 2022, Vol. 43
2022, Vol. 43文章信息
- 蒋露芳, 王莹莹, 彭慧, 李锐, 张芬, 王娜, 邵月琴, 姜庆五.
- Jiang Lufang, Wang Yingying, Peng Hui, Li Rui, Zhang Fen, Wang Na, Shao Yueqin, Jiang Qingwu
- 学龄儿童肥胖与肠道菌群多样性及菌属丰度的关联研究
- Association between obesity with the diversity and genus of gut microbiota in school-aged children
- 中华流行病学杂志, 2022, 43(2): 260-268
- Chinese Journal of Epidemiology, 2022, 43(2): 260-268
- http://dx.doi.org/10.3760/cma.j.cn112338-20210617-00478
-
文章历史
收稿日期: 2021-06-17




2. 复旦大学公共卫生学院公共卫生安全教育部重点实验室, 上海 200032;
3. 上海市嘉定区疾病预防控制中心, 上海 201800
2. Key Laboratory of Public Health Safety, Ministry of Education, School of Public Health, Fudan University, Shanghai 200032, China;
3. Jiading District Center for Disease Control and Prevention, Shanghai 201800, China
超重或肥胖是能量摄入和消耗之间的不平衡而导致的异常脂肪积累。根据《中国居民营养与慢性病状况报告(2020年)》的数据,2015-2019年我国成年人的超重率为34.3%,肥胖率为16.4%,6~17岁青少年的超重率为11.1%,肥胖率为7.9%。在过去30年中,我国青少年超重或肥胖呈急剧上升趋势,超重或肥胖的患病率从1992年的5.7%上升至2020年的19.0%[1]。儿童期肥胖不仅会增加成年后肥胖的风险,也与哮喘、MS、心血管疾病,甚至成年后过早死亡显著关联[2-3]。从饮食习惯上,我国居民对于动物性食物、深加工食物及含糖饮料等消费量逐年上升[4]。近年来,超重或肥胖与肠道菌群的关联性受到广泛关注[5]。肥胖影响肠道菌群的多样性和分布。与正常体重人群相比,肥胖人群肠道中厚壁菌门(Firmicutes)与拟杆菌门(Bacteroidetes)之比增加,肠道微生物多样性减少[6]。与正常体重小鼠相比,经动物油喂养后脂肪量增加的小鼠肠道菌群发生改变,毛螺菌科(Lachnospiraceae)和脱硫弧菌科(Desulfuribriaceae)的相对丰度增加[7]。在接受手术方案减重的肥胖患者中,与术前相比,术后恢复正常体重者多形类杆菌(B.thetaiotaomicron)和阿克曼菌(Akkermansia muciniphila)的相对丰度显著增加[8]。肠道菌群通过改变宿主营养吸收和能量代谢、促进脂肪的生成发挥作用[9]。曾有研究发现,尽管摄入等量的高脂食物,与肠道菌群多样性较高的肥胖者相比,多样性较低的肥胖者体重增加更为明显[10]。越来越多的研究揭示了肠道菌群的重要性,对于婴幼儿(< 3岁)和成年人(≥18岁)的肠道微生物群组成和功能目前已有较详细的描述,但对于儿童和青少年的研究相对缺乏[11]。本研究旨在分析学龄肥胖儿童和正常体重儿童肠道菌群的多样性差异,识别肥胖儿童肠道中的特征菌群,为后续相关的机制研究和学龄儿童肥胖的防治提供依据。
对象与方法1. 研究对象与样本量:
(1)样本量计算:微生物组研究的结局指标一般为微生物的多样性,以α多样性和β多样性作为因变量,计算样本量。采用Faith's PD描述α多样性差异,按照PD差异为2、统计学功效为80%来计算,所需总样本量约为110人;采用Unifrac距离法描述β多样性差异,按照显著性为5%、统计学功效为80%来计算,所需总样本量约为100人[12]。
(2)研究对象:选择上海市嘉定区某小学进行调查,该小学以本地生源为主,人群稳定。根据上述样本量计算结果,以及既往研究中上海市7岁儿童肥胖检出率(男生14.4%,女生11.2%)[13],若需纳入肥胖者55名(即总样本量的一半),则需一般人群382名(全为男生),或491名(全为女生)。2016年5月将该小学1~2年级所有儿童共461名纳入研究队列中,分别于2017年和2018年对其进行重复调查及采样。筛选出3次调查中均判定为肥胖的儿童,排除非酒精性脂肪肝等消化系统疾病、胰岛素抵抗等肥胖相关症状、近3个月有重大饮食调整或极端饮食习惯,以及近3个月长期服用抗生素或益生菌制剂者,最终将63名儿童纳入肥胖组,其中男生43名,女生20名。对照组在3次调查均判定为正常体重的儿童中进行选择,根据年龄(同周岁)、性别,在同班级中选择符合条件的儿童与肥胖组儿童进行1∶1匹配,若有多人均满足同一配对要求,则通过产生随机数在其中随机选择一位,最终共63名儿童纳入对照组中。收集两组儿童的相关样本,以便后续检测与分析。所有研究对象及其父母均签署知情同意书。
2. 研究内容与方法:
(1)问卷调查:采用自行设计的问卷,收集研究对象基本信息、饮食习惯、母乳喂养时间等情况,问卷由儿童主要监护人填写,于样本收集前一天统一发放,并于样本收集当天回收核验。
(2)体格检查与肥胖判断标准:使用同一型号的身高体重仪和软皮尺,按照儿少卫生学要求测量对象身高(cm)和体重(kg),并计算BMI(kg/m2)。学龄儿童肥胖根据《WS/T 586-2018学龄儿童青少年超重与肥胖筛查》[14]中年龄别、性别BMI界值进行判定。
(3)粪便样本收集与处理:现场调查前一日,由班主任将50 ml无菌采样杯(配有无菌采样棒)分发给学生,粪便的采样时间为7:00-8:00左右,要求尽量取粪便中段没有接触空气的部分≥10 g。若当日无法留有粪便,需按照相同取样要求次日或第三日收集粪便。将粪便采样杯尽快送至样本收集处,样本暂时储存在装有冰袋的保温箱里,而后运送至实验室进行分装,置于-80 ℃低温保存,供后续DNA抽提和测序使用。
(4)16S rDNA测序:利用正向引物347F(5'-CCT ACG GRR BGC ASC AGK VRV GAA T-3')和反向引物806R(5'-GGA CTA CNV GGG TWT CTA ATC C-3'),对肠道细菌16S rDNA的V3~V4区进行PCR扩增,通过琼脂糖凝胶电泳确定PCR产物的特异性并纯化PCR产物;利用Illumina Miseq M300测序仪(美国Illumina公司)进行双末端250 bp测序。测序由苏州金唯智生物科技有限公司完成。
3. 生物信息学分析:
(1)原始数据处理和可操作分类单元(OTU)聚类:使用VSEARCH软件,去除测序原始数据中低质量片段,将上述PCR产物进行拼接,去除重复序列、标签序列及引物序列等,进行过滤后得到优化序列。根据序列97%相似性,进行OTU聚类,统计每个样本中OTU的丰度信息,应用核糖体数据库项目(Ribosomal Database Project,RDP)分类器鉴定OTU代表性序列的微生物分类地位,使用RDP数据库进行物种注释。
(2)肠道菌群多样性指数计算:使用R 4.0.4软件分析样本中肠道菌群多样性情况,包括α多样性和β多样性。α多样性主要衡量个体中物种丰度和多样性,常用的度量指标包括Ace、Chao1、Shannon、Simpson 4种指数。其中Ace和Chao1指数侧重估算菌群中物种的丰度及数量,数值越大,物种多样性越高;Shannon和Simpson指数同时考虑物种的丰度和均匀度,Shannon指数越大、Simpson指数越小,物种多样性越高。β多样性主要描述样本之间的多样性,通常将其与降维方法结合使用,包括主坐标分析法(PCoA)、β多样性使用非加权Unifrac距离和加权Unifrac距离进行表示,非加权Unifrac距离仅考虑了物种有无,加权Unifrac距离同时考虑了物种有无和物种丰度。
4. 统计学分析:
(1)差异分析:符合正态分布的连续性资料用x±s表示,非正态分布的连续性资料用M(Q1,Q3)表示,分类资料用人数(构成比)表示。采用成组t检验、χ2检验等方法比较肥胖组和对照组儿童基本信息情况和α多样性指数的差异,使用相似性分析(ANOSIM)比较两组β多样性的差异。使用STAMP 2.1.3软件找出在两组共有菌属中差异有统计学意义的类别。
(2)关联分析:使用R 4.0.4软件进行数据分析与图形绘制,检验水准α=0.05。采用广义线性模型(GLM)分析肥胖与α多样性及显著差异菌属的关联。α多样性指数包括Ace、Chao1、Shannon、Simpson,均呈正态分布,连接函数选择“identity”;在肥胖组和对照组的共有菌属中,挑选在相对丰度上具有明显差异的菌属,菌属相对丰度呈Poisson分布,连接函数选择“log”。模型分别考虑粗模型(模型1)和混杂因素调整模型(模型2和模型3)。模型1中不调整任何变量,模型2中调整年龄、性别、SBP及饮食因素,模型3在模型2的基础上再调整分娩方式、母乳喂养时间以及近一周内使用抗生素情况。
结果1. 基本情况:肥胖组儿童的年龄、BMI、腰臀比、SBP、DBP均高于对照组儿童(P < 0.05)。除了食用零食频次(P=0.038)之外,两组儿童在其他食物食用频次上差异均无统计学意义(P > 0.05)。见表 1。
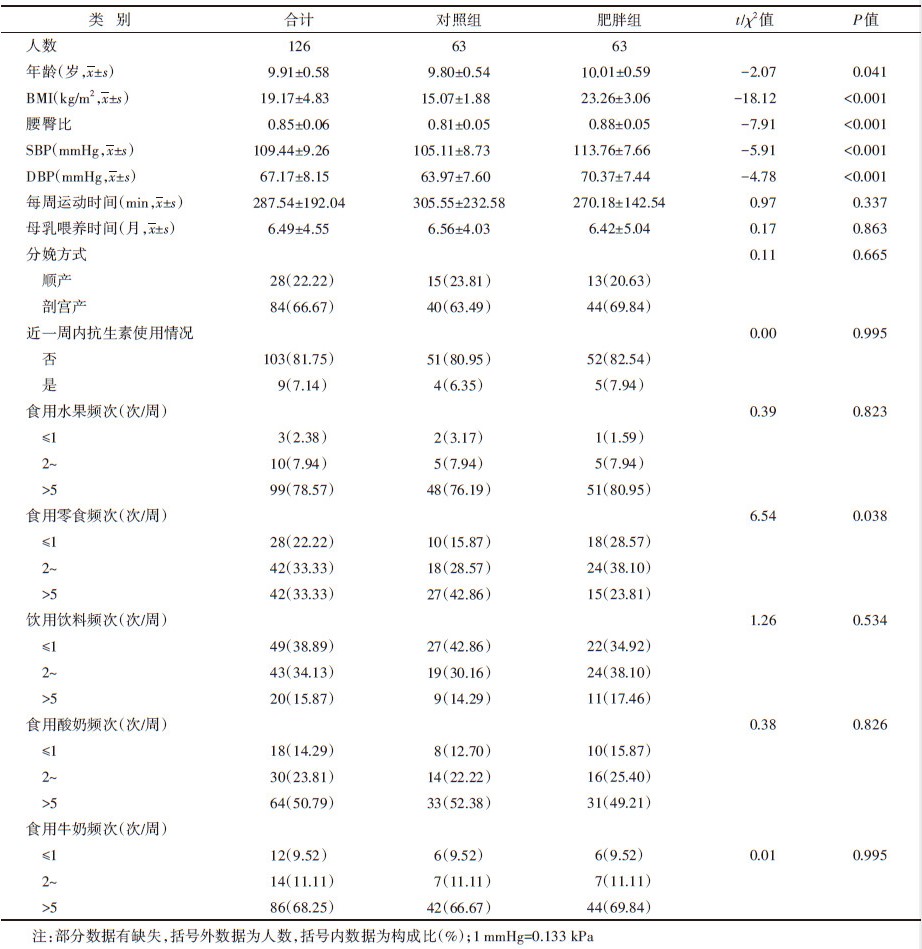
2. 研究对象肠道菌群的α多样性和β多样性:与对照组儿童相比,肥胖组儿童Ace和Chao1指数较低(P值分别为0.026和0.039),Shannon和Simpson指数差异均无统计学意义(P值分别为0.879和0.922)。见图 1。

|
| 注:Ace、Chao1、Shannon、Simpson为α多样性指数 图 1 对照组和肥胖组儿童肠道菌群的α多样性 |
对于β多样性的分析,基于非加权Unifrac距离和加权Unifrac距离的PCoA分析均无法将两组儿童明显区分,ANOSIM比较结果显示两组儿童肠道菌群差异有统计学意义(R > 0),但是组别贡献并不显著(非加权Unifrac距离:R=0.006,P=0.223;加权Unifrac距离:R=0.010,P=0.134)。见图 2。

|
| 注:横、纵坐标上的PC1、PC2、PC3分别为第1、2、3主成分,后面的数字表示在总变异中可解释的百分比 图 2 基于非加权Unifrac距离和加权Unifrac距离的主坐标分析 |
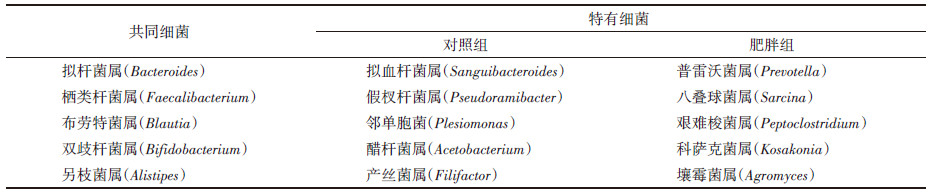
3. 研究对象肠道菌群的属级别差异:在两组儿童共有菌属中,相对丰度最高为拟杆菌属(Bacteroides);对于特有菌属,对照组中相对丰度最高为拟血杆菌属(Sanguibacteroides),而肥胖组中,相对丰度较高的主要为普雷沃菌属(Prevotella)和八叠球菌属(Sarcina)。见表 2。
STAMP分析结果显示,两组共有菌属中,共15个菌属的相对丰度在两组儿童中差异有统计学意义(P < 0.05)。肥胖组粪便中不动杆菌属(Agathobacter)相对丰度高于对照组最多,其次为瘤胃球菌属(Ruminococcus)、粪球菌属(Coprococcus_3)和柯林斯菌属(Collinsella),而阿克曼菌属(Akkermansia muciniphila)的相对丰度低于对照组儿童。见图 3。

|
| 图 3 对照组与肥胖组儿童共有菌属差异分析 |
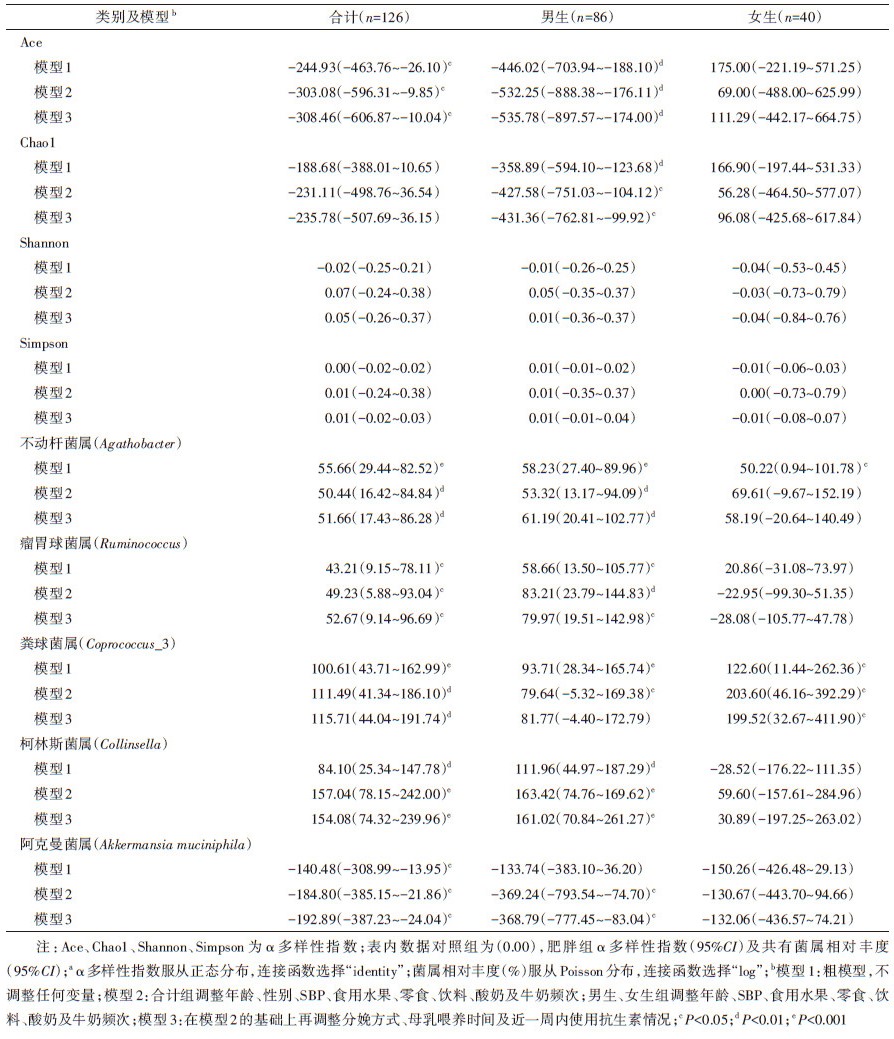
4. 肥胖与肠道菌群的α多样性及共有菌属相对丰度差异的关联分析:GLM分析结果显示,与对照组儿童相比,肥胖组的Ace指数和阿克曼菌属的相对丰度显著较低,粪球菌属、瘤胃球菌属、不动杆菌属、柯林斯菌属的相对丰度较高,校正年龄、性别、饮食频次等相关因素后,此关联依然存在,但并未发现肥胖对Shannon或Simpson指数有影响。根据性别分层之后,在肥胖男生中,另发现Chao1指数低于正常体重男生。但是,在肥胖女生中,仅发现粪球菌属高于正常体重女生。见表 3。
儿童肠道菌群在达到成年之前会发生较大变化,并且儿童时期的肥胖与肠道菌群的组成结构和数量比例有密切联系。本研究选择长期均处于肥胖状态的8~10岁学龄儿童进行观察。在考虑了宿主因素(年龄、性别等)和环境因素(分娩方式、抗生素使用、饮食习惯等)的影响之后,本研究结果显示,与正常体重儿童相比,肥胖儿童肠道菌群的多样性和菌属特征存在差异,且此差异在男生中更为明显。
α多样性主要衡量个体中物种丰度和多样性,肥胖和肠道菌群α多样性降低的关联在多数基于成年人的研究中得到了证实[15]。在本研究中,肥胖儿童的Ace和Chao1指数显著低于正常体重儿童,Chao1指数在肥胖男生中更低,而Shannon和Simpson指数并无组别差异,这与Le Chatelier等[16]的研究结果一致。通过结果可以看到,肥胖组与对照组肠道菌群多样性的差异仅表现在种群数量上,在物种分布均匀度上的差异并不明显。此外,基于非加权Unifrac距离和加权Unifrac距离的PCoA分析,并未发现组别对于菌群差异有显著贡献。但与此不同的是,Liu等[5]曾发现肥胖者肠道菌群的β多样性更高,说明肥胖个体之间有较高的肠道微生物群落异质性。
本研究观察到普雷沃菌属和八叠球菌属是肥胖儿童的特有菌属,这两个菌属仅在肥胖组儿童中被识别,且相对丰度较高。普雷沃菌属是肠型分类中的三大菌属之一,大部分对脂类代谢较弱的人群属于这类肠型。普雷沃菌属具有促炎作用,可诱导白介素(IL)-6(IL-6)、IL-8和肿瘤坏死因子-α等炎症因子的产生[17],而炎症是导致肥胖发生的重要因素[18]。八叠球菌属与胆固醇代谢密切相关[19],通过三甲胺-氧化三甲胺代谢途径,下调胆固醇7a-羟化酶的表达水平,激活法尼酯X受体,从而降低胆固醇的分解速率,并抑制肝脏胆汁酸的合成[20],胆汁酸通过肠肝循环(肝脏、肠道组织、血浆和粪便)参与调节食物摄入和能量代谢,这对于肥胖的发生发展至关重要[21]。
本研究利用STAMP软件挑选出肥胖和正常体重儿童共有菌属中的差异细菌,并做进一步的关联分析。在15个差异菌属中,有5个菌属的相对丰度差异 > 0.1%,结果提示,肥胖可能会提高不动杆菌属、瘤胃球菌属、粪球菌属和柯林斯菌属的相对丰度,而降低阿克曼菌属的相对丰度。Massier等[22]发现,与血液相比,脂肪组织中有更多的不动杆菌属。Lin等[23]在经高脂饮食诱导肥胖的大鼠粪便中,也观察到瘤胃球菌属和粪球菌属的相对丰度增加,这两个菌属均属于厚壁菌门,可协助细胞吸收食物中的糖类[24],促进脂肪组织扩张,与代谢紊乱和炎症性肠病相关[25]。柯林斯菌属在肠道中产气较多,与脂质代谢异常及2型糖尿病有关。与其他研究结果一致[26],本研究发现阿克曼菌属在正常体重者中的相对丰度大于肥胖者。阿克曼菌属是肠道的优势益生菌属,其通过调节肠道屏障的通透性,维持肠道屏障功能,可改善葡萄糖稳态,减少脂肪细胞生长[27]。此外,阿克曼菌属的定植加快黏蛋白的分解,产生多种微生物发酵产物如短链脂肪酸。短链脂肪酸主要由乙酸盐、丙酸盐和丁酸盐组成,在调节宿主代谢、免疫系统、细胞增殖和维持消化道健康等方面具有关键作用,可降低肥胖、糖尿病的风险[28]。
在调整相关因素之后,GLM分析的结果与之基本一致,且在男生中,肥胖与这5个菌属的关联性更强,但在女生中,大部分关联消失。既往研究也发现,肥胖与肠道菌群的关联在男性和女性中表现不同[29],这可能受性腺类固醇的影响[30],但本研究对象均为学龄早期儿童,性发育还未开始,因此本研究中所观察到的性别差异可能与样本量有关,在后续研究中可以扩大样本量进行验证。
本研究存在局限性,研究对象仅来自某一小学,代表性可能欠佳,此外,横断面数据分析也无法解释肥胖和肠道微生物多样性之间的因果关系。在物种识别过程中,由于较多菌种无法准确定义,因此在分析过程中仅讨论到属级别。
综上所述,本研究探索了学龄肥胖儿童和正常体重儿童肠道菌群多样性的差异,识别肥胖儿童肠道中的特征菌群,可为后续对于学龄儿童肠道微生物多态性的深入研究提供初步线索。肠道菌群种类较多,各菌属对宿主机体的作用机制各异,未来可利用相关功能细菌标志物,探索最佳的肥胖诊断和治疗方案。
利益冲突 所有作者声明无利益冲突
作者贡献声明 蒋露芳:实验操作、论文撰写;王莹莹:数据收集与分析、论文撰写;彭慧、李锐、张芬、邵月琴:数据收集;王娜、姜庆五:论文修改、经费支持
| [1] |
Pan XF, Wang LM, Pan A. Epidemiology and determinants of obesity in China[J]. Lancet Diabetes Endocrinol, 2021, 9(6): 373-392. DOI:10.1016/S2213-8587(21)00045-0 |
| [2] |
Llewellyn A, Simmonds M, Owen CG, et al. Childhood obesity as a predictor of morbidity in adulthood: a systematic review and meta-analysis[J]. Obes Rev, 2016, 17(1): 56-67. DOI:10.1111/obr.12316 |
| [3] |
Wang YY, Dong XL, Fu CW, et al. Thyroid Stimulating Hormone (TSH) is associated with general and abdominal obesity: a cohort study in school-aged girls during puberty in East China[J]. Front Endocrinol (Lausanne), 2020, 11: 620. DOI:10.3389/fendo.2020.00620 |
| [4] |
Zeng Q, Li NS, Pan XF, et al. Clinical management and treatment of obesity in China[J]. Lancet Diabetes Endocrinol, 2021, 9(6): 393-405. DOI:10.1016/S2213-8587(21)00047-4 |
| [5] |
Liu RX, Hong J, Xu XQ, et al. Gut microbiome and serum metabolome alterations in obesity and after weight-loss intervention[J]. Nat Med, 2017, 23(7): 859-868. DOI:10.1038/nm.4358 |
| [6] |
Walters WA, Xu Z, Knight R. Meta-analyses of human gut microbes associated with obesity and IBD[J]. FEBS Lett, 2014, 588(22): 4223-4233. DOI:10.1016/j.febslet.2014.09.039 |
| [7] |
Just S, Mondot S, Ecker J, et al. The gut microbiota drives the impact of bile acids and fat source in diet on mouse metabolism[J]. Microbiome, 2018, 6(1): 134. DOI:10.1186/s40168-018-0510-8 |
| [8] |
Hong J, Bo TT, Xi LQ, et al. Reversal of functional brain activity related to gut microbiome and hormones after VSG surgery in patients with obesity[J]. J Clin Endocrinol Metab, 2021, 106(9): e3619-3633. DOI:10.1210/clinem/dgab297 |
| [9] |
Cox AJ, West NP, Cripps AW. Obesity, inflammation, and the gut microbiota[J]. Lancet Diabetes Endocrinol, 2015, 3(3): 207-215. DOI:10.1016/S2213-8587(14)70134-2 |
| [10] |
Cotillard A, Kennedy SP, Kong LC, et al. Dietary intervention impact on gut microbial gene richness[J]. Nature, 2013, 500(7464): 585-588. DOI:10.1038/nature12480 |
| [11] |
Derrien M, Alvarez AS, de vos WM. The gut microbiota in the first decade of life[J]. Trends Microbiol, 2019, 27(12): 997-1010. DOI:10.1016/j.tim.2019.08.001 |
| [12] |
Casals-Pascual C, González A, Vázquez-Baeza Y, et al. Microbial diversity in clinical microbiome studies: sample size and statistical power considerations[J]. Gastroenterology, 2020, 158(6): 1524-1528. DOI:10.1053/j.gastro.2019.11.305 |
| [13] |
王向军, 杨漾, 吴艳强, 等. 上海市7~18岁学生1985至2014年的超重和肥胖流行趋势[J]. 中国循证儿科杂志, 2017, 12(2): 126-130. Wang XJ, Yang Y, Wu YQ, et al. Study on epidemiological trend of overweight and obesity school-age children and adolescents over the past 29 years (1985-2014) in Shanghai, China[J]. Chin J Evid Based Pediatr, 2017, 12(2): 126-130. DOI:10.3969/j.issn.1673-5501.2017.02.009 |
| [14] |
国家卫生和计划生育委员会. WS/T 586-2018学龄儿童青少年超重与肥胖筛查[S]. 北京: 中国标准出版社, 2018. National Health and Family Planning Commission of China. WS/T 586-2018 Screening for overweight and obesity among school-age children and adolescents[S]. Beijing: Standards Press of China, 2018. |
| [15] |
Aranaz P, Ramos-Lopez O, Cuevas-Sierra A, et al. A predictive regression model of the obesity-related inflammatory status based on gut microbiota composition[J]. Int J Obes (Lond), 2021, 45(10): 2261-2268. DOI:10.1038/S41366-021-00904-4 |
| [16] |
Le Chatelier E, Nielsen T, Qin JJ, et al. Richness of human gut microbiome correlates with metabolic markers[J]. Nature, 2013, 500(7464): 541-546. DOI:10.1038/nature12506 |
| [17] |
Larsen JM. The immune response to Prevotella bacteria in chronic inflammatory disease[J]. Immunology, 2017, 151(4): 363-374. DOI:10.1111/IMM.12760 |
| [18] |
Hotamisligil GS. Inflammation and metabolic disorders[J]. Nature, 2006, 444(7121): 860-867. DOI:10.1038/nature05485 |
| [19] |
Gutiérrez-Repiso C, Moreno-Indias I, de Hollanda A, et al. Gut microbiota specific signatures are related to the successful rate of bariatric surgery[J]. Am J Transl Res, 2019, 11(2): 942-952. |
| [20] |
Fennema D, Phillips IR, Shephard EA. Trimethylamine and trimethylamine N-Oxide, a Flavin-Containing Monooxygenase 3(FMO3)-mediated host-Microbiome metabolic axis implicated in health and disease[J]. Drug Metab Dispos, 2016, 44(11): 1839-1850. DOI:10.1124/dmd.116.070615 |
| [21] |
Prinz P, Hofmann T, Ahnis A, et al. Plasma bile acids show a positive correlation with body mass index and are negatively associated with cognitive restraint of eating in obese patients[J]. Front Neurosci, 2015, 9: 199. DOI:10.3389/fnins.2015.00199 |
| [22] |
Massier L, Chakaroun R, Tabei S, et al. Adipose tissue derived bacteria are associated with inflammation in obesity and type 2 diabetes[J]. Gut, 2020, 69(10): 1796-1806. DOI:10.1136/gutjnl-2019-320118 |
| [23] |
Lin H, An YP, Tang HR, et al. Alterations of bile acids and gut microbiota in obesity induced by high fat diet in rat model[J]. J Agric Food Chem, 2019, 67(13): 3624-3632. DOI:10.1021/acs.jafc.9b00249 |
| [24] |
Pedersen HK, Gudmundsdottir V, Nielsen HB, et al. Human gut microbes impact host serum metabolome and insulin sensitivity[J]. Nature, 2016, 535(7612): 376-381. DOI:10.1038/nature18646 |
| [25] |
Peterson CT, Sharma V, Elmén L, et al. Immune homeostasis, dysbiosis and therapeutic modulation of the gut microbiota[J]. Clin Exp Immunol, 2015, 179(3): 363-377. DOI:10.1111/cei.12474 |
| [26] |
Giannoudaki E, Hernandez-Santana YE, Mulfaul K, et al. Interleukin-36 cytokines alter the intestinal microbiome and can protect against obesity and metabolic dysfunction[J]. Nat Commun, 2019, 10(1): 4003. DOI:10.1038/s41467-019-11944-w |
| [27] |
Macchione IG, Lopetuso LR, Ianiro G, et al. Akkermansia muciniphila: key player in metabolic and gastrointestinal disorders[J]. Eur Rev Med Pharmacol Sci, 2019, 23(18): 8075-8083. DOI:10.26355/eurrev_201909_19024 |
| [28] |
Xu Y, Wang N, Tan HY, et al. Function of Akkermansia muciniphila in Obesity: interactions with lipid metabolism, immune response and gut systems[J]. Front Microbiol, 2020, 11: 219. DOI:10.3389/fmicb.2020.00219 |
| [29] |
Gao XF, Zhang MR, Xue JM, et al. Body mass index differences in the gut microbiota are gender specific[J]. Front Microbiol, 2018, 9: 1250. DOI:10.3389/fmicb.2018.01250 |
| [30] |
Mayneris-Perxachs J, Arnoriaga-Rodríguez M, Luque-Córdoba D, et al. Gut microbiota steroid sexual dimorphism and its impact on gonadal steroids: influences of obesity and menopausal status[J]. Microbiome, 2020, 8(1): 136. DOI:10.1186/s40168-020-00913-x |