(表 3),自助法和刀切法的多次统计结果各统计量的平均值均与原回归方程接近,说明重建结果较为理想,重建方程稳定可靠。综合以上检验结果,可知该重建方程稳定而可靠,可以用于重建伊春过去221年的2~3月份的平均最低温度。
利用上述方程重建了1790年来伊春的最低温度。重建值与观测值在共同区段(1957~2009年)总体上吻合较好,但重建值在变化趋势上小于实测值,重建值在高频变化上有一定损失(图 6)。一方面,超过一定阈值的温度变异,并不完全能够表现到年轮的宽窄上; 另一方面,部分温度信号可能被用于补偿其他因素(如降水、 光照、 二氧化碳浓度等)造成的年轮宽度非正常亏损,故重建结果的精确度降低。此外,气象站的气候资料在反映林区气候时具有一定的差异,影响树木年轮宽度序列与气候因子的相关系数,也是导致重建序列方差解释量减小,年轮宽度对极端气温响应偏低的原因。
| 表 3 2~3月最低温度重建回归方程的自助法和刀切法检验结果 Table 3 Statistical characteristics of the February-March minimum temperature regression equation based on Jackknife and Bootstrap test |

|
图 6 2~3月平均最低温度实测序列和重建序列对比 Fig. 6 Comparison between observed and reconstructed mean February-March minimum temperature |
用检验可靠的方程重建伊春过去221年(1790~2010年)2~3月最低温度序列。由重建序列可以看出(图 7),伊春过去221年中2~3月的最低温度一直处于不断变化之中,其特点是冷、 暖期交替出现,且冷、 暖期持续时间较长。例如1795~1819年的降温变化持续了25年,而1923~1955年的升温变化也持续了33年。重建序列1790年以前的温度可能受复本量的影响变化较为剧烈,伊春过去221年里存在7个偏冷期:1790~1791年、 1795~1819年、 1834~1841年、 1851~1854年、 1898~1899年、 1901~1907年和1959~1982年; 4个偏暖期:1822~1831年、 1872~1886年、 1923~1955年和1988~2010年,其他时段温度在均值附近波动,变化较小。重建序列中1785~1820年的低温期正好位于太阳活动的道尔顿极小期(Dalton Minimum,1790~1830年)[30],说明伊春地区的最低温度变化可能受太阳活动影响; 已有研究发现采样点附近红松树轮信号中也存在11年的太阳黑子活动周期,太阳活动可通过影响该区域的温度变化影响阔叶红松林动态[31]。重建序列中1960~1980年的低温时期在其他重建序列中均有发现(见图 7),表明该时段低温具有广泛的区域性和普遍性。此外,1980年以后显著升温(见图 7),伊春2~3月的低温一直处于均值以上,在李牧和王晓春[8]重建的敦化地区4~9月的最低温度以及Zhu等[32]重建长白山2~4月温度序列中均有发现,这也与第五次IPCC(Intergovernmental Panel on Climate Change)报告[33]的研究结果一致。

|
图 7 重建伊春2~3月最低温度(a)与敦化地区2~4月均温[32](b)、 敦化4~9月低温[8](c)以及哈尔滨2~3月实测低温(d)比较 红色波动线为年际变化,粗黑色曲线为11年滑动均值,青色和红色阴影区为一致的低温和高温时期 图 7a中蓝色与绿色曲线表示重建值的95%置信区间 1790年前的重建即使可靠性较低,但因其有一定的参考意义,故在图中给出的重建从1740年开始 Fig. 7 Comparison of reconstructed February-March minimum temperature (a) in Yichun,reconstructed February-April temperature[32] (b) and April-September temperature[8](c)in Dunhua and actual February-March minimum temperature (d) in Harbin. Red fluctuation lines represent means of reconstructed temperature series. Bold black curve is the 11-year moving average of each series. Cyan and red shaded areas represent low and high temperature zones with good agreements in these four temperature series,respectively. The blue and cyan curves showing in (a) represent the 95% confidence interval of predicted values |
伊春2~3月最低温度重建序列与Zhu等[32]重建的敦化2~4月均温(1750~2002年)、 李牧和王晓春[8]重建的敦化生长季(4~9月)最低温度(1854~2010年)及哈尔滨市2~3月最低温度实测值序列(1909~2010年)比较可以看出(见图 7),它们之间的变化趋势存在较好的一致性,但各序列并未完全对应,可能是由重建指标自身的差异所导致(最低温和均温)。重建的伊春过去2~3月最低温度与李牧和王晓春[8]重建的敦化地区4~9月的低温序列更为接近(即使其重建序列相对较短),与Zhu等[32]重建的敦化2~4月均温序列相似性略低,表明除研究区固有差异外,重建代理资料、 重建窗口及重建指标(如温度又可细分为均温、 最低温和最高温等)的差异均可能是各重建序列间存在差异的原因。此外,该重建序列与《中国气象灾害大典·黑龙江卷》[34]中记载的低温冷害、 霜冻灾害事件有着很强的一致性。例如清嘉庆五年(1800年)、 六年(1801年)、 八年(1803年)、 二十一年(1816年)、 二十二年(1817年),清道光十六至十八年(1836~1838年)、 二十一年(1841年)、 清光绪元年至五年(1875~1879年)、 十三年(1887年)、 二十七至二十八年(1901~1902年),清宣统元年至民国二年(1909~1913年)、 民国二十三至二十四年(1934~1935年)和民国二十八至三十二年(1939~1943年)……等时段记录低温冷害、 霜冻灾害事件[34]均在重建序列中被发现,进一步印证了本重建序列的准确性和可靠性。
多带谱分析结果表明(图 8),伊春2~3月最低温存在18.3年和4.9~4.8年的极显著周期(p<0.01),其中4.9~4.8年的周期与厄尔尼诺-南方涛动(El Nio-Southern Oscillation,简称ENSO)2~7年的准周期一致[35],表明伊春的最低温度可能受ENSO的影响; 大尺度气候波动虽然不会直接影响某一区域树木的生长,但能通过影响局地气候间接影响树木生长[36],该地区其他研究也发现树轮信号或利用树轮重建的气候信号中存在与ENSO对应的2~7年显著周期[6, 7, 8, 9, 31, 32, 37],进一步说明ENSO事件很大程度上影响该区的树木生长与气候(温度和降水)变化。18.3年的温度周期可能与太平洋年代际振荡(Pacific Decadal Oscillation,简称PDO)有关[38],PDO影响东北地区树木生长或温度变化已被其他研究所证实[39, 40]。此外,重建的温度序列中存在太阳活动(11.8~13.1a)和大西洋多年代际振荡(Atlantic Multi-decadal Oscillation,简称AMO)(41.0~44.4a、 51.3~60.2a和78.7a)的显著(p<0.05,图 8中未给出)周期,太阳活动与AMO已被证实在很大程度上影响着东北地区的树木生长与气候变化[31, 39, 40, 41]。

|
图 8 重建2~3月最低温度序列的功率谱分析 图中数字代表显著(p<0.01)的周期 Fig. 8 Multi-taper method spectrums analysis for the reconstructed mean February-March minimum temperature. Significance was tested at two levels(99% and 95%)against a red-noise background. Digital values on the plot are the significant(p<0.01)periods |
通过对小兴安岭伊春地区水曲柳的树轮宽度资料的分析,探讨树木径向生长对气候因子的响应,利用转换方程重建了伊春1790年以来2~3月的最低温度序列,并分析其变化特征。得出以下结论:
(1)制约小兴安岭伊春地区水曲柳径向生长最主要的气候因子是温度,尤其是前一年及当年冬季最低温度,与2~3月平均最低温度,均呈极显著正相关(p<0.01),降水影响相对较弱。休眠期(冬季)温度与生长季(早春、 夏秋)温度一样均对北半球高纬度地区落叶阔叶树水曲柳径向生长有明显的促进作用,冬季水曲柳树体内各种生理生化活动及外界复杂的生物地球化学循环过程可能对水曲柳生长季的径向生长至关重要。
(2)采用一元线性回归方程对伊春过去221年的2~3月平均最低温进行了重建,重建方程为:Ti=9.73 Ii+1-29.01,该方程的方差解释量为51.0%(R2adj=50.1%,F=53.2,p<0.0001),经分段、 自助法(Bootstrap)和刀切法(Jackknife)方法检验,重建结果稳定可靠。
(3)伊春过去221年存在7个偏冷期(1790~1791年、 1795~1819年、 1834~1841年、 1851~1854年、 1898~1899年、 1901~1907年和1959~1982年)和4个偏暖期(1822~1831年、 1872~1886年、 1923~1955年和1988~2010年)。最低温度变化存在18.3年和4.9~4.8年的极显著周期(p<0.01),温度周期性变化可能与太阳活动、 ENSO、 AMO和PDO有关。
| 1 | Fritts H. Tree Rings and Climate. London:Academic Press, 1976. 1~567 |
| 2 | Liang E Y, Dawadi B, Pederson N et al. Is the growth of birch at the upper timberline in the Himalayas limited by moisture or by temperature?. Ecology, 2014,95 (9):2453~2465 |
| 3 | Yang B, Qin C, Wang J L et al. A 3500-year tree-ring record of annual precipitation on the northeastern Tibetan Plateau. Proceedings of the National Academy of Sciences of the United States of America, 2014,111 (8):2903~2908 |
| 4 | 吴祥定. 树木年轮分析在环境变化研究中的应用. 第四纪研究, 1990,(2):188~196 Wu Xiangding. Application of tree ring analysis to the study on environment variation. Quaternary Sciences, 1990,(2):188~196 |
| 5 | 邵雪梅, 吴祥定. 利用树轮资料重建长白山过去气候变化. 第四纪研究, 1997,(1):76~85 Shao Xuemei, Wu Xiangding. Reconstruction of climate change on Changbai Mountain, Northeast China using tree ring data. Quaternary Sciences, 1997,(1):76~85 |
| 6 | 李宗善, 刘国华, 傅伯杰等. 利用树木年轮宽度资料重建川西米亚罗地区过去200年夏季温度的变化. 第四纪研究, 2011,31 (3):522~534 Li Zongshan, Liu Guohua, Fu Bojie et al. Tree ring-based summer temperature reconstruction over the past 200 years in Miyaluo of western Sichuan, China. Quaternary Sciences, 2011,31 (3):522~534 |
| 7 | 李 牧. 东北三大硬阔年轮与气候关系. 哈尔滨: 东北林业大学, 2013. 1~30 Li Mu. Climate-growth Relationships of Three Hardwood Species in the Northeast China. Harbin:Northeast Forestry University, 2013. 1~30 |
| 8 | 李 牧, 王晓春. 敦化三大硬阔、红松年轮-气候关系及生长季低温重建. 南京林业大学学报(自然科学版), 2013,37 (3):29~34 Li Mu, Wang Xiaochun. Climate-growth relationships of three hardwood species and Korean pine and minimum temperature reconstruction in growing season in Dunhua, China. Journal of Nanjing Forestry University(Natural Sciences Edition), 2013,37 (3):29~34 |
| 9 | 王蔚蔚, 张军辉, 戴冠华等. 利用树木年轮宽度资料重建长白山地区过去240年秋季气温的变化. 生态学杂志, 2012,31 (4):787~793 Wang Weiwei, Zhang Junhui, Dai Guanhua et al. Variation of autumn temperature over the past 240 years in Changbai Mountains of Northeast China:A reconstruction with tree-ring records. Chinese Journal of Ecology, 2012,31 (4):787~793 |
| 10 | 尹 红, 郭品文, 刘洪滨等. 利用树轮重建小兴安岭五营1796年以来的温度变化. 气候变化研究进展, 2009,5 (1):18~23 Yin Hong, Guo Pinwen, Liu Hongbin et al. Reconstruction of the October mean temperature since 1796 at Wuying from tree ring data. Advances in Climate Change Research, 2009,5 (1):18~23 |
| 11 | 张寒松, 韩士杰, 李玉文等. 利用树木年轮宽度资料重建长白山地区240年来降水量的变化. 生态学杂志, 2007,26 (12):1924~1929 Zhang Hansong, Han Shijie, Li Yuwen et al. Reconstruction of temporal variations of precipitation in Changbai Mountains area over past 240 years by using tree-ring width data. Chinese Journal of Ecology, 2007,26 (12):1924~1929 |
| 12 | Piao S L, Ciais P, Huang Y et al. The impacts of climate change on water resources and agriculture in China. Nature, 2010, 467 (7311):43~51 |
| 13 | 徐丽娜, 金光泽. 小兴安岭凉水典型阔叶红松林动态监测样地: 物种组成与群落结构. 生物多样性, 2012,20 (4):470~481 Xu Lina, Jin Guangze. Species composition and community structure of a typical mixed broad-leaved-Korean pine(Pinus koraiensis)forest plot in Liangshui Nature Reserve, Northeast China. Biodiversity Science, 2012,20 (4):470~481 |
| 14 | Stokes M A, Smiley T L. An Introduction to Tree-ring Dating. Chicago:University of Chicago Press, 1968. 1~73 |
| 15 | Holmes R L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bulletin, 1983,43 (1):69~78 |
| 16 | Cook E R, Holmes R L. Users Manual for Program ARSTAN. Tucson:Laboratory of Tree-Ring Research, University of Arizona, 1986. 1~9 |
| 17 | Kendall M G, Gibbons J D. Rank Correlation Methods. London:Edward Arnold, 1990. 1~272 |
| 18 | Cook E R, Meko D M, Stahle D W et al. Drought reconstructions for the continental United States. Journal of Climate, 1999,12 (4):1145~1162 |
| 19 | Efron B. Bootstrap methods: Another look at the jackknife. The Annals of Statistics, 1979,7 (1):1~26 |
| 20 | Mann M E, Lees J M. Robust estimation of background noise and signal detection in climatic time series. Climatic Change, 1996,33 (3):409~445 |
| 21 | 陈祥伟, 张 羽, 马洪波. 紫椴、水曲柳人工幼林霜冻发生与控制研究. 应用生态学报, 2000,11 (6):809~813 Chen Xiangwei, Zhang Yu, Ma Hongbo. Occurrence and control of frost in Tilia amurensis and Fraxinus mandshurica young plantations. Chinese Journal of Applied Ecology, 2000,11 (6):809~813 |
| 22 | Hollesen J, Buchwal A, Rachlewicz G et al. Winter warming as an important co-driver for Betula nana growth in western Greenland during the past century. Global Change Biology, 2015,21 (6):2410~2423 |
| 23 | Schimel J P, Bilbrough C, Welker J M. Increased snow depth affects microbial activity and nitrogen mineralization in two Arctic tundra communities. Soil Biology and Biochemistry, 2004,36 (2):217~227 |
| 24 | Inouye D W. Effects of climate change on phenology, frost damage, and floral abundance of montane wildflowers. Ecology, 2008,89 (2):353~362 |
| 25 | Morgner E, Elberling B, Strebel D et al. The importance of winter in annual ecosystem respiration in the High Arctic:Effects of snow depth in two vegetation types. Polar Research, 2010,29 (1):58~74 |
| 26 | Semenchuk P R, Elberling B, Amtorp C et al. Deeper snow alters soil nutrient availability and leaf nutrient status in High Arctic tundra. Biogeochemistry,2015, doi 10.1007/s10533~015-0082~7 |
| 27 | 李俊楠, 王文娜, 谢玲芝等. 去叶对水曲柳和落叶松苗木当年生长及细根动态的影响. 植物生态学报, 2014,38 (10):1082~1092 Li Junnan, Wang Wenna, Xie Lingzhi et al. Effects of defoliation on current-year stem growth and fine root dynamics in Fraxinus mandschuricaand Larix gmeliniiseedlings. Chinese Journal of Plant Ecology, 2014,38 (10):1082~1092 |
| 28 | 范志强, 王政权, 吴 楚等. 水曲柳苗木根系和叶片氮的分配及对生物量影响. 中国农学通报, 2008,24 (1):45~51 Fan Zhiqiang, Wang Zhengquan, Wu Chu et al. Nitrogen and biomass partitioning pattern in root and leaf of Fraxinus mandshurica seedlings. Chinese Agricultural Science Bulletin, 2008,24 (1):45~51 |
| 29 | 祝 宁, 江 洪, 金永岩. 中国东北天然次生林主要树种的物候研究. 植物生态学报, 1990,14 (4):336~349 Zhu Ning, Jiang Hong, Jin Yongyan. A phenology study on the common tree species of natural secondary forests in Northeast China. Chinese Journal of Plant Ecology, 1990,14 (4):336~349 |
| 30 | Eddy J A. The maunder minimum. Science, 1976, 192 (4245):1189~1202 |
| 31 | 朱良军, 金光泽, 王晓春. 典型阔叶红松林干扰历史重建及干扰形成机制. 植物生态学报, 2015,39 (2):125~139 Zhu Liangjun, Jin Guangze, Wang Xiaochun. Reconstruction of disturbance history of a typical broad-leaved Pinus koraiensis forest and mechanisms of disturbance occurrence. Chinese Journal of Plant Ecology, 2015,39 (2):125~139 |
| 32 | Zhu H F, Fang X Q, Shao X M et al. Tree ring-based February-April temperature reconstruction for Changbai Mountain in Northeast China and its implication for East Asian winter monsoon. Climate of the Past, 2009,5 (4):661~666 |
| 33 | Intergovernmental Panel on Climate Change(IPCC). Climate Change 2013:The Physical Science Basis. Contribution of Working Group Ⅰ to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge:Cambridge University Press, 2014. 1~254 |
| 34 | 孙永罡主编. 中国气象灾害大典: 黑龙江卷. 北京: 气象出版社, 2007. 7~32 Sun Yonggang ed. Grand Dictionary of China Meteorological Disasters:Volume Heilongjiang. Beijing:China Meteorological Press, 2007. 7~32 |
| 35 | McGregor H, Fischer M, Gagan M et al. A weak El Niño/Southern Oscillation with delayed seasonal growth around 4300 years ago. Nature Geoscience, 2013,6 (11):949~953 |
| 36 | 张先亮, 崔明星, 马艳军等. 大兴安岭库都尔地区兴安落叶松年轮宽度年表及其与气候变化的关系. 应用生态学报, 2010,21 (10):2501~2507 Zhang Xianliang, Cui Mingxing, Ma Yanjun et al. Larix gmelinii tree-ring width chronology and its responses to climate change in Kuduer, Great Xing'an Mountains. Chinese Journal of Applied Ecology, 2010,21 (10):2501~2507 |
| 37 | 徐 海, 洪业汤, 朱咏煊等. 安图红松树轮稳定δ 13 C, δ18O 序列记录的气候变化信息. 地质地球化学, 2002,30 (2):59~65 Xu Hai, Hong Yetang, Zhu Yongxuan et al. Information on climate change recorded in δ 13 C and δ18O series of Pinus koraiensis tree ring cellulose in Antu area. Geology-Geochemistry, 2002,30 (2):59~65 |
| 38 | Yasuda I. The 18.6-year period moon-tidal cycle in Pacific Decadal Oscillation reconstructed from tree-rings in western North America. Geophysical Research Letters, 2009,36 (5):L05605 |
| 39 | Chen Z J, He X Y, Chen W et al. Solar activity, global surface air temperature anomaly and Pacific Decadal Oscillation signals observed in urban outskirts tree ring records from Shenyang, China. Advances in Space Research, 2006,38 (10):2272~2284 |
| 40 | 姚启超, 王晓春. 小兴安岭不同海拔臭冷杉年轮-气候关系及大尺度气候影响. 北京林业大学学报(自然科学版), 2013,35 (2):30~38 Yao Qichao, Wang Xiaochun. Climate-growth relationships of Abies nephrolepis and its connection with large-scale climate change in Xiaoxing'an Mountains, Northeastern China. Journal of Beijing Forestry University(Natural Sciences Edition), 2013,35 (2):30~38 |
| 41 | Wang X C, Brown P M, Zhang Y N et al. Imprint of the Atlantic Multidecadal Oscillation on tree-ring widths in Northeastern Asia since 1568. PloS One, 2011,6 (7):e22740 |
Abstract
Xiaoxing'an Mountain, located in Northeast China, is the most typical distribution area of primary broadleaved-Korean pine forests, and is also one of the most significant areas affected by global climate change. To understand the past climate change and predict it in the future in this region, long-term and high-resolution climate records is critical and essential. Thus, long-term climate proxy is desperately needed due to shorter instrumental records. Here we present a 221-year February-March minimum temperature reconstruction based on tree-ring widths of Manchurian ash(Fraxinus mandshurica Rupr.)at Liangshui National Natural Reserve in Yichun, Northeast China. A total of 46 Manchurian ash cores from 23 living trees were extracted from this area(47.19°N, 128.87°E; 475m a.s.l.)in June, 2011. According to the standard methods developed by the International Tree-Ring Data Bank(ITRDB), these cores were pretreated, cross-dated, measured, detrended and standardized. Then, three chronologies(STD, RES and ARS)of Manchurian ash were developed. Correlation functions between tree-ring index and climatic variables indicate that winter temperature is essential for radial growth of deciduous Manchurian ash at high latitudes of Northern Hemisphere. Winter processes are as important as summer processes in controlling the Manchurian ash radial growth. According to the stronger relationship(r =0.71, p <0.01)between February-March minimum temperature in the previous year and tree-ring index(ARSTANT chronologies), we reconstruct the February-March minimum temperature in Yichun using a simple liner regression model. The reconstruction equation explained 51% of the variance of the minimum temperature(1958~2010). Seven cold periods(1790~17911795~18191834~18411851~18541898~18991901~1907 and 1959~1982)and four warm periods(1822~18311872~18861923~1955 and 1988~2010)occurred in this region for the past 221 years. The variations of February-March minimum temperature in Yichun exist 4.8~4.9a and 18.3a of significant(p <0.01)periods. Periodic changes in minimum temperature may be related to activities of the solar activity, El Nio -Southern Oscillation(ENSO), Atlantic Multi-decadal Oscillation(AMO)as well as the Pacific Decadal Oscillation(PDO).