 2016, Vol. 37
2016, Vol. 37文章信息
- 冯云, 张海林, 杨卫红, 章域震, 黄丽娟, 邓淑珍, 孙玉杰, 杨杜鹃, 周济华 .
- Feng Yun, Zhang Hailin, Yang Weihong, Zhang Yuzhen, Huang Lijuan, Deng Shuzhen, Sun Yujie, Yang Dujuan, Zhou Jihua .
- 云南省1977-2010年流行性乙型脑炎病毒分子流行病学研究
- Molecular epidemiology of Japanese encephalitis viruses isolated in Yunnan province, 1977-2010
- 中华流行病学杂志, 2016, 37(11): 1519-1525
- Chinese journal of Epidemiology, 2016, 37(11): 1519-1525
- http://dx.doi.org/10.3760/cma.j.issn.0254-6450.2016.11.017
-
文章历史
收稿日期: 2016-05-20




流行性乙型脑炎(乙脑)是由乙脑病毒(JEV)引起经蚊媒传播的一种中枢神经系统感染的急性传染病。我国各地几乎都有流行[1-2]。云南省是我国乙脑高发地区,为当地分布最广、危害较大的一种虫媒病毒病[3-4]。此前,我们已从云南省不同地区患者及蚊虫、蠓、鸟类和蝙蝠中分离出100多株JEV[4-9],但大多数分离株仅进行了C/PrM区序列测定和分析,缺少系统的基因分型和分子流行病学研究。本研究对1977-2010年分离自云南省的63株JEV进行包膜蛋白(envelope protein,E)全编码区基因序列测定与分析,以期阐明云南省JEV基因型变化及其与其他省区和国外流行株的进化关系。
1、 材料与方法1.病毒株:选择本室分离自云南的63株乙脑病毒,其中20世纪70年代2株,80年代10株,90年代3株,2004年3株,2007年20株,2009年9株,2010年16株。
2.病毒株的复苏:从低温冰箱中取出冻干管,每管加入0.5 ml的基础培养基(MEM),10 000 rpm/min冷冻离心10 min,取上清液,颅内接种乳小白鼠(昆明种小白鼠,大理大学实验动物中心提供),每只接种0.02 ml,每天观察发病状况,如乳鼠发病处于濒死症状时,解剖取脑组织制成接种液,再传代一次,乳鼠规律发病者剖取脑组织制备病毒悬液,-20 ℃冰箱备用。每株病毒接种乳鼠一窝(8~10只),其中4只乳鼠用于制备病毒样本,其余乳鼠用于观察发病死亡状况。
3. RT-PCR和序列测定:取病毒分离物上清液,用QIAamp Viral RNA Mini Kit(美国QIAGEN公司)按说明书提取病毒RNA,用Amersham Bioscience Ready-to-GoTM You First-Strand Beads(美国Amersham Pharmacia Biotech公司)制备cDNA。取2 μl cDNA做反应模板,用JEV E基因引物进行PCR扩增(表 1)。依次加入10×Buffer 5 μl、2.5 mmol/L dNTP 5 μl、上下游引物各1.25 μl、ExTaq酶0.75 μl、去离子水31.75 μl,充分混匀,94 ℃预变性5 min,94 ℃ 30 s,55 ℃ 30 s,72 ℃ 60 s,35个循环,72 ℃延伸10 min,反应结束后,取2 μl产物进行1%琼脂糖凝胶电泳检测扩增效果。所有扩增产物的测序均在生工生物工程(上海)股份有限公司完成。
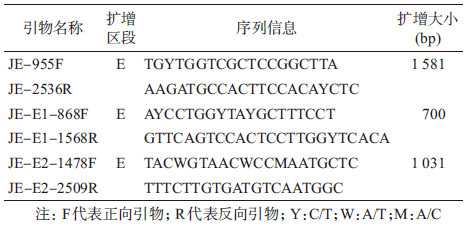
4.序列分析:使用DNAstar软件包中的SeqMan对序列片段进行拼接、编辑、校正。Clustal X 1.83和DNAStar软件包中的Megalign、Alignment等生物学软件进行核苷酸和氨基酸序列分析。采用Mega 5.0软件邻接法(neighbor-joining)进行JEV基因核苷酸序列系统发生树分析。本研究引用GenBank中来自不同国家和地区、不同年代的42株JEV的E区段核苷酸序列,其中基因1型20株,2型5株,3型14株,4型2株,5型1株,外群为墨累山谷脑炎病毒(MVE-1-51株)。
2、 结果1.病毒传代:63株JEV均能在乳鼠脑内增殖,并引起乳鼠发病和死亡,潜伏期2~3 d,主要表现为拒乳、震颤、抽搐、昏迷、瘫痪等,接种乳鼠的病死率为100%,一般于接种后70~80 h死亡,乳鼠接种到死亡时间的间隔中位数为73 h;大多于发病后10~20 h死亡,乳鼠发病到死亡时间的间隔中位数为16 h。结果表明这些病毒株在低温保存状态下仍然保持其较强的致病性和毒力。
2.序列测定:经E基因序列测定,获得这63株JEV的E蛋白编码区全基因组序列,长度为1 500 bp,位于病毒基因组第978~2 477位。
3.进化分析:云南63株JEV与来自GenBank中42株不同基因型JEV的E基因核苷酸序列构建系统进化树,其中云南47株JEV与基因1型同为一个进化分支,云南16株JEV与基因3型同处一个进化分支,所有云南株与基因2、4和5型JEV的进化关系稍远,与外群病毒MVE-1-51株差异较大(图 1)。云南省47株基因1型JEV中,分离自1977年1株,1982年1株,2007年20株,2009年9株,2010年16株;16株基因3型JEV中,分离自1978年1株,1981年1株,1982年2株,1983年1株,1986年1株,1987年1株,1989年3株,1990年2株,1997年1株,2004年3株。

|
| 图 1 云南省63株JEV与GenBank中42株乙脑1~5基因型病毒的E区核苷酸序列系统进化分析 |
本次获得的63株JEV的E基因序列已提交GenBank,其中47株基因1型接受号为KU295054~KU295100,16株基因3型接受号为KU295101~KU295116。
基因1型JEV进化树显示,47株云南基因1型大致可分为2个进化群(图 2),其中2007-2010年分离自云南省东、南、西和中部共45株病毒同为一个进化群,但分布于两个次级进化分支,其中40株与我国四川、广西、河南、甘肃、辽宁、上海和台湾地区以及韩国、越南流行株亲缘关系较近,同处一个进化支;2007年分离自滇西的5株病毒聚集为一个进化支,与山东(SD0810)、云南(YN05155、YN05124)、日本(Ishikawa)和辽宁(LN02102)、上海(SH80)以及韩国(K94P05)流行株亲缘性较近。1977和1982年分离自云南的M28和BN82215与泰国早期分离株(U70416,1982;DQ084229,年代不详)同处一个进化支,亲缘关系最为接近。图 2还显示,泰国1979年基因1型分离株(U70401)与云南M28和BN82215以及其他基因1型均有明显差异。

|
| ▲为云南分离株 图 2 云南省47株基因1型JEV与来自GenBank中20株基因1型JEV的E区核苷酸序列系统进化分析 |
基因3型JEV进化树显示,16株云南基因3型可分为3个进化群(图 3),20世纪80和90年代分离自云南南部和西部的9株病毒为同一进化群,并与日本、斯里兰卡和Beijing-1病毒株高度同源;2004年分离自滇西南的5株与同期分离自广东(GZ042,2004)、黑龙江(HLJ02134,2002)和福建(FJ0394,2003)为一个进化群;20世纪70和80年代分离自滇西南的4株病毒与韩国(K87P39,1987)、印度尼西亚(JKT1724,1979)和尼泊尔(B2524,1985)分离株同为一个进化群。

|
| ▲为云南分离株 图 3 云南省16株基因3型JEV与GenBank中14株基因3型JEV的E区核苷酸序列系统进化分析 |
4.同源性分析:根据进化树分析结果,选择云南基因1型和3型JEV不同时期代表株各10株并与来自GenBank的19株不同基因型JEV的E基因进行核苷酸和氨基酸同源性分析。结果显示,云南基因1型间的核苷酸(氨基酸)同源性为94.9%~100.0%(98.8%~100.0%),与7株基因1型参考株的核苷酸(氨基酸)同源性为94.4%~99.1%(98.6%~100.0%),与基因2、3、4和5型参考株依次为88.8%~89.0%(98.2%~98.6%)、85.0%~87.2%(95.0%~98.4%)、81.1%~82.2%(94.8%~95.2%)、76.2%~77.4%(91.0%~91.6%);云南基因3型间的核苷酸(氨基酸)同源性为96.1%~99.7%(97.6%~99.8%),与6株基因3型参考株的核苷酸(氨基酸)同源性为95.4%~99.8%(96.4%~99.8%),与基因1、2、4和5型分别为87.5%~88.6%(96.0%~96.8%)、87.8%~88.9%(97.2%~97.8%)、83.0%~83.8%(94.0%~96.0%)、77.1%~77.7%(90.8%~91.6%)。虽然基因1型或3型JEV的核苷酸和氨基酸间均存在一定差异,但与其他型别的JEV株存在明显差异。
5.氨基酸抗原和毒力位点:选择云南不同时期分离的JEV基因1型(8株)和基因3型(16株)与我国广泛使用的乙脑减毒活疫苗SA14-14-2株进行比较,所有云南株与SA14-14-2株在E蛋白重要抗原表位相关的结构域Ⅰ(E1~E 51)、Ⅱ(E52~E137、E197~E292)和Ⅲ(E310~E411)的位点相同,仍然保持其抗原特性。所有云南株与SA14-14-2株在E蛋白毒力相关的8个重要位点(E107、E138、E167、E177、E264、E279、E315、E439)中存在7个位点差异,仅E167位点同为F。
3、 讨论JEV基因型研究对阐明该病毒的遗传进化和流行病学特征均有重要意义。起初,Chen等[10]根据JEV C/PrM区段的240个核苷酸将JEV分为4个基因型;随后,为避免因较短的核苷酸序列分析可能导致结果的不准确性,Solomon等[11]采用E基因的1 500个核苷酸对JEV进行基因分型,将JEV分为5个基因型,并得到国际公认。其中1和3型分布较广,2、4和5型分布较局限。我国广泛存在1和3型[12-13],5型仅在西藏林芝地区蚊虫中分离到[14]。以往,我们曾采用C/PrM区段对早期分离自云南省的19株JEV进行分型[15],其中17株为基因3型,2株为基因1型。本研究采用E基因对时间跨度长达34年(1977-2010年)的云南63株JEV进行基因型研究,无论是系统进化树或同源性分析,两法分型结果一致,阐明了云南省存在基因1和3型JEV的流行,但不同基因型病毒在时间和地域分布上存在明显差异。云南基因1型最早分离于1977和1982年,在随后25年间未再分离到该型病毒,直至2007年再次分得,此后分离的均为基因1型;1978-2004年的分离株几乎均为基因3型(仅1982年分离到1株基因1型)。这些结果表明,云南省JEV具有基因1型替换基因3型成为主要流行型的趋势。
国外研究认为,东南亚热带地区是JEV的重要起源地,最早JEV可能起源于印度尼西亚及马来西亚地区,伴随着乙脑基因型的遗传进化,该病毒在亚洲及其周边地区不断蔓延扩散[11]。JEV基因型研究和进化分析表明,1970年代之前,亚洲流行的JEV均为基因3型,随后出现1、2、4、5型[10-11],近10年1型已成为主要流行型[13]。高晓艳等[16-17]研究认为,亚洲南端是JEV基因1型的起源地。云南省南部和西部与乙脑主要流行区的越南、泰国、老挝和缅甸相邻,北部和东部与我国乙脑主要流行区四川、贵州、广西相连,自然环境较为特殊,加之三带喙库蚊等乙脑传播媒介广泛分布[6-9],养猪较普遍,也是候鸟迁徙地,适于JEV传播和扩散。本研究发现,云南基因1型病毒株间存在明显差异,如云南最早(1977和1982年)分离的基因1型病毒与泰国1982年分离株(U70416)和DQ084229株(分离年代不详)基因1型病毒同处一个进化群,并与2007-2010年云南分离株存在明显差异,还发现2007-2010年的基因1型云南分离株大多为同一进化群。根据GenBank中的JEV序列,云南1977年分离的基因1型M28株(KU295054)属亚洲最早分离的基因1型JEV,之后为泰国分离株(U70401,1979),至1982年中国云南省和泰国再次分离到该型病毒。2001年王环宇等[18]从捕自上海奉贤的蚊虫中分离到基因1型JEV,2003-2005年泰国和我国其他省区相继也分离到该型病毒[2, 19-20]。由此推测,中国云南省及相邻的泰国等地可能是基因1型JEV的起源地,并逐渐扩散至我国及亚洲其他地区。有关基因1型JEV扩散和演变机制尚需进一步研究。
云南基因3型JEV同样存在差异,如20世纪70、80和90年代的病毒株可分为两个进化群,并均与亚洲其他国家流行株亲缘性较近,而2004年云南基因3型分离株则自成一个进化群并与同期我国其他省区流行株具有较近的亲缘关系,提示该进化群可能为我国后期基因3型的主要流行病毒株。综合云南基因1和3型JEV的进化分析,认为在自然界中,JEV随着时间的推移并与宿主的共进化,该病毒发生某些改变,以至云南省JEV流行株具有多样性特点,并发现云南株不同进化群具有时间和地域分布特点。
我国现在使用的乙脑减毒活疫苗株和灭活疫苗株均为基因3型病毒,而基因1型的广泛流行是否会带来疫苗免疫保护问题,是当前人们关注的问题。由于JEV仅有一个血清型[2],加之本研究证实,云南JEV流行株中,无论是基因1型还是3型,或不同进化群,它们的抗原性重要功能位点均未发生明显改变,从理论上提示该乙脑疫苗的免疫能保护基因1型病毒的感染。JEV的减毒机制研究认为[21],E138位的谷氨酸(E)替换成赖氨酸(K)后病毒的毒力会有较明显的降低,但本研究中所有云南株的E138位仍为谷氨酸(E),由此认为,云南不同时期流行的基因1型或3型JEV均具有较强的致病性和毒力。为进一步阐明这些问题,尚需深入开展致病性、抗原性、免疫原性及免疫效果等研究。
利益冲突: 无| [1] | Wang HY, Li YX, Liang XF, et al. Japanese encephalitis in mainland China[J]. Jpn J Infect Dis , 2009, 62(5) : 331–336. |
| [2] |
张海林.流行性乙型脑炎//曹务春.流行病学:第二卷[M]. 3版.北京:人民卫生出版社, 2015:798-810.
Zhang HL. Japanese encephalitis//Cao WC. Epidemiology:Vol. 2[M]. 3rd ed. Beijing:People's Health Press, 2015:798-810. |
| [3] |
邓淑珍, 张海林, 刘晓强, 等.
1976-2007年云南省流行性乙型脑炎流行病学特征分析[J]. 地方病通报 , 2009, 24(3) : 1–4, 7.
Deng SZ, Zhang HL, Liu XQ, et al. Analysis of epidemiological characteristics of Japanese encephalitis in Yunnan Province from 1976 to 2007[J]. Endem Dis Bull , 2009, 24(3) : 1–4, 7. DOI:10.13215/j.cnki.jbyfkztb.2009.03.061 |
| [4] | Feng Y, Fu SH, Zhang HL, et al. High incidence of Japanese encephalitis, Southern China[J]. Emerg Infect Dis , 2013, 19(4) : 672–673. DOI:10.3201/eid1904.120137 |
| [5] | Feng Y, Fu SH, Zhang HL, et al. Distribution of mosquitoes and mosquito-borne viruses along the China-Myanmar Border in Yunnan Province[J]. Jpn J Infect Dis , 2012, 65(3) : 215–221. DOI:10.7883/yoken.65.215 |
| [6] |
邓淑珍, 张海林, 李金梅.
云南省蚊虫分布特点及自然感染乙型脑炎病毒的研究[J]. 中国媒介生物学及控制杂志 , 2009, 20(4) : 344–348.
Deng SZ, Zhang HL, Li JM. Distribution characteristics of mosquito and their natural infection with Japanese encephalitis virus in Yunnan province[J]. Chin J Vector Biol Control , 2009, 20(4) : 344–348. |
| [7] |
杨杜鹃, 付士红, 张海林, 等.
云南省东北等地区蚊虫及蚊媒病毒调查研究[J]. 中国媒介生物学及控制杂志 , 2011, 22(4) : 304–308, 312.
Yang DJ, Fu SH, Zhang HL, et al. Investigation of mosquitoes and mosquito-borne viruses in northeast Yunnan province[J]. Chin J Vector Biol Control , 2011, 22(4) : 304–308, 312. |
| [8] | Zhang HL, Zhang YZ, Yang WH, et al. Mosquitoes of western Yunnan province, China:seasonal abundance, diversity, and arbovirus associations[J]. PLoS One , 2013, 8(10) : e77017. DOI:10.1371/journal.pone.0077017 |
| [9] |
冯云, 张海林, 付士红, 等.
云南省德宏州2007年和2010年蚊虫及蚊媒病毒调查[J]. 中华流行病学杂志 , 2014, 35(5) : 528–532.
Feng Y, Zhang HL, Fu SH, et al. Investigation on mosquitoes and mosquito-borne viruses in Dehong prefecture, Yunnan province, 2007 and 2010[J]. Chin J Epidemiol , 2014, 35(5) : 528–532. DOI:10.3760/cma.j.issn.0254-6450.2014.05.013 |
| [10] | Chen WR, Tesh RB, Rico-Hesse R. Genetic variation of Japanese encephalitis virus in nature[J]. J Gen Virol , 1990, 71(12) : 2915–2922. DOI:10.1099/0022-1317-71-12-2915 |
| [11] | Solomon T, Ni HL, Beasley DWC, et al. Origin and evolution of Japanese encephalitis virus in Southeast Asia[J]. J Virol , 2003, 77(5) : 3091–3098. DOI:10.1128/JVI.77.5.3091-3098.2003 |
| [12] | Wang HY, Takasaki T, Fu SH, et al. Molecular epidemiological analysis of Japanese encephalitis virus in China[J]. J Gen Virol , 2007, 88(3) : 885–894. DOI:10.1099/vir.0.82185-0 |
| [13] | Pan XL, Liu H, Wang HY, et al. Emergence of genotypeⅠof Japanese encephalitis virus as the dominant genotype in Asia[J]. J Virol , 2011, 85(19) : 9847–9853. DOI:10.1128/JVI.00825-11 |
| [14] | Li MH, Fu SH, Chen WX, et al. Molecular characterization of full-length genome of Japanese encephalitis virus genotypeⅤ isolated from Tibet, China[J]. Biomed Environ Sci , 2014, 27(4) : 231–239. DOI:10.3967/bes2014.046 |
| [15] |
王静林, 张海林, 周济华, 等.
云南省乙型脑炎病毒基因分型研究[J]. 中华实验和临床病毒学杂志 , 2008, 22(2) : 87–90.
Wang JL, Zhang HL, Zhou JH, et al. Genotyping of Japanese encephalitis viruses isolated in Yunnan[J]. Chin J Exp Clin Virol , 2008, 22(2) : 87–90. DOI:10.3760/cma.j.issn.1003-9279.2008.02.003 |
| [16] | Gao XY, Liu H, Wang HY, et al. Southernmost Asia is the source of Japanese encephalitis virus (Genotype 1) diversity from which the viruses disperse and evolve throughout Asia[J]. PLoS Negl Trop Dis , 2013, 7(9) : e2459. DOI:10.1371/journal.pntd.0002459 |
| [17] |
高晓艳, 周海卫, 刘红, 等.
乙脑病毒的空间播散及迁徙事件研究[J]. 病毒学报 , 2015, 31(3) : 264–268.
Gao XY, Zhou HW, Liu H, et al. Study on spatial dispersal and migration events of Japanese encephalitis virus[J]. Chin J Virol , 2015, 31(3) : 264–268. DOI:10.13242/j.cnki.bingduxuebao.002694 |
| [18] |
王环宇, 付士红, 李晓宇, 等.
我国首次分离到基因Ⅰ型乙型脑炎病毒[J]. 中华微生物学和免疫学杂志 , 2004, 24(11) : 843–849.
S H, Li XY, et al. Isolation and identification of genotypeⅠ Japanese encephalitis virus in China[J]. Chin J Microbiol Immunol , 2004, 24(11) : 843–849. DOI:10.3760/j.issn.0254-5101.2004.11.001 |
| [19] | Nitatpattana N, Dubot-Pérès A, Gouilh MA, et al. Change in Japanese encephalitis virus distribution, Thailand[J]. Emerg Infect Dis , 2008, 14(11) : 1762–1765. DOI:10.3201/eid1411.080542 |
| [20] | Chen YY, Fan YC, Tu WC, et al. Japanese encephalitis virus genotype replacement, Taiwan, 2009-2010[J]. Emerg Infect Dis , 2011, 17(12) : 2354–2356. DOI:10.3201/eid1712.110914 |
| [21] | Ni HL, Chang GJJ, Xie H, et al. Molecular basis of attenuation of neurovirulence of wild-type Japanese encephalitis virus strain SA14[J]. J Gen Virol , 1995, 76(2) : 409–413. DOI:10.1099/0022-1317-76-2-409 |