 2015, Vol. 36
2015, Vol. 36文章信息
- 周璕, 张永红, 江波, 张诗卉, 高昕, 彭浩, 王艾丽. 2014.
- Zhou Xun, Zhang Yonghong, Jiang bo, Zhang Shihui, Gao Xin, Peng Hao, Wang Aili. 2014.
- 杀伤细胞免疫球蛋白样受体基因多态性与原发性高血压关系的病例对照研究
- Association between polymorphisms of killer cell immunoglobulin-like receptor gene and the risk of essential hypertension:a case-control study
- 中华流行病学杂志, 2015, 36(6): 589-593
- Chinese Journal of Epidemiology, 2015, 36(6): 589-593
- http://dx.doi.org/10.3760/cma.j.issn.0254-6450.2015.06.011
-
文章历史
- 投稿日期:2014-11-11




2. 215123 苏州大学 2012级临床医学五年制;
3. 江南大学附属医院
2. Grade 2012, Five-Year Program Clinic Medicine, Medical College of Soochow University, Suzhou 215123, China;
3. Affiliated Hospital of Jiangnan University
原发性高血压的危险因素有高钠低钾饮食、超重/肥胖、饮酒、吸烟和年龄等[1, 2]。但这些危险因素并不能解释所有的原发性高血压发病。近年研究提示,原发性高血压可能是一种自身免疫性炎症疾病[3, 4, 5],炎症和血管内皮功能异常可能在原发性高血压发病过程中发挥作用[6, 7, 8, 9],而自身免疫反应促进了炎症的发展[10]。杀伤细胞免疫球蛋白样受体(KIR)主要表达于自然杀伤(NK)细胞和部分T淋巴细胞表面,通过转导活化或抑制信号调节细胞功能。研究表明KIR基因簇多态性在自身免疫性疾病的发病过程中发挥作用。Yen等[11]研究发现,携带KIR2DS2基因型的个体,患银屑病合并脉管炎的危险性显著升高(OR=5.56,P=0.001)。作为调节自身免疫的重要受体,KIR基因多态性在高血压发生发展中的作用尚未明确。目前国内外鲜见KIR基因多态性与原发性高血压关系的研究报道,而流行病学的研究尚未见报道。因此,本研究采用病例对照方法探讨KIR基因多态性与原发性高血压的关系,为原发性高血压发病相关的炎症机制研究提供线索。 对象与方法
1. 研究对象:来自2010年3-5月苏州市横断面抽样调查所获得的汉族人群,共3 016人。病例组为随机选取的原发性高血压患者,符合1999年WHO/ISH高血压诊断标准[SBP≥140 mmHg(1 mmHg=0.133 kPa)和/或DBP≥90 mmHg,或者正在服用降压药物];对照组纳入标准为SBP<140 mmHg 且DBP<90 mmHg,以及近两周内未服用降压药者。两组人群排除继发性高血压、冠心病、肾病和糖尿病患者。使用SAS软件将病例与对照按年龄±2岁、同性别进行1 ∶ 1配对,获得病例和对照共205对。所有研究对象均为独立个体,无血缘关系,且长期居住于苏州市郊区。吸烟定义为每日至少1支且持续≥1年;饮酒定义为每日至少50 g且持续≥1年。知情同意后由统一培训的人员进行问卷调查,按照标准方法测量并记录身高、体重、腰围、臀围。
2. PCR-SSP扩增:采集5 ml清晨空腹静脉血标本,置-80 ℃低温冰箱贮存。按试剂盒说明书进行DNA抽提(血液基因组提取试剂盒:Biomiga San Diego,CA)。16个KIR基因分别使用两组不同的引物组A和引物组B进行扩增,少数样本基因扩增结果出现引物组A和B 不一致时,使用引物组C扩增确认。同时扩增β-actin为内参。每个KIR基因扩增盘均设置阳性、阴性和空白对照。PCR-SSP扩增引物和条件见相关文献[12, 13, 14]。利用1.5%琼脂糖电泳判断PCR-SSP扩增结果的阳性和阴性。
3. 统计学分析:数据处理分析采用SAS 9.1软件。计量资料的比较采用t检验。率的比较采用χ2检验。KIR基因阳性率f通过计数测得,KIR基因频率(GF)计算公式为GF=1-。对各基因进行两两连锁不平衡检验,选择不存在连锁不平衡的KIR基因进入模型分析。采用广义多因子降维模型(GMDR)分析KIR基因-基因交互作用与原发性高血压的关联关系。运用条件logistic回归模型分析不同KIR基因型及其交互作用对原发性高血压的患病危险度。所有统计采用双侧概率检验(α=0.05),以P<0.05为差异有统计学意义。 结 果
1. 一般指标比较:病例组饮酒人数和BMI高于对照组,差异有统计学意义(P<0.05)。两组在年龄、性别、吸烟、高血压家族史分布的差异均无统计学意义(P>0.05)(表 1)。
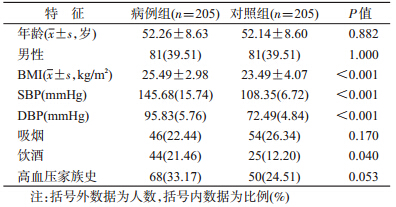
2. KIR基因频率和基因型分布的比较:用PCR-SSP方法进行KIR基因分型,检测16个KIR基因的基因频率,与对照组相比,病例组KIR2DL2、KIR2DS1、KIR2DS2、KIR2DS5的基因频率高于对照组,但差异无统计学意义(P>0.05),见表 2。病例组(6例)KIR基因型9(不携带KIR3DS1和2DS3)的携带率高于对照组,差异有统计学意义(P<0.05),其余各基因型分布的差异在两组间均无统计学意义(P>0.05)。
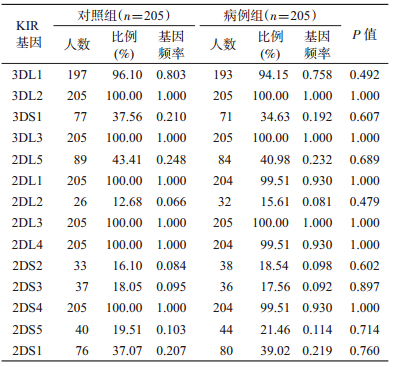
3. GMDR基因-基因交互作用模型:连锁不平衡分析显示,KIR3DS1、KIR2DS1、KIR2DS3、KIR2DS5和KIR2DL5存在连锁不平衡,KIR2DL2和2DS2,KIR3DS1和2DS1与2DS5存在连锁不平衡(P<0.05),因此选择KIR3DL1、KIR2DS1、KIR2DS2、KIR2DS3和KIR2DS5基因进入GMDR软件分析,在调整了BMI、吸烟、饮酒和高血压家族史传统危险因素后,得到与原发性高血压相关的KIR2DS2和 KIR2DS3两个基因最优模型,该模型检验精确度为55.13%,交叉验证一致性为10/10,P=0.054(表 3),提示KIR2DS2和KIR2DS3之间的基因-基因交互作用与原发性高血压患病危险可能存在相关关系。
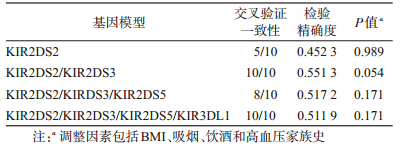
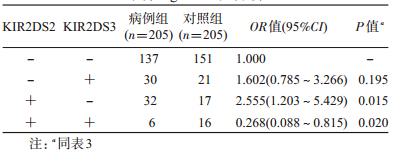
4.KIR基因多态性与原发性高血压的关联关系:无论是未调整或经多因素调整后,KIR各基因与高血压均不存在关联关系(P>0.05),KIR基因型9(不携带KIR3DS1和2DS3)与高血压也不存在显著的关联关系(OR=1.622,95%CI:0.716~3.677)。多因素条件logistic回归分析显示,在调整BMI、吸烟、饮酒和高血压家族史后,KIR2DS2阳性而KIR2DS3阴性者患原发性高血压的危险增加(OR=2.555,95%CI:1.203~5.429,P=0.015);KIR2DS2阳性且KIR2DS3阳性者患原发性高血压的危险降低(OR=0.268,95%CI:0.088~0.815,P=0.020);KIR2DS2阴性而KIR2DS3阳性者与原发性高血压不存在关联关系(OR=1.602,95%CI:0.785~3.266,P=0.195),与KIR2DS2阴性且KIR2DS3阴性者相比较(表 4),调整了BMI、吸烟、饮酒和高血压家族史后,KIR2DS2和KIR2DS3的交互作用对原发性高血压的影响存在统计学意义(OR=0.065,95%CI:0.013~0.317,P=0.001)。
原发性高血压是一种由遗传因素和环境因素共同作用导致的复杂多因素疾病,其病因至今仍不明确。目前有研究表明,自身免疫反应可促进炎症发展,而炎症和血管内皮功能异常可能在高血压发病过程中发挥作用。KIR是调节细胞免疫的重要受体,其相关基因簇构成了机体自身免疫的重要遗传学基础[15, 16, 17]。KIR主要表达于NK细胞和部分T淋巴细胞表面,其所表达的KIR活化受体和抑制受体控制该细胞的免疫活性,由于个体所携带KIR基因型的不同[18, 19],导致其所携带的转导活化信号和转导抑制信号基因的数量不尽相同,从而使得不同个体的细胞表面KIR对NK细胞和T淋巴细胞的免疫调节功能不一致,这可能与一些自身免疫性疾病有关。
已有研究表明KIR基因簇多态性在自身免疫性疾病的发病过程中发挥一定的作用。针对多发性硬化症(一种与自身免疫相关的疾病)与KIR基因多态性的研究显示[20],同时带有KIR3DS1基因型和HLA-B4的个体,患多发性硬化症的危险性显著升高(OR=2.068,95%CI:1.231~3.475,P=0.006)。另外,有研究报道[21, 22],Ⅰ型糖尿病与自身免疫反应有关并与KIR基因的多态性有关。
本研究首先比较两组人群间KIR各基因的基因频率分布,结果病例组KIR2DL2、KIR2DS1、KIR2DS2、KIR2DS5基因的阳性率高于对照组,但差异无统计学意义(P>0.05),且KIR各基因与高血压不存在关联关系。病例组KIR基因型9的携带率虽显著高于对照组,但未发现与高血压存在关联关系。高血压具有病因的多元性和复杂性,可能受到多个基因位点及环境危险因素的影响,从而产生复杂的高阶交互作用。为进一步分析KIR基因交互作用与高血压的关系,本研究根据连锁不平衡分析结果,选择KIR3DL1、KIR2DS1、KIR2DS2、KIR2DS3和KIR2DS5基因进入GMDR模型,在调整了BMI、吸烟、饮酒和原发性高血压家族史传统危险因素后,提示KIR2DS2和KIR2DS3基因之间的协同作用可能与原发性高血压患病危险存在相关关系,应用条件logistic回归模型调整BMI、吸烟、饮酒和高血压家族史后,KIR2DS2和KIR2DS3的交互作用与原发性高血压显著关联。Momot等[23]检测102例多发性硬化症患者和100例健康对照的9个KIR基因,显示多发性硬化症患者的KIR2DS2+KIR2DL2-基因型频率显著高于对照组(12% vs. 2%,P=0.005)。Yen等[11]研究发现,携带KIR2DS2基因型的个体,患银屑病并合并脉管炎的危险性显著增加(OR=5.56,P=0.001)。本研究结果显示,携带有KIR2DS2但不携带 KIR2DS3的个体患原发性高血压的危险增高,与以上研究结果一致,说明KIR2DS2是影响原发性高血压患病危险的一个重要因素。
Lu等[24]在中国山东省进行了200例肺结核患者和200例健康对照的研究,结果显示肺结核患者的KIR2DS3携带率显著高于健康对照(OR=4.60,95%CI:2.90~7.30)。Dring等[25]在爱尔兰人群进行含有543个HCV感染病例的研究发现,HCV感染者携带KIR2DS3的频率明显增高(OR=2.26,95%CI:1.30~3.94,P=0.012)。提示KIR2DS3可能在机体的自然免疫中仅发挥微弱的作用。本研究发现,同时携带有KIR2DS2和KIR2DS3的个体比均不携带的个体患原发性高血压的危险降低。KIR2DS3和KIR2DS2有相似的细胞外结构,并与配体HLA的结合位点完全相同[26],且与其他KIR相比,KIR2DS2与HLA的结合力较弱[27],KIR2DS3可能与KIR2DS2竞争性结合配体,从而减弱了KIR2DS2在自身免疫中的作用,因此降低原发性高血压的患病危险。
本研究存在不足。首先研究对象仅为单一地区的汉族人群,由于民族、地域的差异,可能在其他人群中无法重复该结果,且仅在KIR基因的低分辨分型水平做了关联研究;其次样本量尚显不足。
总之,本研究发现KIR2DS2可能是汉族人群原发性高血压的易感基因。今后有待于在不同民族、大样本人群中进一步验证,并在KIR基因高分辨分型水平进行更深入研究。
| [1] Department of Disease Control of MOH,Primary Hypertension League(China),National Cardiovascular Disease Center. Guidelines for prevention and treatment of primary hypertension in China(2010)[R]. 2010:2-3. (in Chinese)卫生部疾病控制司,原发性高血压联盟(中国),国家心血管病中心. 中国原发性高血压防治指南(2010版)[R]. 2010:2-3. |
| [2] Nicolson DJ, Dickinson HO, Campbell F, et al. Lifestyle interventions or drugs for patients with essential hypertension:a systematic review[J]. J Hypertens,2004,22(11):2043-2048. |
| [3] De Gusmäo Correia ML. Adiposity and hypertension: the inflammatory link[J]. J Hypertens,2010,28(7):1377-1379. |
| [4] Gupta V,Sachdeva S,Khan AS,et al. Endothelial dysfunction and inflammation in different stages of essential hypertension[J]. Saudi J Kidney Dis Transpl,2011,22(1):97-103. |
| [5] Pietri P,Vyssoulis G,Vlachopoulos C,et al. Relationship between low-grade inflammation and arterial stiffness in patients with essential hypertension[J]. J Hypertens,2006,24(11):2231- 2238. |
| [6] Bautista LE,Vera LM,Arenas IA,et al. Independent association between inflammatory markers(C-reactive protein,interleukin-6,and TNF-alpha) and essential hypertension[J]. J Hum Hypertens,2005,19(2):149-154. |
| [7] Cottone S,Mulè G,Nardi E,et al. Relation of C-reactive protein to oxidative stress and to endothelial activation in essential hypertension[J]. Am J Hypertens,2006,19(3):313-318. |
| [8] Sung KC,Suh JY, Kim BS, et al. High sensitivity C-reactive protein as an independent risk factor for essential hypertension[J]. Am J Hypertens,2003,16(6):429-433. |
| [9] Wang AL, Wang LF, Jiri MT, et al. Relationship between C-reactive protein and essential hypertension in Mongolian population[J]. Chin J Public Health,2010,26(4):413-415. (in Chinese)王艾丽,王凌飞,吉日木图,等. 蒙古族人群C-反应蛋白与原发性高血压关系[J]. 中国公共卫生,2010,26(4):413-415. |
| [10] Rodríguez-Iturbe B,Vaziri ND,Herrera-Acosta J,et al. Oxidative stress,renal infiltration of immune cells,and salt-sensitive hypertension:all for one and one for all[J]. Am J Physiol Renal Physiol,2004,286(4):606-616. |
| [11] Yen JH,Moore BE,Nakajima T,et al. Major histocompatibility complex class I-recognizing receptors are disease risk genes in rheumatoid arthritis[J]. J Exp Med,2001,193(10):1159-1167. |
| [12] Markus U,Peter P,Peter W. Definition of gene content for nine common group B haplotypes of the Caucasoid population:KIR haplotypes contain between seven and eleven KIR genes[J]. Immunogenetics,2002,54(4):221-229. |
| [13] Vilches C,Castaño J, Gómez-Lozano N,et al. Facilitation of KIR genotyping by a PCR-SSP method that amplifies short DNA fragments[J]. Tissue Antigens,2007,70(5):415-422. |
| [14] Ashouri E, Ghaderi A,Reed EF,et al. A novel duplex SSP-PCR typing method for KIR gene profiling[J]. Tissue Antigens,2009,74(1):62-67. |
| [15] Kulkarni S,Martin MP,Carrington M. The Yin and Yang of HLA and KIR in human disease[J]. Semin Immunol,2008,20(6):343-352. |
| [16] Rajalingam R. Human diversity of killer cell immunoglobulin- like receptors and disease[J]. Korean J Hematol,2011,46(4):216-228. |
| [17] Parham P. The genetic and evolutionary balances in human NK cell receptor diversity[J]. Semin Immunol,2008,20(6):311-316. |
| [18] Makrigiannis AP,Parham P. The evolution of NK cell diversity[J]. Semin Immunol,2008,20(6):309-310. |
| [19] Gardiner CM. Killer cell immunoglobulin-like receptors on NK cells:the how,where and why[J]. Int J Immunogenet,2008,35(1):1-8. |
| [20] García-León JA,Pinto-Medel MJ,García-Trujillo L,et al. Killer cell immunoglobulin-like receptor genes in Spanish multiple sclerosis patients[J]. Mol Immunol,2011,48(15/16):1896- 1902. |
| [21] Åkesson C,Uvebrant K,Oderup C, et al. Altered natural killer (NK) cell frequency and phenotype in latent autoimmune diabetes in adults (LADA) prior to insulin deficiency[J]. Clin Exp Immunol,2010,161(1):48-56. |
| [22] Ramos-Lopez E,Scholten F,Aminkeng F,et al. Association of KIR2DL2 polymorphism rs2756923 with type 1 diabetes and preliminary evidence for lack of inhibition through HLA-C1 ligand binding[J]. Tissue Antigens,2009,73(6):599-603. |
| [23] Momot T,Koch S,Hunzelmann N,et al. Association of killer cell immunoglobulin-like receptors with scleroderma[J]. Arthritis Rheumatol,2004,50(5):1561-1565. |
| [24] Lu C,Bai XL,Shen YJ,et al. Potential implication of activating killer cell immunoglobulin-like receptor and HLA in onset of pulmonary tuberculosis[J]. Scand J Immunol,2012,76(5):491-496. |
| [25] Dring MM, Morrison MH, McSharry BP,et al. Innate immune genes synergize to predict increased risk of chronic disease in hepatitis C virus infection[J]. Proc Natl Acad Sci USA,2011,108(14):5736-5741. |
| [26] Campbell KS,Purdy AK. Structure/function of human killer cell immunoglobulin-like receptors:lessons from polymorphisms,evolution,crystal structures and mutations[J]. Immunology,2011,132(3):315-325. |
| [27] Stewart CA, Laugier-Anfossi F, Vély F,et al. Recognition of peptide-MHC classⅠ complexes by activating killer immunoglobulin-like receptors[J]. Proc Natl Acad Sci USA,2005,102(37):13224-13229. |