 2018, Vol. 39
2018, Vol. 39文章信息
- 王茜, 李木丽, 陈叶, 王冰, 陶晓燕, 朱武洋.
- Wang Qian, Li Muli, Chen Ye, Wang Bing, Tao Xiaoyan, Zhu Wuyang.
- 中国不同地区狂犬病病毒种群分布的差异
- Differences on geographic distribution of rabies virus lineages in China
- 中华流行病学杂志, 2018, 39(4): 491-494
- Chinese Journal of Epidemiology, 2018, 39(4): 491-494
- http://dx.doi.org/10.3760/cma.j.issn.0254-6450.2018.04.021
-
文章历史
收稿日期: 2017-08-08




2. 518020 深圳市人民医院病理科;
3. 110031 沈阳市疾病预防控制中心
2. Department of Pathology, Shenzhen People's Hospital, Shenzhen 518020, China;
3. Shenyang Center for Disease Control and Prevention, Shenyang 110031, China
狂犬病疫情在我国有明显的地域差异。自2005年起国家狂犬病监测实验室在全国范围开展病原学监测,已初步确定我国流行的狂犬病病毒(RABV)可分为6个种群(ChinaⅠ~Ⅵ),每个毒株群的规模和流行范围各不相同[1-4]。尽管2013年在吉林省蝙蝠中监测到Ikrut病毒[5],表明我国还存在狂犬病病毒属(Lyssavirus)蝙蝠狂犬病的流行,但影响人狂犬病疫情的仍是RABV[6-7]。我国狂犬病防控疫情更多是以省为单位,甚至范围更小[8],而不同省份流行的RABV病原是否有差异,是单一种群的流行还是多种群病毒交叉存在,不同种群病毒在区域水平的分布特点尚未明确。为此本研究探讨我国各省份RABV流行的种群类别和不同种群的流行规模及流行范围变化,为当地有针对性地制定有效防控措施提供科学依据。
资料与方法1.资料来源:中国狂犬病流行株(即街毒株,不包括固定毒株、疫苗株和基因突变株等衍生毒株)的N、G基因和基因组全序列及毒株背景信息来源于美国NCBI网站GenBank数据库(截至2016年12月31日),经Excel 2013软件整理为“中国狂犬病流行株基因数据库”,包括712个毒株。再纳入本实验室2015年在辽宁省监测到的2个毒株[6],数据库流行株总数为714个。纳入的流行株宿主来源包括病例、犬、狐狸、鼬獾、貉、牛、骡、驴、羊、骆驼、猪、马、鹿和鼠13种动物。
2.种系发生分析:病毒N、G基因在进化分析中的地位是等同[2-3, 9],有N基因或全基因组的毒株选取N基因全序,只有G基因序列的毒株整理G基因全序,以MEGA 7.0软件构建N和G基因种系发生树[6],明确各毒株的种群(China Ⅰ~ Ⅵ)划分。
结果1. RABV种群地区分布:云南和湖南是我国大陆RABV种群最丰富的省份,均有4个种群类型流行,且均以China Ⅰ群为主;河南、福建、贵州、广西、宁夏和内蒙古6个省份均为3个种群同时流行,种群类型也较为丰富;东部地区的上海、江苏、浙江、山东、安徽、江西和西南部地区的重庆以及东北部地区的吉林8省份均有2个种群共存;其余14个省份目前只监测到1个毒株群流行(表 1)。
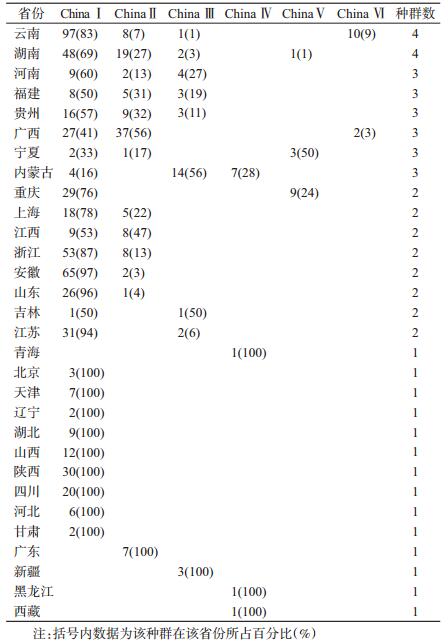
2. RABV种群流行规模:我国目前流行的RABV种群依然为ChinaⅠ~Ⅵ,类型未增加[2-3](图 1)。China Ⅰ是我国的优势种群,毒株数量占我国流行株总数的74.79%,分布范围广泛(25个省份),不仅贯通南北地区,且已蔓延至东北部和西部地区。目前只有西部地区的西藏、青海、新疆以及东北部地区的黑龙江尚未监测到China Ⅰ;南部地区的广东作为传统高发省份,尚未发现有China Ⅰ流行,可能与毒株数据较少有关[2]。

|
| 注:A、B、C分别为ChinaⅠ~Ⅵ群分布,D为ChinaⅣ~Ⅵ群分布;地图版本为ArcGIS10.3.1 图 1 我国狂犬病病毒种群(China Ⅰ~Ⅵ)地域分布 |
China Ⅱ的流行规模仅次于China Ⅰ,数量占我国毒株总数的15.67%,主要分布于我国南部省份(广东、广西)、东部省份(福建、江西、浙江、安徽、山东、上海)以及西南部省份(云南、贵州)和中部省份(湖南、河南);1986年自宁夏分离的1株病毒也属China Ⅱ。
China Ⅲ的数量占我国毒株总数的4.62%,目前已流行于吉林、江苏、内蒙古、湖南、福建、河南、新疆、贵州和云南9个省份,相比较China Ⅰ和Ⅱ,分布较为分散。
其他3个种群,毒株数量较少,流行区域局限。China Ⅳ数量占我国毒株总数的1.40%,在内蒙古、青海、西藏和黑龙江4省份有报道;目前China Ⅴ仅自重庆、宁夏和湖南3个省份的标本中分离到,占我国毒株总数的1.82%;China Ⅵ数量占我国毒株总数的1.68%,目前仅分布于云南和广西地区。
讨论目前我国不同省份流行的病原种群数量为1~4种,通常疫情较为严重且持续时间较长的省份其病原种群类型较多[2],如2001和2002年高居全国疫情首位且之后长期列前5位的湖南省,病原种群类型多达4个,凸显该地区在病原传播扩散中的汇集、中转作用[10]。云南省也拥有丰富的病原种群类型[11]。宁夏地区2000年前后基本处于无疫情状态,自2011年后才开始疫情上升,有3个种群类型,属于China Ⅱ和Ⅴ的毒株均是1986年分离获得,近年监测到的流行株均属于China Ⅰ[12]。以往的研究结果提示China Ⅰ在1996年以后替代China Ⅱ成为目前优势毒株群[2],宁夏地区上一流行阶段的种群(China Ⅱ、Ⅴ)可能已经不复存在,目前只受China Ⅰ的影响,该地区种群的多样性是长期监测的结果。因此病原数量少的省份(如广东)更应持续开展监测。
根据最新分析数据,我国大陆分布的RABV病原种群类别依然是6群,而随着监测的深入开展,各种群的流行规模均有所扩展,如以往推测[2, 6],China Ⅰ仍为目前狂犬病疫情优势毒株群,已扩散至我国东北及西部地区。尚未被China Ⅰ入侵的青海、西藏和黑龙江,目前流行的种群为主要在野生动物中流行的China Ⅳ,与以犬传播为主的China Ⅰ不同,尚未在与人更为密切的犬中建立循环,当地疫情主要是野生动物狂犬病溢出的结果,所以一般疫情上升压力较小[6]。因此,流行病原种群不同的省份应采取不同针对性的防控措施。
利益冲突: 无
| [1] | Badrane H, Tordo N. Host switching in Lyssavirus history from the chiroptera to the carnivora orders[J]. J Virol, 2001, 75(17): 8096–8104. DOI:10.1128/JVI.75.17.8096-8104.2001 |
| [2] | Tao XY, Tang Q, Rayner S, et al. Molecular phylodynamic analysis indicates lineage displacement occurred in Chinese rabies epidemics between 1949 to 2010[J]. PLoS Negl Trop Dis, 2013, 7(7): e2294. DOI:10.1371/journal.pntd.0002294 |
| [3] | Guo ZY, Tao XY, Yin CP, et al. National borders effectively halt the spread of rabies:the current rabies epidemic in China is dislocated from cases in neighboring countries[J]. PLoS Negl Trop Dis, 2013, 7(1): e2039. DOI:10.1371/journal.pntd.0002039 |
| [4] |
陶晓燕, 李浩, 焦文涛, 等. 中国狂犬病病毒的分群和进化特征[J]. 疾病监测, 2013, 28(5): 340–343.
Tao XY, Li H, Jiao WT, et al. Grouping and evolution characteristics of rabies virus in China[J]. Dis Surveill, 2013, 28(5): 340–343. DOI:10.3784/j.issn.1003-9961.2013.05.003 |
| [5] | Liu Y, Zhang SF, Zhao JH, et al. Isolation of Irkut virus from a Murina leucogaster bat in China[J]. PLoS Negl Trop Dis, 2013, 7(3): e2097. DOI:10.1371/journal.pntd.0002097 |
| [6] | Tao XY, Guo ZY, Li H, et al. Rabies cases in the west of China have two distinct origins[J]. PLoS Negl Trop Dis, 2015, 9(10): e0004140. DOI:10.1371/journal.pntd.0004140 |
| [7] | Zhou H, Vong S, Liu K, et al. Human rabies in China, 1960-2014:a descriptive epidemiological study[J]. PLoS Negl Trop Dis, 2016, 10(8): e0004874. DOI:10.1371/journal.pntd.0004874 |
| [8] | Song M, Tang Q, Rayner S, et al. Human rabies surveillance and control in China, 2005-2012[J]. BMC Infect Dis, 2014, 14: 212. DOI:10.1186/1471-2334-14-212 |
| [9] | Wu XF, Franka R, Velasco-Villa A, et al. Are all Lyssavirus genes equal for phylogenetic analyses?[J]. Virus Res, 2007, 129(2): 91–103. DOI:10.1016/j.virusres.2007.06.022 |
| [10] | Tao XY, Tang Q, Li H, et al. Molecular epidemiology of rabies in southern People's Republic of China[J]. Emerg Infect Dis, 2009, 15(8): 1192–1198. DOI:10.3201/eid1508.081551 |
| [11] | Zhang HL, Zhang YZ, Yang WH, et al. Molecular epidemiology of reemergent rabies in Yunnan province, southwestern China[J]. Emerg Infect Dis, 2014, 20(9): 1433–1442. DOI:10.3201/eid2009.130440 |
| [12] | Lang SL, Tao XY, Guo ZY, et al. Molecular characterization of viral G gene in emerging and re-emerging areas of rabies in China, 2007 to 2011[J]. Virol Sin, 2012, 27(3): 194–203. DOI:10.1007/s12250-012-3248-7 |