 2015, Vol. 36
2015, Vol. 36文章信息
- 魏柏青, 熊浩明, 杨晓艳, 杨永海, 祁美英, 靳娟, 辛有全, 李翔, 杨汉青, 韩秀敏, 代瑞霞. 2014.
- Wei Baiqing, Xiong Haoming, Yang Xiaoyan, Yang Yonghai, Qi Meiying, Jin Juan, Xin Youquan, Li Xiang, Yang Hanqing, Han Xiumin, Dai Ruixia. 2014.
- 青海高原藏系绵羊鼠疫流行病学和病原学特征分析
- The epidemiology and etiology research of Tibetan sheep plague in Qinghai plateau
- 中华流行病学杂志, 2015, 36(3): 271-274
- Chinese Journal of Epidemiology, 2015, 36(3): 271-274
- http://dx.doi.org/10.3760/cma.j.issn.0254-6450.2015.03.017
-
文章历史
- 投稿日期:2014-09-19




青海省自1954年证实为鼠疫自然疫源地,现存在两种类型的鼠疫自然疫源地分布于全省33个县(市),疫源地面积近20万km2,迄今共发现喜马拉雅旱獭(Marmota himalayana)、藏系绵羊(Tibetan sheep,藏羊)等19种染疫动物、12种染疫昆虫。1975年在玉树县上拉秀地区证实藏羊能自然感染鼠疫并成为人类鼠疫的传染源[1]。目前为止,青海省由藏羊作为传染源引起人间鼠疫共10起,发病25例,死亡13例。1990-2009年青海省喜马拉雅旱獭疫源地鼠疫病例因剥食藏羊而感染约占13%,因此藏羊鼠疫存在的风险不容忽视。为此本研究从生态流行病学角度分析青海高原藏羊鼠疫病原学和流行病学特征及其地理分布、传播生态学因素等,为制订适合青海省鼠疫预防控制策略和措施提供科学依据。 资料与方法
1. 资料和菌株来源:藏羊鼠疫背景资料由青海省鼠疫防治信息资源库提供。1975-2009年从青海省鼠疫藏羊分离的14株鼠疫菌由青海省国家鼠疫菌种保藏中心提供。
2. 流行病学分析方法:对1975-2009年青海省藏羊鼠疫背景资料进行汇总,分析地区、时间、人间分布及感染途径等。人间鼠疫病例和鼠疫菌结果判定参照《鼠疫诊断标准》(WS 279-2008)及附录B。采用青海省地方病预防控制所地理信息系统(GIS)进行青海高原藏羊鼠疫空间分布特征分析[2]。
3. 实验室检测:
(1)毒力测定:菌株毒力测定按常规方法,计算最小致死量(MLD),并参照朱锦沁等[3]方法对毒力等级分类。实验动物为490只昆明系小白鼠,清洁级,体质量18~20 g,由青海省实验动物中心提供。
(2)细菌表型特征:按常规方法进行鼠疫菌生化、糖醇类酵解试验、毒力因子等表型特征分析,根据传统的鼠疫菌生态型6项指标分析鼠疫菌生态型[4]。
(3)质粒分析和DNA提取:参照文献[5]方法分析质粒,质粒DNA相对分子质量(Mr)的测定采用标准质粒对照法。鼠疫菌DNA提取按照经典的苯酚-氯仿混合抽提法[6]。
(4)细菌差异片段(DFR)分型:用23对DFR(DFR01~DFR23)分型引物和PMT1(质粒验证引物),同时检索中国鼠疫菌DFR分型数据库[6,7],对被试菌株DNA进行分型验证,以91001、620024菌株DNA的等量混合物为阳性对照,同时设置阴性对照,引物序列及结果分析参见文献[6,7]。 结果
1. 藏羊鼠疫流行病学特征:
(1)藏羊动物间鼠疫:1975-2009年青海省从藏羊体内共分离14株鼠疫菌。其中分离自玉树县10株、囊谦县2株、治多县1株、德令哈市1株(图 1);1975-1979年分离4株,1996-1998年分离7株,2003-2009年分离3株;以11月分离鼠疫菌株数最多(6株)。藏羊体表寄生物主要为蠕形蚤科蠕形蚤属和长喙蚤属蚤及草原硬蜱等,均未分离出鼠疫菌。
(2)染疫藏羊作为传染源的人间鼠疫:1975-2009年由藏羊作为传染源引起人间鼠疫10起,发病25例,死亡13例。其中腺鼠疫9例,肺鼠疫6例,败血型鼠疫3例,肠鼠疫1例,由肠鼠疫、腺鼠疫继发为肺鼠疫、败血型鼠疫6例。25例中24例为藏族牧民,1例为汉族农民,其中男性11例,女性14例;年龄9~72岁,其中<20岁9例,20~45岁8例,>45岁为8例。人间鼠疫全部发生在青海省南部江河源-甘南生态区的玉树、玛多、囊谦、杂多和玛沁县(图 1);首发病例均为剥食鼠疫病死的藏羊,其次为接触鼠疫病例而感染;鼠疫病例均发生在6-11月,其中3起发生在旱獭入蛰后的10-11月,以11月发病人数最多(11例),约占鼠疫总病例的44%(11/25)。
 |
| 注:▲共分离14株鼠疫菌(德令哈市1996年分离1株,治多县2003年分离1株,玉树县1975、1979、1997、1998、2005年分别分离2、2、3、1、2株,囊谦县1997年分离2株); ■共发生10起人间鼠疫(玛多县发生2起,其中1977年11月发病4例、死亡2例,1982年6月发病2例、死亡2例;玛沁县发生1起,1983年9月发病1例、死亡0例;杂多县发生2起,其中1988年10月发病2例、死亡2例,1989年6月发病2例、死亡2例;玉树县发生3起,其中1975年8月发病2例、死亡1例,1979年9月发病3例、死亡2例,1980年7月发病、死亡各1例;囊谦县发生2起,其中1979年6月发病、死亡各1例,1997年11月发病7例,无死亡) 图 1 1975-2009年青海高原藏羊鼠疫的空间分布 |
2. 藏羊鼠疫菌病原学特征:
(1)生化特性:14株菌均能分解阿胶糖、麦芽糖和甘油,不分解鼠李糖和密二糖,脱氮阳性,生态型为青藏高原型。
(2)毒力因子试验:14株菌均产生荚膜抗原(F1)和鼠疫杆菌素Ⅰ(PstⅠ);11株菌能产生毒力抗原因子(VW),3株则不产生;11株菌色素沉着因子(Pgm)为“+”,1株为“±”,2株为“-”。
(3)毒力测定:14株菌小白鼠毒力测定,其中MLD为10个菌的有5株,50个菌的有3株,100个菌的有4株,1 000个菌的有2株,均为强毒株。
(4)质粒谱:14株菌均携带有3种质粒,Mr值分别为6×106、45×106、65×106或52×106;在质粒组成上又分别有2种不同的组合形式,其Mr值分别为6×106、45×106、65×106和6×106、45×106、52×106,其中具有分类意义的大质粒有65×106和52×106。玉树、囊谦、治多县分离的菌株携带65×106大质粒;而德令哈市分离的菌株携带52×106大质粒。
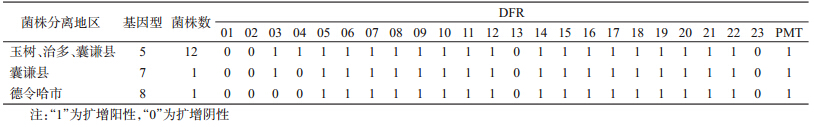
(5)基因型:检索中国鼠疫菌DFR分型数据库,23个DFR和PMT1进行PCR扩增后显示,玉树、治多县分离的菌株均为5型,囊谦县分离的菌株分别为5型(1株)和7型(1株),德令哈市分离的菌株为8型(表 1)。
藏羊广泛分布于青藏高原鼠疫自然疫源地。在自然条件下,藏羊为摄取钙、磷和盐等无机物,有舔食动物尸体和尸骨的习性。资料显示[1],在以喜马拉雅旱獭为主要储存宿主的青海鼠疫疫源地内,藏羊鼠疫来自早獭鼠疫,其感染途径主要有经口和媒介疫蚤(谢氏山蚤和斧形盖蚤)叮咬两种感染方式。本研究显示,1975-2009年青海省分离自藏羊鼠疫的鼠疫菌株生态型为青藏高原型;78.57%(11/14)的鼠疫菌具有4个毒力因子,且均为强毒菌;大质粒分布及基因型也显示均具备青藏高原鼠疫病原体特性,表明人感染藏羊鼠疫后同样具有发病急、病情重、传染性强、病死率高等特点,对人类有极其严重威胁。
青海省从1958年确诊第1例人间鼠疫病例至2011年为止,全省共发生人间鼠疫198起,由藏羊引起有10起(5.03%),自藏羊分离出鼠疫菌14株。无论是从鼠疫菌的分离数量还是作为传染源引发人间鼠疫病例,藏羊的流行病学意义仅次于旱獭而位于第二。目前,青海藏羊鼠疫流行地区除德令哈市外,全部分布在南部江河源-甘南生态区的玉树、囊谦、杂多等县,这些地区历年来旱獭鼠疫流行猛烈,有其独特的地理景观、地形地貌、植被、生物群落及气候特点,鼠疫菌的主要宿主及传播媒介密度较高,数量稳定,具有鼠疫菌生存及延续的完整生态系统。本研究还证实,藏羊引发的人间鼠疫,在6-11月均有流行,与藏羊动物鼠疫流行时间、地区近乎一致。旱獭入蛰后的11月自藏羊体内分离的鼠疫菌株数及其作为传染源引发的人间鼠疫病例数最多。资料显示[1],在以喜马拉雅旱獭为主要储存宿主的青海鼠疫疫源地内,藏羊对鼠疫菌属高感受中等敏感动物,为该疫源地的偶然宿主,藏羊感染鼠疫菌的潜伏期多为1~2 d,偶有5 d。实验室感染的羊只其病程68.35%为迁延型,其中个别病羊可带菌达1个月之久,急型和亚急型仅占31.6%,58.96%的病羊可自愈。因此染疫藏羊在旱獭入蛰后仍可作为人类鼠疫传染源而引起人间鼠疫,构成了青海高原藏羊鼠疫在流行时间上滞后的特殊现象。就人群感染途径而言,藏羊人间鼠疫的发生主要取决于主动接触、剥食病死鼠疫的藏羊,并以腺鼠疫为首发病例,由此继发为肺鼠疫、败血型鼠疫病例多且病亡率高。
青海鼠疫自然疫源地是全国甚至是全球鼠疫高发地区,该疫源地由于受多种生态地理环境要素的影响,形成了鼠疫生态地理景观的多态性,造就了鼠疫自然疫源地长期存在且复杂而活跃。青海省藏羊数量约占我国绵羊总数的1/3,与喜马拉雅旱獭的自然分布区基本重叠[8]。1975年首次从藏羊体内检测到鼠疫菌以来,藏羊间鼠疫流行不断,局部地区牧场还发生鼠疫爆发性流行[9]。藏羊参与鼠疫流行为世界罕见,也是青藏高原鼠疫流行的独特现象。藏羊参与鼠疫流行使青海鼠疫流行演变出许多新的特点,如流行周期延长,传播途径更复杂,易感人群基数增加,对人类的危害更加严重。另外,由于藏羊是当地牧民赖以生存的家畜,人类与之接触频繁密切,且随交通条件的改善和发展,每年还有相当数量的未经深加工的藏羊畜产品(例如生肉、皮张等)运往全国各地,且近年来还陆续从西藏山羊、藏原羚、岩羊、牦牛等偶蹄类动物体内分离出鼠疫菌或检测出鼠疫F1抗体[10]。因此藏羊鼠疫在鼠疫流行病学上的意义和地位将更加突出。
| [1] Wang WH,Li C. The study on plague of Tibetan sheep[J]. Chin J Endemiol,2001,20(6):467-468. (in Chinese) 王文华,李超. 藏系绵羊鼠疫的调查研究[J]. 中国地方病学杂志,2001,20(6):467-468. |
| [2] Tang XY,Wang GJ,Li Y,et al. The establishment of comprehensive geographic information system for kinds of endemic diseases in Qinghai province[J]. Chin J Endemiol,2010,29(6):687-689. (in Chinese) 唐新元,王国钧,李勇,等. 青海省地方病多病种综合地理信息系统的建立[J]. 中国地方病学杂志,2010,29(6):687-689. |
| [3] Zhu JQ, Li M,Wang L,et al. Study on biological features and epidemiological significance of Yersinia pestis in Qinghai plague natural focus[J]. Endem Dis Bull,1994,9(1):1-5. (in Chinese) 朱锦沁,李敏,王丽,等. 青海省鼠疫疫源地鼠疫菌某些生物学特性及其流行病学意义的研究[J]. 地方病通报,1994,9(1):1-5. |
| [4] Dai RX,Wei BQ,Li CX,et al. The pathogenic ecology research on plague in Qinghai plateau[J]. Chin J Prev Med,2013,47(12):1083-1088. (in Chinese) 代瑞霞,魏柏青,李存香,等. 青海高原鼠疫病原生态学研究[J]. 中华预防医学杂志,2013,47(12):1083-1088. |
| [5] Li M, Yu XT,Li L,et al. Screening of plasmids in Yersinia pestis strains isolated from China[J]. Chin J Microbiol Immunol,1995,15(5):341-343. (in Chinese) 李敏,于晓涛,黎莉,等. 我国鼠疫菌质粒种类和组成特征的研究[J]. 中华微生物学和免疫学杂志,1995,15(5):341-343. |
| [6] Li Y,Dai E,Cui Y,et al. Different region analysis for genotyping Yersinia pestis isolates from China[J]. PLoS One,2008,3(5):e2166. |
| [7] Yang XY,Wei BQ,Jin J,et al. Regional genotyping and the geographical distribution regarding Yersinia pestis isolates in China[J]. Chin J Epidemiol,2014,35(8):943-948. (in Chinese) 杨晓艳,魏柏青,靳娟,等. 中国鼠疫耶尔森菌差异区段分型及其地理分布特征[J]. 中华流行病学杂志,2014,35(8):943-948. |
| [8] Wang ZY. The research of Tibetan sheep plague in Qinghai province and the prevention and control countermeasures[J]. Chin J Zoonoses,1999,15(6):105-106. (in Chinese) 王祖郧. 青海省藏系绵羊鼠疫及其防治对策[J]. 中国人兽共患病杂志,1999,15(6):105-106. |
| [9] Yu XT,Cui BZ,Zhao XL,et al. A Tibetan sheep plague outbreak investigation in Guoqing Ranch,Yushu county,Qinghai province[J]. Chin J Epidemiol,2006,27(12):1060. (in Chinese) 于晓涛,崔百忠,赵小龙,等. 青海省玉树县果青牧场藏系绵羊鼠疫疫情调查[J]. 中华流行病学杂志,2006,27(12):1060. |
| [10] Wu KM,Li C,Wang YZ,et al. A retrospective analysis of plague in Three-River Source Region of Qinghai from 1954 to 2006[J]. Chin J Endemiol,2008,27(6):647-650. (in Chinese) 吴克梅,李超,汪元忠,等. 1954-2006年青海省三江源地区鼠疫及其流行病学分析[J]. 中国地方病学杂志,2008,27(6):647-650. |