 2022, Vol. 33
2022, Vol. 33 

b Department of Chemistry, University at Buffalo, The State University of New York, Buffalo, NY 14260, United States;
c Department of Chemistry, Hofstra University, Hempstead, NY 11549, United States;
d Academy of Chinese Medical Science, Henan University of Chinese Medicine, Zhengzhou 450046, China;
e Chengdu Institute of Organic Chemistry, Chinese Academy of Sciences, Chengdu 610041, China
As a major class of protein secondary structures, reverse turns are sites of chain reversal responsible for the globular character of a protein [1,2]. β-Turns, i.e., tight turns consisting of two amino acid residues, are frequently found in hairpin loops of globular proteins [3-5]. The design of type Ⅱ' d-Pro-Gly β-turn [6-9], type Ⅰ' Asn-Gly [10-12], Aib-d-Ala [13] β-turns, and other such as aza-amino acids [14] have facilitated the formation and stabilization of isolated β-hairpins consisting of α-amino acids, and have played crucial roles in initiating the folding of peptides and proteins [15,16]. Stable β-turns also exhibit important biological functions such as serving as epitopes in protein–protein [17-20] and protein–nucleic acid [21-25] interactions.
Interests in developing foldamers have led to the creation of many folding oligomers containing unnatural building blocks that adopt defined conformations including sheets and turns [26-30]. For example, aromatic residues derived from 5-amino-2-methoxybenzoic acid were incorporated into β-strands by Nowick to enhance the stability of templated and cyclized β-sheets [31]. β-Turns based on nipecotic acid developed and those containing β−2, 3-amino acid residues reported by Seebach [32,33] were shown to facilitate the formation of β-hairpin structures of β-peptides. Besides, β-hairpins containing centrally located α/γ-and α/δ-hybrid dipeptide loops that connect oligopeptide strands of α-amino acid residues were also reported [34].
We created oligoamide strands consisting of alternating meta-linked aromatic units and α-amino acid residues [35]. Such oligoamides strands, as mimics of β-strands, associate into double-stranded sheets with stabilities proportional to the number of inter-strand H-bonds [36-38]. A recent attempt to enhance the diversity of our H-bonded duplexes, based on the incorporation of glycine and β-alanine, the most flexible α- and β-amino acid residues, into the oligoamide strands, led to hybrid peptide 1 (Fig. 1a) that was found to adopt a stably folded hairpin, i.e., an expanded β-turn, containing a β-Ala-Gly-dipeptide loop flanked by two doubly H-bonded aromatic γ-amino acid residues [39]. Subsequent studies revealed that such an expanded β-turn motif is capable of accommodating a variety of different combinations of α/β-dipeptide sequences [40,41].

|
Download:
|
| Fig. 1. (a) Hybrid peptide 1 shown as its expanded β-turn conformation. (b) Hybrid peptides 2a-2e shown as their likely β-turn conformations. The amino acid residues are labeled in red. The selected hydrogens are labeled in blue. | |
To probe whether the β-turn motif adopted by 1 and its analogous hybrid peptides could be extended to constructing two-residue loops consisting of α-amino acid residues, four-residue hybrid peptides 2, which differ in the α-amino acid sequences of their central dipeptide segments, are designed (Fig. 1b). Each of peptides 2, if folding into the conformation as shown, will give a β-hairpin consisting of, from the N- to the C-termini, four consecutive residues with position i being an aromatic γ-amino acid, positions i + 1 and i + 2 being two α-amino acid, and i + 3 being another aromatic γ-amino acid, residues. By studying the folding of 2, we hope to address the following questions: Could such a β-turn motif accommodate different α-amino acid residues? How do different α-amino acid residues influence the stability of the corresponding β-turns?
Herein we report that the folding of peptides 2a-2e, with residues i + 1 and i + 2 being Gly-Gly, l-Ala-Gly, l-Phe-Gly, l-Ala-l-Ala and d-Pro-Gly, follows a general pattern by adopting β-hairpin conformations as shown by results from 1D and 2D 1H NMR studies and confirmed by the crystal structures of 2a-2d. The stabilities of the hairpin conformations are found to vary with the α-amino acid sequences of the dipeptide loops.
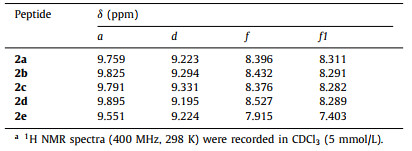
The 1H NMR spectra of 2a-2e from 0.1 mmol/L to 25 mmol/L in CDCl3 reveal sharp peaks, except for proton a of 2e (Fig. S1 in Supporting information). The well dispersed 1H resonances of these hybrid peptides indicate that, although having multiple H-bond donors and acceptors, peptides 2a-2e do not engage in noticeable self-aggregation and most likely exist as discrete species adopting defined conformations in solution. Consistent with the adoption of β-turn conformations by 2a-2e, amide protons a, b, d and e show insignificant changes in their chemical shifts, indicating their involvement in intramolecular H-bonding; in contrast, protons c of all five peptides show much larger downfield shifts, suggesting their engagement in intermolecular H-bonding.
Comparing the chemical shifts of amide protons a and d of 2a-2d (Table 1) at 5 mmol/L provides for a qualitative assessment on the relative stabilities of the corresponding H-bonds and thus those of the β-turns. Compared to that of 2a, the signals of proton d of 2b and 2c move downfield, which imply that the H-bonded loops of 2b and 2c are stabilized due to the presence of the l-Ala-and l-Phe-side chains; the signal of proton d of 2d moves slightly upfield, suggesting that the two l-Ala-residues more or less twist the H-bonded loop and weakens the H-bond involving proton d.
|
|
Table 1 Chemical shifts of protons a, d, f, and f1 in CDCl3.a |
{kind=link}
The chemical shifts of protons a which are away from the H-bonded loops indicate the strength of the H-bonds and also indicate the stability of the β-turns. Relative to that of 2a, the signals of protons a of 2b-2d move downfield. The resonance of 2d shows the largest downfield shift, followed by that of 2b, and then by 2c, based on a stability order of 2d > 2b > 2c > 2a results. Thus, the presence of α-amino acid sidechains in hybrid peptides 2b-2d seems to enhance the stability of the hairpins, most likely by imposing stereochemical constraints that limit the conformational flexibility of the turn loops.
Compared to the sharp resonances of protons a of 2a-2d, proton a of 2e, which contains a d-Pro-Gly-loop that strongly promotes turn and hairpin conformation in peptides of α-amino acids [6], gives a broadened peak (Fig. S1e) that moves upfield to 9.551 ppm (Table 1). Besides, the peaks of protons f and f1 of 2e also move upfield to 7.915 and 7.403 ppm, respectively, relative to those of protons f and f1 of 2a-2d (Table 1). Thus, the H-bond involving proton a of 2e seems to be weakened, resulting in a conformation of 2e in which the two aromatic residues and the N- and C-termini undergo dynamic relative motion.
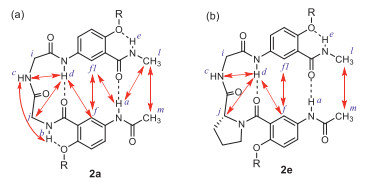
Peptides 2a-2e were then examined with 2D NMR (NOESY) in CDCl3 (Fig. S2 in Supporting information). The NOEs observed with 2a (Fig. 2a) are those between protons a and l, a and f1, d and f, and f and f1, indicating that the two aromatic rings are brought into close proximity by H-bonding involving amide protons a and d. Besides, the NOE between protons l and m demonstrates that the two termini of 2a are in close proximity, which is consistent with the expected hairpin conformation. NOEs involving the protons of the Gly-Gly-segment of 2a include those between protons c and b, c and d, and d and j, which indicate the presence of a well-defined loop. The NOESY spectra of 2b-2d (Figs. S2b-d) reveal that NOEs share the same overall pattern observed in that of 2a, suggesting that 2b-2d also adopt well-defined β-turn conformations in CDCl3.

|
Download:
|
| Fig. 2. NOEs shown as double-headed arrows, between remote (non-adjacent) protons revealed by the NOESY spectra of (a) 2a and (b) 2e in CDCl3 (5 mmol/L, 298 K, 400 MHz, mixing time 0.3 s). | |
{kind=link}
The NOEs revealed for 2e (Fig. 2b) include those between protons c and d, d and j, d and f, f and f1, and l and m, indicating that 2e also adopts a β-turn conformation. That no NOE between protons a and f1, or a and l is observed, along with the significant upfield shift of this proton resonance relative to the signals of protons a of 2a-2d, is consistent with the weakening of the H-bond involving proton a of 2e.
Single crystals of 2a-2d were obtained, which led to the determination of the X-ray structures of these hybrid peptides. As shown in Fig. 3, each of the four hybrid peptides, regardless of its two-residue sequence involving residues i + 1 and i + 2, folds into a typical β-hairpin conformation that is held together by two intramolecular H-bonds involving amide protons a and d of residues i and i + 3. Although the two-residue sequences Gly-Gly, l-Ala-Gly, l-Phe-Gly and l-Ala-l-Ala-do not have high β-turn propensities [4,5], peptides 2a-2d nevertheless fold into β-hairpins, which demonstrate the general applicability of this β-turn motif for accommodating different loop sequences.

|
Download:
|
| Fig. 3. Crystal structures of (a) 2a, (b) 2b, (c) 2c, and (d) 2d Two independent molecules with their main chain atoms adopting enantiomeric turn types having are found for 2a, 2c, or 2d For clarity, all hydrogen atoms except for those of the amide groups, are removed. | |
{kind=link}
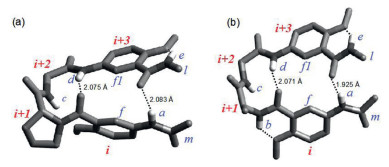
Based on the crystal structures of 2a-2d, dihedral angles φ and ψ of residues i + 1 and i + 2 in the turns are obtained, which reveal the β-turn types (Fig. 3). The crystal structure of achiral 2a (CCDC: 2, 053, 064) reveals two independent molecules adopting the enantiomeric type Ⅰ and type Ⅰ' β-turn conformations (Fig. 3a). With its chiral l-Ala-residue, peptide 2b (CCDC: 2, 053, 065) exists as two independent molecules with very similar φ and ψ angles and folds into a type Ⅱ β-turn (Fig. 3b). Interestingly, peptides 2c (CCDC: 2053066) (Fig. 3c) and 2d (CCDC: 2053067) (Fig. 3d), although having one (L-Phe) and two (L-Ala) chiral residues, respectively, still fold into both types Ⅰ and I' β-turns, i.e., the presence of chiral α-amino acid residues is incapable of biasing the main chain atoms of 2c or 2d to settle on one of the two enantiomeric turn types.
Attempts to grow single crystals of peptide 2e have been unsuccessful. The conformation of 2e was computationally probed using the ADF and VASP software packages with revPBE-D3 [42-44]. The most stable conformation of 2e is a β-hairpin in which the planes of the two aromatic residues have a sharp dihedral angle of ~60° (Fig. 4a). Besides, the benzene ring of residue i and the amide group linking residues i and i + 1 deviate from coplanarity, with a dihedral angle of ~50°. In contrast, the optimized conformation of 2b shows a much less curved structure, with a dihedral angle of ~117° between the benzene rings of the two aromatic residues (Fig. 4b). The benzene ring of residue i and the amide group between residues i and i + 1 are coplanar. The twisted conformation of 2e seems to be caused by the need to avoid steric hindrance between the five-membered ring of the Pro-residue and the adjacent methoxy sidechain of aromatic residue i. As a result, the two H-bonds involving protons a and d are slightly longer, and thus weaker, in 2e than those in 2b.

|
Download:
|
| Fig. 4. Energy-minimized structures of the hairpin conformations of (a) 2e and (b) 2b. The isopentyloxy side chains are replaced with methoxy groups. For clarity, all hydrogen atoms except for those of the amide groups, are removed. | |
{kind=link}
Quantum molecular dynamics calculations on 2b and 2e in a NVT ensemble (298 K, Supporting information for movies) revealed that the length of the H-bond involving proton a of 2b fluctuated in a narrow range (~1.7–2.5 Å), while the H-bond of 2e involving proton a was drastically more dynamic and fluctuated over a much larger range (~1.7–3.8 Å) (Fig. S7 in Supporting information). Thus, structural deformation caused by the d-Pro-residue results in weakened H-bonding interactions and thus lowered stability for the hairpin of 2e.
The stabilities of the folded structures of 2a-2e were further examined by comparing the effects of temperature and solvent polarity on the strength of H-bonds involving protons a and d (Figs. S3 and S4, Table S1 in Supporting information). As shown in Fig. 5a, from 0 to 45 ℃, the signals of protons a and d all move upfield. The signal of proton a of 2e shows an upfield shift (0.43 ppm) that is significantly larger than those of the other four peptides, and those of protons a of 2a-2d shift within a much narrower range of 0.06 to 0.14 ppm. Among protons d, that of 2e shows the largest upfield shift (0.21 ppm), while those of 2a-2d are much smaller, ranging from 0.05 to 0.10 ppm. The temperature-dependent upfield shifts of amide protons a and d indicate that, the presence of the d-Pro-residue in 2e weakens the H-bonds involving protons a and d much more significantly than in 2a-2d. Among 2a-2d, the presence of side chains in the β-turn loops of 2b-2d slightly weakens the H-bonds involving protons a, while enhancing the H-bonds involving protons d.

|
Download:
|
| Fig. 5. Changes in chemical shifts (Δδ) amide protons a and d (400 MHz) of 2a-d (5 mmol/L) (a) from 0—45 ℃ in CDCl3 (Δδ = δ(45 ℃) – δ(0 ℃)) and (b) in mixed solvents containing 0 to 20% DMSO-d6 in CDCl3 at 25 ℃ (Δδ = δ(20% DMSO) – δ(0% DMSO)). | |
{kind=link}
In mixed solvents containing 0 to 20% DMSO-d6 in CDCl3 (Fig. 5b), the signals of both protons a and d of 2a-2e show downfield shifts. The shifts of protons a and d of 2e, being 0.12 and 0.19 ppm, respectively, are much larger than those of the other four peptides, confirming that the protons a and d are more exposed in 2e, i.e., the corresponding H-bonds are less shielded, leading to a confirmation of 2e that is less stable than those of 2a-2d. The downfield shifts of protons a and d of 2a are larger than those of protons a of 2b-2d, indicating that the α-amino acid side chains of 2b-2d enhance the stabilities of these three β-turns, a conclusion that is consistent with the one based on comparing the chemical shift values of protons a.
The critical role played by the aromatic γ-amino acid residues in promoting the folding of peptides 2 was demonstrated by comparing 3a-3c which differ in their central dipeptide sequences but share the same l-Val-and l-Leu-terminal residues (Fig. 6a). Peptide 3c is known to fold into a β-hairpin conformation that is promoted by its d-Pro-Gly-loop sequence [6]. As shown in Fig. S5 and Table S2 (Supporting information), from 0.1 mmol/L to 25 mmol/L in CDCl3, the signals of amide protons b, c, and d of 3a show noticeable downfield shifts of ~0.56 to 0.77 ppm and that of proton a shows a very small shift (~0.02 ppm); while all four amide protons of 3b undergo very large downfield shifts from ~1.61 to 1.74 ppm. Consistent with the folding of 3c into a β-hairpin, the downfield shifts of amide protons a and d of 3c, being 0.13 and 0.05 ppm, are much smaller than those of 3a and 3b (except for proton a of 3a), which also suggests that 3a and 3b undergo intermolecular H-bonding interaction and do not fold like 3c.

|
Download:
|
| Fig. 6. (a) Tetrapeptides 3a-c differing in their central residues (highlighted in red-dashed box) flanked by the same l-Val-and l-Leu-residues. (b) NOEs (red double-headed arrows) between remote protons revealed by the NOESY spectrum of 3c (5 mmol/L in CDCl3, 400 MHz, mixing time: 0.3 s). | |
{kind=link}
The NOESY spectrum of 3c (Fig. S6 in Supporting information) reveals NOEs between remote protons a and d, d and i, and e and f. In contrast, no NOEs between the corresponding remote protons, especially between protons a and d, and e and f, are found in the NOESY spectra of 3a and 3b, showing that 3a and 3b, unlike 3c, do not fold into β-hairpin conformations.
In summary, hybrid tetrapeptides 2a-2e, each has a central dipeptide segment of α-amino acid residues and two aromatic γ-amino acid residues, fold into well-defined β-turn hairpins. Results from 1D and 2D NMR, and computational studies, along with crystal structures, demonstrate that the five different two-residue sequences of α-amino acid residues are all incorporated as the turn loops of the β-hairpins. d-Pro-Gly, a turn sequence that strongly promotes the adoption of β-hairpin conformations by peptides of α-amino acids, results in a β-turn with the lowest stability. Compared to peptide 2a which has a Gly-Gly-loop, peptides 2b-2d with l-Ala-Gly, l-Phe-Gly and l-Ala-l-Ala-loops, showed enhanced stabilities. The inability of peptides 3a and 3b, which shares their central dipeptide segments with 2a and 2d, respectively, demonstrates the critical importance of the two aromatic γ-amino acid residues in ensuring the adoption of hairpin conformations by peptides 2a-2e. The same folding pattern of the five hybrid peptides has offered a reliable hairpin motif capable of accommodating different two-residue sequences of α-amino acids, which will greatly enhance the diversity of β-turns and β-hairpins.
Declaration of competing interestThe authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
AcknowledgmentsThis work was supported by the National Natural Science Foundation of China (No. 21778012 to Z.L. Lu, 21801020 to R. Liu), the American Chemical Society – Petroleum Research Fund (PRF# 58364-ND7, to B. Gong), and the Center for Computational Research (CCR) (to D.P. Miller and E. Zurek), and Hofstra University (to D.P. Miller).
Supplementary materialsSupplementary material associated with this article can be found, in the online version, at doi:10.1016/j.cclet.2021.06.019.
| [1] |
P.N. Lewis, F.A. Momany, H.A. Scheraga, BBA-Protein Struct 303 (1973) 211-229. DOI:10.1016/0005-2795(73)90350-4 |
| [2] |
G.D. Rose, L.M. Glerasch, J.A. Smith, Adv. Protein Chem. 37 (1985) 1-109. DOI:10.1039/d0sc03060k |
| [3] |
C.M. Wilmot, J.M. Thornton, J. Mol. Biol. 203 (1988) 221-232. DOI:10.1016/0022-2836(88)90103-9 |
| [4] |
E.G. Hutchinson, J.M. Thornton, Protein Sci. 3 (1994) 2207-2216. DOI:10.1002/pro.5560031206 |
| [5] |
B. Madan, S.Y. Seo, S.G. Lee, Proteins 82 (2014) 1721-1733. DOI:10.1002/prot.24526 |
| [6] |
T.S. Haque, J.S. Little, S.H. Gellman, J. Am. Chem. Soc. 118 (1996) 6975-6985. DOI:10.1021/ja960429j |
| [7] |
T.S. Haque, S.H. Gellman, J. Am. Chem. Soc. 119 (1997) 2303-2304. DOI:10.1021/ja963653h |
| [8] |
J.F. Espinosa, S.H. Gellman, Angew. Chem. Int. Ed. 39 (2000) 2330-2333. DOI:10.1002/1521-3773(20000703)39:13<2330::AID-ANIE2330>3.0.CO;2-C |
| [9] |
F.A. Syud, H.E. Stanger, S.H. Gellman, J. Am. Chem. Soc. 123 (2001) 8667-8677. DOI:10.1021/ja0109803 |
| [10] |
E. de Alba, M.A. Jiménez, M. Rico, J. Am. Chem. Soc. 119 (1997) 175-183. DOI:10.1021/ja962325e |
| [11] |
A.J. Maynard, M.S. Searle, Chem. Commun. (1997) 1297-1298. DOI:10.1007/978-1-4939-1486-9_2 |
| [12] |
E.R. Simpson, J.K. Meldrum, R. Bofill, et al., Angew. Chem. Int. Ed. 44 (2005) 4939-4944. DOI:10.1002/anie.200500577 |
| [13] |
S. Aravinda, N. Shamala, R. Rajkishore, et al., Angew. Chem. Int. Ed. Engl. 41 (2002) 3863-3865. DOI:10.1002/1521-3773(20021018)41:20<3863::AID-ANIE3863>3.0.CO;2-A |
| [14] |
N. Tonali, I. Correia, J. Lesma, et al., Org. Biomol. Chem. 18 (2020) 3452-3458. DOI:10.1039/C9OB02654A |
| [15] |
A.M.C. Marcelino, L.M. Gierasch, Biopolymers 89 (2008) 380-391. DOI:10.1002/bip.20960 |
| [16] |
D. Du, Y. Zhu, C.Y. Huang, et al., Proc. Natl. Acad. Sci. U. S. A. 101 (2004) 15915-15920. DOI:10.1073/pnas.0405904101 |
| [17] |
J.D.A. Tyndall, B. Pfeiffer, G. Abbenante, et al., Chem. Rev. 105 (2005) 793-826. DOI:10.1021/cr040689g |
| [18] |
W.L. DeLano, M.H. Ultsch, A.M. de Vos, et al., Science 287 (2000) 1279-1283. DOI:10.1126/science.287.5456.1279 |
| [19] |
Y. Jiang, H. Long, Y. Zhu, Y. Zeng, Chin. Chem. Lett. 29 (2018) 1067-1073. DOI:10.1016/j.cclet.2018.05.028 |
| [20] |
P. Yang, C. Yang, K. Zhang, et al., Chin. Chem. Lett. 29 (2018) 1181-1184. DOI:10.1016/j.cclet.2018.01.045 |
| [21] |
R.T. Shukla, Y.U. Sasidhar, Proteins 83 (2015) 1352-1367. DOI:10.1002/prot.24831 |
| [22] |
I.L. Batalha, I. Lychko, R.J.F. Branco, O. Iranzo, A.C.A. Roque, Org. Biomol. Chem. 17 (2019) 3996-4004. DOI:10.1039/C9OB00564A |
| [23] |
Y.G. Shi, Y.F. Wang, L. Jayaraman, et al., Cell 94 (1998) 585-594. DOI:10.1016/S0092-8674(00)81600-1 |
| [24] |
R.P. Leon, M. Tecklenburg, R.A. Sclafani, Genetics 179 (2008) 1757-1768. DOI:10.1534/genetics.108.088690 |
| [25] |
M.L. Li, D.R. Zhou, H. Zhao, et al., Chin. Chem. Lett. 20 (2009) 1315-1318. DOI:10.1016/j.cclet.2009.06.008 |
| [26] |
J.S. Nowick, D.L. Holmes, G. Mackin, et al., J. Am. Chem. Soc. 118 (1996) 2764-2765. DOI:10.1021/ja953334a |
| [27] |
H.Q. Zeng, X.W. Yang, R.A. Flowers, et al., J. Am. Chem. Soc. 124 (2002) 2903-2910. DOI:10.1021/ja010701b |
| [28] |
L. Sebaoun, V. Maurizot, T. Granier, et al., J. Am. Chem. Soc. 136 (2014) 2168-2174. DOI:10.1021/ja412729s |
| [29] |
J.E. Ross, P.C. Knipe, S. Thompson, et al., Chem. Eur. J. 21 (2015) 13518-13521. DOI:10.1002/chem.201502690 |
| [30] |
B. Gole, B. Kauffmann, V. Maurizot, et al., Angew. Chem. Int. Ed. 58 (2019) 8063-8067. DOI:10.1002/anie.201902378 |
| [31] |
J.D. Pham, N. Chim, C.W. Goulding, et al., J. Am. Chem. Soc. 135 (2013) 12460-12468. DOI:10.1021/ja4068854 |
| [32] |
D. Seebach, S. Abele, K. Gademann, et al., Angew. Chem. Int. Ed. 38 (1999) 1595-1597. DOI:10.1002/(SICI)1521-3773(19990601)38:11<1595::AID-ANIE1595>3.0.CO;2-0 |
| [33] |
X. Daura, K. Gademann, H. Schäfer, et al., J. Am. Chem. Soc. 123 (2001) 2393-2404. DOI:10.1021/ja003689g |
| [34] |
S. Chatterjee, P.G. Vasudev, S. Raghothama, et al., J. Am. Chem. Soc. 131 (2009) 5956-5965. DOI:10.1021/ja900618h |
| [35] |
B. Gong, Acc. Chem. Res. 45 (2012) 2077-2087. DOI:10.1021/ar300007k |
| [36] |
B. Gong, Y. Yan, H. Zeng, et al., J. Am. Chem. Soc. 121 (1999) 5607-5608. DOI:10.1021/ja990904o |
| [37] |
Y.D. Shi, Q. Tang, Y.F. Jiang, et al., Chem. Commun. 54 (2018) 3719-3722. DOI:10.1039/C8CC01564C |
| [38] |
Y.K. Zhang, R.K. Cao, J. Shen, et al., Org. Lett. 20 (2018) 1555-1558. DOI:10.1021/acs.orglett.8b00283 |
| [39] |
Y.K. Zhang, Y.L. Zhong, A.L. Connor, et al., J. Am. Chem. Soc. 141 (2019) 14239-14248. DOI:10.1021/jacs.9b06094 |
| [40] |
Q. Tang, Y.L. Zhong, D.P. Miller, et al., Org. Lett. 22 (2020) 1003-1007. DOI:10.1021/acs.orglett.9b04547 |
| [41] |
Y.L. Zhong, Q. Tang, D.P. Miller, et al., Front. Chem. 8 (2020) 530083. DOI:10.3389/fchem.2020.530083 |
| [42] |
G. te Velde, F.M. Bickelhaupt, E.J. Baerends, et al., J. Comput. Chem. 22 (2001) 931-967. DOI:10.1002/jcc.1056 |
| [43] |
G. Kresse, J. Furthmuller, Phys. Rev. B 54 (1996) 11179-11186. DOI:10.1103/PhysRevB.54.11169 |
| [44] |
S. Grimme, J. Antony, S. Ehrlich, et al., J. Chem. Phys. 132 (2010) 154104. DOI:10.1063/1.3382344 |