 2020, Vol. 31
2020, Vol. 31 


Biomimetic scaffolds were applied in the regeneration of wounded nervous tissue, in order to avoid secondary injury from the autologous tissue grafting and rejection from allotransplantation [1, 2]. Biodegradable polymers, such as poly(lactic-co-glycolic acid), polycaprolactone and poly(L-lactic) acid (PLLA), were used in tissue engineering for the central and peripheral neural regeneration, due to their easy processability and good in vivo compatibility [3-5]. Especially, aligned PLLA nano/micro-fibrous film could be used as the morphology scaffold to guide axons growth and the nerve regeneration [6-9].
Conducting polymers (CPs), such as polypyrrole (PPy), were potential tissue replacements due to their good conductivity, biocompatibility and easy modification, especially PPy was also easily coated PLLA fibers [10-12]. With the modifications of protein and growth factors, PPy could further be used in the fields of biosensors, tissue engineering, and neural probes [11-13]. Furthermore, the electrical stimulation (ES) through CPs could increase the nerve cells activity [14, 15], improving neural axons growth and extension [16, 17].
Biomolecules significantly affected cell adhesion, differentiation and axon elongation. The components of cell-derived extracellular matrix (ECM), such as laminin, fibronectin and collagen, could improve cell adhesion and proliferation [18]. Laminin, as one of ECM components, could promote cell attachment with syndecans and spreading with integrin α2β1 [19, 20]. Fibronectin and collagen were reported to improve cell attachment and topographically to guide axon elongation [21, 22]. Furthermore, biosynthetic conduits with neural stem cell-derived ECM could promote the regeneration of peripheral and central nervous system, because these ECMs from neural stem cells included several trace neural growths [23, 24].
Although ECM from non-neuronal cells did not contain the trace neural growths, its components could improve the attachment of neural cells on the surface of some hydrophobic polymers. Many growth factors have been applied in the nerve differentiation and growth, and nerve growth factor (NGF) is a most important and effective neurotrophic factor to induce and promote nerve growth [25, 26]. Epidermal growth factor (EGF) could also affect Trk A and p75-NTR autophosphorylation sites to stimulate axon growth in the early period [17, 27]. EGF and NGF synergistically acted on nerve tissue engineering responsible for never differentiation and affording nutrients via chimeric EGF-NGF receptors to elongate axon [27-31]. Furthermore, conductive NGF-immobilized fibrous scaffolds could induce PC12 cell (pheochromocytoma clonal cell line) differentiation and axon growth [32], because PC12 is a widely used cell line as the paradigm for neurobiological study [33]. It is reported that neurotrophic factors, including EGF and NGF, could promote the differentiation of mesenchymal stem cells [34]. ES could alter the voltage-gated ion channels of axon to accelerate functional recovery of sciatic nerve gap bridged by aligned microchannels scaffolds [35, 36]. Recently, Yang et al. found that ECM from non-neuronal cells on conducting aligned fiberfilms improved the attachment of PC12 cells, and the amounts of cells adhered on ECM-coated films were significantly higher than that on laminin-coated film or collagen-coated film [37]. However, the synergitic effect of EGF/NGF and ECM from non-neuronal cells on the axon elongation is challenging.
L929 cell, as a murine fibroblast cell line, could be cultured and lysed for the acquirement of ECM from non-neural cells. PC12 cell, as a neural cell model, was also used in neuro-chemical research due to their readily quantifiable, rapid and reversible response to NGF [38, 39]. In this work, the conducting porous PPy-PLLA fiberfilms were first prepared by electrospinning and oxidation polymerization. The combined concentration of EGF and NGF in culture media was analyzed to obtain the optimal condition of axon growth. The synergistic effect of EGF/NGF and ECM-coated PPy-PLLA fiber-films on PC12 differentiation and axon extension under ES was also investigated.
PLLA fiber-filmwas prepared via electrospinning as described in previous reports [40-42]. Subsequently, in situ chemical oxidation polymerization of pyrrole on the surface of aligned PLLA fibers was performed [43, 44]. Different concentrations of EGFand NGF (Pepro Tech, USA) were mixed together to analyze effect of their various concentrations or ratios on differentiation of PC12 cells and axon growth. First, NGFand EGF were separatelyadded in original Ham's F-12 K media, containing 5% (v/v) fetal bovine serum, 10% (v/v) horse serum and 1% (v/v) penicillin-streptomycin. Subsequently, the concentrations of EGF/NGF in the original media were gradually changed as following: 25/0, 0/50, 12.5/50 and 25/50 (ng/mL), according to the previous report [27-31]. Prior to cells seeding into tissue culture plates, laminin (Sigma-Aldrich, USA) solution was used to soak and coat 24 wells plate for 24 h in order to better attach cells on plate. Then the above media with different concentration of EGF and NGF were added into 3 wells with PC12 cells suspensions of 1 ×104 cells/cm2, respectively, and cells were cultured for 5 days, and observed by optical microscopy and imaged at 1, 2, 3 and 5 d, respectively. All media were refreshed every other day. Afterwards, the axons bearing cells and axons length were observed and analyzed with microscope and ImagePro Plus, respectively. Their significant difference was analyzed with a one-way analysis of variance (ANOVA).
PC12 cells on ECM-coated PPy-PLLA fibers with the different media were analyzed the axon growth and orientation along the fibers. The cells on aligned ECM-fiber-films were immune-stained and analyzed effects of NGF (50 ng/mL) and EGF/NGF (12.5/50 ng/mL) on axon growth. The average axons length and neurite bearing cells in different conditions were statistically analyzed by measuring the length from the tip of the axon to the cell body [45]. Their significant difference was analyzed with ANOVA.
A ES device (Fig. S2 in Supporting information) for axons growth was fabricated according to previous report [45]. PC12 cells were seeded on sterilized ECM-PPy-PLLA fiber-films at a density of 1 ×104 cells/cm2 and cultured in the medium with 12.5 ng/mL of EGF and 50 ng/mL of NGF. Voltages of ES is a crucial factor to affect neuron elongation [37, 46, 47]. With a direct voltage resource (Qianfeng Electronic Co., Shanghai, China), the voltage of 100, 200, 400 and 800 mV/cm were exerted between two electrodes, respectively, and the direction of electricity field was parallel to the fibers axis. Before ES, PC 12 cells were cultured on ECM-PPyPLLA for 12 h to permit cells attachment. Cells on ECM-PPy-PLLA fiber-films were continuouslyexposed under the electrical field for 2, 4 and 8 h per day, respectively, totally cultured under ES for 2 d. A cell-sample cultured on ECM-PPy-PLLA fiber-films for 48 h without ES was as control group.
Subsequently, cells were immune-stained to analyze the optimal ES conditions. Inverted fluorescence microscope (IFM, Olympus IX71, Japan) and confocal laser scanning microscope (CLSM, Olympus FV1000 IX81, Japan) were used to observe and analyze the length, alignment and morphology of axons, respectively. The alignment of axons on aligned fibers was quantified via measuring the angle between axon major axis and the fiber axis (or the direction of electric field). The axon of less than 10° angle with fiber axis was considered to align along the fiber axis [48]. In addition, these cell-samples were fixed with 4% glutaraldehyde in PBS (pH ~7.2) for 3 h. Then the fixed cells were dehydrated with a series of ethanol/water solutions (30%, 50%, 70%, 80%, 85%, 90%, 95% and 100%, respectively, each for 15 min and 100% for another 15 min). These cells on PPy-PLLA fiber-films were placed into a pressure vessel of supercritical drying (Quorum K850x, the United Kingdom), and supercritical CO2 was slowly added at 4–6 ℃ for 6 h; subsequently the system temperature was gradually increased till the supercritical state (72.9 ×101, 325 Pa and 31.1 ℃) and this state was kept for 0.5 h; then CO2 gas was quickly released till the normal press, to obtain the dried cell samples. Finally, the samples were sputter-coated with Au nanoparticles (NPs) for SEM observation. All of cellular fluorescent images were obtained under fluorescence microscope after these cells were immunestained. The immunostaining details are shown in Supplementary Information. A one-way analysis of variance (ANOVA) was used to analyze their significant difference.
The morphology and aligned structure of fiber-film were observed with SEM. As shown in Fig. 1a, aligned porous PLLA fibers with 200–600 nm diameter are obtained by electrospinning. The inset image in Fig. 1a shows that there are lots of elliptical pores with 50–300 nm size in PLLA fibers. The image in Fig. 1b shows that PPy-PLLA fibers were rough and porous, and the magnified image shows PPy coating layer with many small irregular NPs with 10–50 nm diameter, and some pores were filled with PPy NPs. The cross-section images of PLLA and PPy-PLLA fibers in Figs. S3a and b (Supporting information) indicatethe inner pores and external openings in the PPy-PLLA fibers. TEM micrographs of PLLA (Figs. S3c and e in Supporting information) and PPyPLLA (Figs. S3d and f in Supporting information) show that PPyPLLA fibers were core-shell structure, with the existence of inner pores. According to the reported preparation condition [46], the longer PPy polymerization time of 12 and 24 h resulted in the more and larger aggregations of PPy NPs absorbed on the fibers surface (Fig. S4 in Supporting information).

|
Download:
|
| Fig. 1. SEM images of (a) aligned porous PLLA fiber-films, and (b) PPy-PLLA fiberfilms. | |
{kind=link}
FTIR spectra of PLLA, PPy NPs and PPy-PLLA in Fig. S5 (Supporting information) reveal that PPy NPs were coated onto PLLA fibers, and PGlu and DBS were co-doped into PPy NPs. XRD patternsof PPy-PLLA fiber-films in Fig. S6 (Supporting Information) show the amorphous broad peaks and corresponding weaker typical crystallization peaks at 16.8°, 19.2° and 22.5° from PLLA, 20.5° and 26.8° from PPy, indicating the deposition of PPy NPs and barely change on crystal form during the fabrication process. However, the interplaner spacing induced peak shifted from 26.8° to 26.5° of PPy, resulting from the weaker polar van der Waals' force between the pyrrole rings and the ester groups of PLLA. 13C NMR spectra of PLLA, PPy and PPy-PLLA shown in Fig. S7 (Supporting Information) revealed that PPy NPs were weakly coated on PLLA fibers, and the slight shifts of C-N of PPy should result from the van der Waals' bond between surplus protons of N atom in pyrrole rings of PPy and O atom of ester groups of PLLA. Contact angles of PPy-PLLA are shown in Fig. S8 (Supporting information), suggesting the DBS-doped PPy coating layers increased the hydrophilicity of fiber-films. The result in Fig. S9 (Supporting information) shows that the conductivity of PPy-PLLA fiber-films are 7.94 and 11.51 mS/m at perpendicular and parallel directions of fibers axis, respectively, indicating a better conductivity in the parallel direction of PPy-PLLA, due to the array of PPy NPs along the fiber axis [49].
As the biomaterial, it is necessary to evaluate the cytocompatibility of PPy-PLLA fiber-films. Fig. S10a (Supporting information) show the results of CCK-8 test of L929 and PC12 cells, respectively, which were cultured on aligned PLLA and PPy-PLLA fibers, demonstrating the good cytocompatibility of PPy-PLLA fiber-films on L929 and PC12. The fluorescent images in Figs. S10b–g (Supporting information) show laminin, collagen type IV and fibronectin from L929 cells cultured on the PPy-PLLA fiber-films, respectively, and L929 cells were adhered on PPy-PLLA and a lot of fluorescence dots were displayed at each cell in the images of Figs. S10b–d. After the cells were lysed, the immunostaining protocol was performed. The fluorescent images in Figs. S10e–g show that, although there was no intact cell, there were a large amount of corresponding fluorescence dots on the fiber-films (including laminin, collagen IV and fibronectin), indicating that cell-derived ECM was obtained via L929 culture and lysing. Fig. S10 h (Supporting information) shows the difference of fluorescence intensities of a series of images of three types of proteins, suggesting the significantly higher fluorescence intensity of laminin on fiber-film and most laminins from the lyed L929 cells conjugated on fibers-film. The results in Figs. S11 and S12 (Supporting information) indicated that the tensile strength and hydrophilicity of the ECM-coate films were better than those of non ECM-coated films. Fig. S13a (Supporting information) show the results of CCK-8 test of PC12 cells cultured on PPy-PLLA and ECM-PPy-PLLA fiber-films, indicating the better cytocompatibility of ECM-PPy-PLLA fiber-films on PC12 cells. Fig. S13b (Supporting information) showed that the hemolysis rate of ECM-coated films was much lower than that of non-ECM films at each concentration, due to the biocompatible ECM from L929.
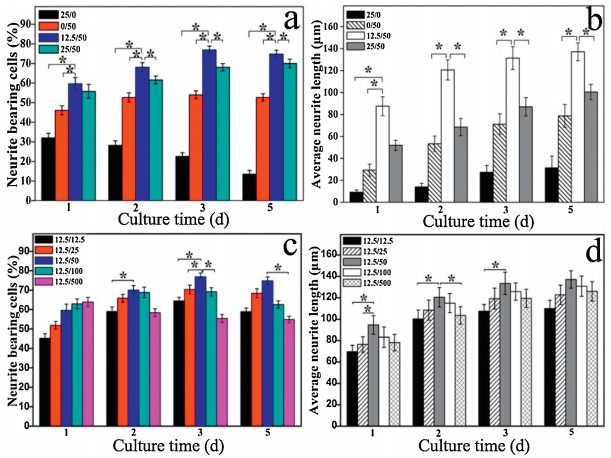
In order to obtain the optimal ratio of two growth factors, PC12 cells were cultured for 1–3 d in the media with different concentrations of EGF and NGF. The images of cells cultured for 1, 2, 3 and 5 days under different concentrations of EGF and NGF (25/0, 0/50, 12.5/50, 25/50 ng/mL) are shown in Fig. S14 (Supporting information), and the percentages of neurite bearing cells cultured in the media with 25/0, 0/50, 12.5/50 and 25/50 ng/ mL of EGF/NGF at 1, 2, 3 and 5 d, respectively, are shows Fig. 2a. The percentages of neurite bearing cells in only EGF-added group reduced from ~32% at 1 d to ~12% at 5 d, indicating the early and lower effect of EGF on differentiation of PC12 cells. The percentages of neurite bearing cells treated by EGF/NGF groups show firstly increased and then decreased with time, reaching to the peak values at the 3rd day. The percentages of neurite bearing cells in EGF and NGF co-added groups were higher than that of only NGF group at the corresponding days, indicating that co-addition of EGF and NGF (optimal ratio is 12.5/50) could significantly promote PC12 differentiation to protrude axons. The results of average neurite length in Fig. 2b show that the axons length of only EGF group (25/0) at 5 d culture only reached to ~32 μm, significantly less than other groups with NGF, indicating the primary role and obvious promotion of NGF on PC12 differentiation and axons extension, because NGF could be incorporated by specific receptors (TrkA and p75) to terminate cell proliferation subsequently, activate the cell differentiation to protrude the axon. EGF could only stimulate Ras/extracellular signal-regulated kinases (RAS/ ERK) in a transient way, leading to cell differentiation at a booming speed in the early stage and then quickly terminate [17, 27, 31]. The average neurite length of 12.5/50 group was gradually increased with the time of culture, namely, ~87, ~125, ~131, ~136 μm at 1, 2, 3 and 5 d, respectively, significantly larger than the corresponding values of other groups; however, the axons could not significantly extend after 3 d culture. When NGF concentration was 50 ng/mL, the axons length only in 12.5 ng/mL of EGF reached to the largest values (~131 μm at 3 d and ~136 μm at 5 d), because with the increase of EGF, a large portion of fiber-film was occupied by EGF specific receptors, leading to prefer differentiation to proliferation of PC12 cells [17, 27, 50].

|
Download:
|
| Fig. 2. (a) Percentages of neurites bearing cells and (b) neurite lengths of cells cultured with the concentrations of EGF/NGF of 25/0, 0/50, 12.5/50, 25/50 (ng/mL) for 1, 2, 3 and 5 d. (c) Percentages of neurites bearing cells and (d) neurite lengths treated with the concentrations of EGF/NGF of 12.5/12.5, 12.5/25, 12.5/50, 12.5/100, 12.5/500 (ng/mL) for 1, 2, 3 and 5 d. * shows the significant statistical differences between two corresponding groups, P < 0.05. | |
{kind=link}
PC12 differentiation in the media with same EGF and different NGF with the concentrations of 12.5/12.5, 12.5/25, 12.5/50, 12.5/ 100 and 12.5/500 ng/mL, respectively (Fig. S15 in Supporting information). The percentages of neurite bearing cells of different EGF/NGF groups in Fig. 2c show that the percentage of 12.5/500 ng/mL group reached to ~64% at 1 d (higher than others) and then slowed down in the next few days, indicating that higher concentration NGF could promote cell differentiation in the early stage, and too much NGF would inhibit the differentiation of PC12 cells in the later stage. The percentage of neurite bearing cells on 12.5/50 group reached to the highest values for 3 and 5 d, further indicating the long term synergy of 12.5 ng/mL of EGF and 50 ng/mL of NGF. Axons length of 12.5/50 groups are ~91, ~125, ~132 and ~137 μm at 1, 2, 3 and 5 d, respectively, and there was the obvious length increase after cells are cultured for 2 d (Fig. 2d), indicating positive effect of EGF and NGF combination on early period. Axons lengths of 12.5/50 groups were larger than the corresponding values of other groups at 1, 2, 3 and 5 d, further indicating that 12.5 ng/mL of EGF and 50 ng/mL of NGF had the good synergy to promote axon elongation.
It can be visually observed that many PC12 cells were attached onto L-PPy-PLLA and ECM-PPy-PLLA fiber-films, suggesting that PC12 cells were easily adhered to L-PPy-PLLA and ECM-PPy-PLLA, because biomolecule–modified substrates could induce more cells to attach (Fig. S16a in Supporting information). The results in Fig. S16b (Supporting information) confirm that the amounts of cells attached on PPy-PLLA were ~2305 and 3909 cells/cm2 at 8 and 12 h, respectively, significantly higher than those of control groups (~13 and 17 cells/cm2), demonstrating that PPy-PLLA provided more sites for cells adhesion. The adhered cells amounts on L-PPy-PLLA and ECM-PPy-PLLA were ~5860 and ~9250 cells/cm2 at 12 h, respectively, indicating that laminin induced more cells to adhere onto PPy-PLLA fibers. The amounts of cells attached on ECM-PPy-PLLAwere significantly larger than the other groups at each interval, indicating that ECM-PPy-PLLA most supported the attachment of PC12 cells, due to several kinds of biomolecules in ECM. FAK played important effect in the organization of adhesion complexes [51, 52]. The FAK results in Fig. S17 (Supporting information) obviously indicatethat, ~430 pg/mL of FAK concentration was obtained in ECM-coated PPy-PLLA groups, obviously larger than those of PPy-PLLA (~421 pg/mL) and ECM-PPy-PLLA with ES groups (~425 pg/mL), due to the direct ligation of integrin extracellular induced by ECM proteins. Although ES could improve cells activity, more dynamic were used to assemble and transport microfilament and microtubule, then pulling the neurite forward, therefore, active cells could not easily be attached on substrates under ES.
Furthermore, the optimal groups were seed on naked and ECMcoated fiber-film to explore effect of ECM on PC12 attachment and axon protrusion. Figs. S18a and b (Supporting information) show the images of PC12 cells on no ECM and ECM-coated fiber-films cultured with 50 ng/mL of NGF for 2 d, respectively, and more cells were better attached on the ECM-coated fiber-film, due to the incorporation of ECM. The results of Fig. S18c (Supporting information) show that the percentage of neurite bearing cells on ECM-coated fibers was ~33.6%, significantly higher than ~24.5% of naked fiber-films. The results of Fig. S18d (Supporting information) clearly show that the axons length on ECM-coated fibers (~67 μm) was significantly larger than that on naked fiberfilms (~42 μm), further indicating that the axons were rapidly adhered and grown on the surface of fibers with ECM conjugation.
The images in Figs. S19a and b (Supporting information) show axons growth and elongation on the aligned ECM-coated fibers added 50 ng/mL of NGFand EGF/NGF (12.5/50 ng/mL), respectively. The percentages of neurite bearing cells in EGF/NGF groups were ~75% at 1 d, ~86% at 2 d and ~83% at 3 d, significantly higher than the corresponding values of NGF groups (~55%, ~76% and ~69%, respectively, P < 0.05, Fig. S19c in Supporting information). The axon lengths in EGF/NGF group reached to ~134, ~149 and ~152 μm at 1, 2 and 3 d, respectively, significantly larger than those in NGF group of ~91, ~95 and 95 μm at 1, 2 and 3 d (P < 0.05, Fig. S19d in Supporting information). These results further suggest that enough biological cues released from EGF/NGF (12.5/50 ng/mL) could better support PC12 differentiation and axons elongation on aligned ECM-fiber-films.
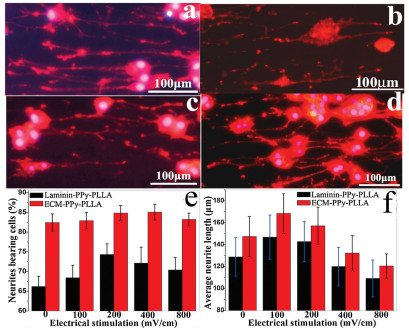
In order to analyze effect of ES on the neurites, the voltage of 100, 200, 400 and 800 mV/cm was exerted onto cells for 2 h/day in 2 day through aligned ECM-fiber-films. Figs. 3a–d show the fluorescent images of PC12 cells in the media with EGF/NGF (12.5/ 50 ng/mL) cultured on ECM- and L-PPy-PLLA, respectively. The results in Fig. 3e show that, the percentages of neurites bearing cells on ECM-PPy-PLLA, larger than those on laminin-PPy-PLLA, were slightly increased from ~82% (100 mV/cm) to ~85% (400 mV/cm) and then slightly decreased to 83% at 800 mV/cm, indicating the higher voltage could not obviously elicit to protrude more axons, and there was no significant difference with that of none ES group (~86%). The results in Fig. 3f show that, axons on ECM-PPy-PLLA were longer than those on L -PPy-PLLA, and axon length under 100 mV/cm on ECM-PPy-PLLA reached to ~174 μm, larger than those in 0, 200, 400 and 800 mV/cm groups (~149, ~154, ~129 and ~118 μm, respectively). These results suggest that the lower ES voltage promoted axon extension, and synergistic effect of ECM and ES could better increase axon extension. The images of cells cultured in the media without NGF (Fig. S20 in Supporting information) suggest that only ES could not well activate differentiation signal of PC12 cells due to addition of no NGF, and ES only improve neurite growth after the cell differentiation.

|
Download:
|
| Fig. 3. Fluorescence images of PC12 cells cultured on aligned ECM-PPy-PLLA fibers in the media with EGF/NGF (12.5/50 ng/mL)under ES for 2 h/d with (a) 100, (b) 200, (c) 400 and (d) 800 mV/cm, respectively. The statistical results of (e) percentage of neurite bearing cells and (f) average neurite length of PC12 cultured on ECM- and laminin-PPy-PLLA under the different ES conditions. * represents significant difference, P < 0.05. | |
{kind=link}
Figs. 4a–d show the axons images on aligned ECM-fiber-films under ES of 100 mV/cm for the different time (0, 2, 4 and 8 h/day in 2 day), suggesting the obvious axons elongation in different ES time. Fig. 4e shows that the percentages of aligned axons along the electro-field direction (less than 10° angle between axons and the direction of fiber axis [45]) are ~52% (0 h/day), ~53% (2 h/day), ~57% (4 h/day) and ~64% (8 h/day), indicating that the axon along major direction of aligned fibers were increased with ES time, and the alignment of axons could further be enhanced under ES of 100 mV/cm for 8 h/day, which is in accordance with the previous results, including dorsal root ganglia explants and neural stemcells [53, 54]. Without ES, the average lengths of axons on ECM-fiberfilms are ~149 μm. Neurite length results in Fig. 4f reveal that, the longest axons length (~251 μm) was obtained in 4 h/day ES group, significantly larger than those of ES of 2 and 8 h/day (~183 μm, ~159 μm, respectively), indicating that 100 mV/cm ES for 4 h/day in 2 day through ECM-fiber-film could obviously promote axon elongation. When ES time reached to 8 h/day, the axons became shorter than that of 2 h/day group, suggesting inhibition effect of ES with long time. The results of alkaline phosphatase (ALP) and reactive oxygen species (ROS) analysis in Fig. S21 (Supporting information) indicated the synergistic effect of ES with EGF/NGF, and ES could enhance the differentiation ability of PC12 cells [55], and improve axons growth due to the generation of lower ROS level. When both of ALP and ROS reached best balance, the axons bearing cells and axons length arrived at the optimal level.

|
Download:
|
| Fig. 4. Immunostaining images of PC12 cells cultured on aligned PPy-PLLA fibers with the treatment of EGF/NGF (12.5/50 ng/mL in the medium and electrical stimulation with 100 mV/cm for (a) 0 h, (b) 2 h, (c) 4 h and (d) 8 h, respectively. (e) Neurite alignment and (f) average neurite length of PC12 cells cultured under the above conditions. Arrows indicate direction of electrical field and aligned fibers axis. * shows the significant difference between two corresponding groups, P < 0.05. Western Blotting results of MAPK, Trk A, EGFR and NF-L form PC12 cells cultured on PPy-PLLA with EGF + NGF with/without ES: (g) PAGE images, (h) semi-quantitative results. | |
{kind=link}
SEM images of PC12 cells on the treatment of NGF (50 ng/mL) and EGF/NGF (12.5/50 ng/mL) in Fig. S22 (Supporting information) suggest that this concentration combination of EGF and NGF could inspire more protrusion sites of axons and promote axon elongation. CLSM images of an axon on the aligned PPy-PPLA fibers under the treatment of EGF/NGF in Figs. S23a and b (Supporting information) show that many fine and slender filopodia grew out from the leading edge of growth cones under the treatment of EGF/NGF, facilitating axons the elongation. In the amplified image of growth cone of the axon tip (Fig. S23b), the inflated outline of leading edge and the swelling central domain show the engorgement of organelles, which were the energy resource of axon extension [56, 57], indicating that EGF/NGF could synergistically induce axons to grow longer. In Fig. S23c (Supporting information), the longerand thinner axon is obtained under ES, and many finer and slender filopodia appear (shown by green arrows in Fig. S23d in Supporting information), indicating that ES accompanied with EGF/NGF could further promote axons growth and filopodia extension. The central domain of the growth cone (Fig. S23d) was shifted to the frontier, indicating that a lot of actins were accumulated in the tip of the filopodia for the promotion of axon elongation.
In order to further research the neurobiologyof PC12 under NGF and synergistic effect of ES and two growth factors, four cellular special proteins were analyzed with western blotting. The images of PAGE (polyacrylamide gel electrophoresis) in Fig. 4g show that, MAPK (microtubule-activated protein kinase), Trk A (NGF receptor), EGFR (EGF receptor) and NF-L (Neurofilament-L) play the important role in nerve cell differentiation and axon growth, and ES leads to the produce of more cellular proteins. The results in Fig. 4h show that the amounts of MAPK, Trk A and NF-L in EGF + NGF groups are higher than the corresponding value of NGF group, especially the amounts of Trk A, EGFR and NF-L in ES group are obviously higher than those without ES groups, indicating that ES can significantly promote the production and expression of related neural proteins.
According to these results, a schematic was illustrated in Fig. S24 (Supporting information), to explain effect of these factors on the growth and elongation of axons. ECM proteins (e.g., laminin, collagen) can be bound with clusters of adhesion receptors [58], and laminins can also enhance the growth of axons [59]. Fig. S24c shows the growth cone easily adhered onto the rough surface. The highly dynamic filopodia at the front end of growth cone could continuously extend and retract, which would help them to response to extracellular cues [60, 61]. Axon on ECM-coated PPyPLLA fibers were elongated through actin and microtubule rearrangement in the growth cone activated by NGF [62, 63]. In Fig. S24d, only some NGF were conjugated on the filopodia surface, because the high-affinity Trk A could steadily bind with NGF, while low-affinity p75-NTR randomly and unsteadily linked with the factors [17, 27]. Tiny amount of EGF could strongly activate cell growth, thus enable high dynamic of filopodia. Moreover, when EGF was combined with corresponding receptor of filopodia, the receptors of NGF (Trk A and p75-NTR) were more inclined to form heteromeric receptors, which could enhance the affinity of NGF with its receptors, leading to the assembly and transportation of more microtubules for the extension of axon (Fig. S24e) [31, 50]. Conducting PPy surface could change the cell membrane potential under ES (Fig. S24f) to improve the dynamic of filopodia and axon, also appeal axon attachment [64]. Subsequently, ES could further enhance actin assembly in filopodia of growth cone [65]. Thus, ES cooperated with EGF/NGF through ECM-coatedfiber-films improved axons attachment, growth and elongation along the fibers axis.
In summary, the porous aligned PPy-PLLA fiber-films with good conductivity and cytocompatibility was fabricated via chemical oxidation polymerization of pyrrole on aligned PLLA fibers, and PPy NPs were coated and combined on PLLA fibers via polar Wan der Waal's force, leading to their surface conductivity of 11.5 mS/m. Non neural-cells-derived ECM was coated on PPy-PLLA fiber-films via L929 culture and lysing, and not only improved attachment of PC12 cells on PPy-PLLA fibers, but also induced the axon extension along the fiber orientation. Compared with the group of only NGF (50 ng/mL), EGF/NGF (12.5/50 ng/mL) in media can modulate cellular interactions, promote PC12 differentiation and improve the percentage of axon-bearing cells. ES on PC12 cells through the ECM-coated fiber-films further improved axons elongation and alignment compared to unstimulated cells. The potential of 100 mV/cm exerted for 4 h/day in 2 day encouraged more cells to protrude their axons for the further elongation, and aligning to the fibers orientation. Axons growth and elongation on aligned conductive PPy-PLLA fiber-films stimulated with the double factors and ES would be a promising process, which provides novel cues for neural tissue regeneration.
AcknowledgmentsThis work has been supported by the National Natural Science Foundation of China (No. 51273122) and Sichuan Science and Technology Project (No. 2018JY0535).
Appendix A. Supplementary dataSupplementary material related to this article can be found, in the online version, at doi:https://doi.org/10.1016/j.cclet.2019.07.002.
| [1] |
L. Tian, M.P. Prabhakaran, S. Regenerative, Biomaterials 2 (2015) 31-45. |
| [2] |
H. Shin, S. Jo, A.G. Mikos, Biomaterials 24 (2003) 4353-4364. DOI:10.1016/S0142-9612(03)00339-9 |
| [3] |
X. Wen, P.A. Tresco, Biomaterials 27 (2006) 3800-3809. DOI:10.1016/j.biomaterials.2006.02.036 |
| [4] |
M.B. Runge, M. Dadsetan, J. Baltrusaitis, et al., Biomaterials 31 (2010) 5916-5926. DOI:10.1016/j.biomaterials.2010.04.012 |
| [5] |
F. Yang, R. Murugan, S. Wang, S. Ramakrishna, Biomaterials 26 (2005) 2603-2610. DOI:10.1016/j.biomaterials.2004.06.051 |
| [6] |
N. Bhardwaj, S.C. Kundu, Biotech. Adv. 28 (2010) 325-347. DOI:10.1016/j.biotechadv.2010.01.004 |
| [7] |
S.H. Lim, H.Q. Mao, Adv. Drug Deliv. Rev. 61 (2009) 1084-1096. DOI:10.1016/j.addr.2009.07.011 |
| [8] |
Y. Yu, X. Lü, F. Ding, J. Biomed. Nanotech. 11 (2015) 816-827. DOI:10.1166/jbn.2015.1973 |
| [9] |
J. Xie, W. Liu, M.R. MacEwan, P.C. Bridgman, Y. Xia, ACS Nano 8 (2014) 1878-1885. DOI:10.1021/nn406363j |
| [10] |
N.K. Guimard, N. Gomez, C.E. Schmidt, Prog. Polym. Sci. 32 (2007) 876-921. DOI:10.1016/j.progpolymsci.2007.05.012 |
| [11] |
P.M. George, A.W. Lyckman, et al., Biomaterials 26 (2005) 3511-3519. DOI:10.1016/j.biomaterials.2004.09.037 |
| [12] |
C.E. Schmidt, V.R. Shastri, J.P. Vacanti, R. Langer, Proc. Natl. Acad. Sci. U. S. A. 94 (1997) 8948-8953. DOI:10.1073/pnas.94.17.8948 |
| [13] |
L. Jin, Z.Q. Feng, M.L. Zhu, et al., J. Biomed. Nanotech. 8 (2012) 779-785. DOI:10.1166/jbn.2012.1443 |
| [14] |
J. Zhang, K. Qiu, B. Sun, et al., J. Mater. Chem. B:Mater. Biol. Med. 2 (2014) 7945-7954. DOI:10.1039/C4TB01185F |
| [15] |
Y. Lu, T. Li, X. Zhao, et al., Biomaterials 31 (2010) 5169-5181. DOI:10.1016/j.biomaterials.2010.03.022 |
| [16] |
D. Blumberg, M. Radeke, S. Feinstein, J. Neurosci. Res. 41 (1995) 628-639. DOI:10.1002/jnr.490410509 |
| [17] |
D.R. Tyson, S. Larkin, Y. Hamai, R.A. Bradshaw, Int. J. Dev. Neurosci. 21 (2003) 63-74. DOI:10.1016/S0736-5748(02)00139-9 |
| [18] |
H. Lu, T. Hoshiba, N. Kawazoe, et al., Biomaterials 32 (2011) 9658-9666. DOI:10.1016/j.biomaterials.2011.08.091 |
| [19] |
K. Hozumi, N. Suzuki, P.K. Nielsen, M. Nomizu, Y. Yamada, J. Biol. Chem. 281 (2006) 32929-32940. DOI:10.1074/jbc.M605708200 |
| [20] |
R. Timpl, H. Rohde, P.G. Robey, et al., J. Biol. Chem. 254 (1979) 9933-9937. |
| [21] |
M.D. Pierschbacher, E. Ruoslahti, Nature 309 (1983) 30-33. |
| [22] |
V.J. Mukhatyar, M. Salmeron-Sanchez, S. Rudra, et al., Biomaterials 32 (2011) 3958-3968. DOI:10.1016/j.biomaterials.2011.02.015 |
| [23] |
A. Alovskaya, T. Alekseeva, J. Phillips, V. King, R. Brown, Top Tissue Eng. 3 (2007) 1-26. |
| [24] |
G. Fabregat, B. Teixeira-Dias, L.J. del Valle, et al., ACS Appl. Mater. Interf. 6 (2014) 11940-11954. DOI:10.1021/am503904h |
| [25] |
R. Levi-Montalcini, Prog. Brain Res. 45 (2011) 235-256. |
| [26] |
T. Dvir, B.P. Timko, D.S. Kohane, R. Langer, Nat. Nanotechn. 6 (2011) 13-22. DOI:10.1038/nnano.2010.246 |
| [27] |
Y.Y. Wu, R.A. Bradshaw, J. Biol. Chem. 271 (1996) 13033-13039. DOI:10.1074/jbc.271.22.13033 |
| [28] |
R.S. Morrison, H.I. Kornblum, F.M. Leslie, R.A. Bradshaw, Science 238 (1987) 72-75. DOI:10.1126/science.3498986 |
| [29] |
N.P. Tsai, Y.C. Tsui, J.E. Pintar, H.H. Loh, L.N. Wei, P. Natl. Acad. Sci. U. S. A. 107 (2010) 3216-3221. DOI:10.1073/pnas.0912367107 |
| [30] |
G.J.R. Delcroix, K.M. Curtis, P.C. Schiller, C.N. Montero-Menei, Differentiation 80 (2010) 213-227. DOI:10.1016/j.diff.2010.07.001 |
| [31] |
H. Yan, J. Schlessinger, M.V. Chao, Science 252 (1991) 561-563. DOI:10.1126/science.1850551 |
| [32] |
J.Y. Lee, C. Bashur, C. Milroy, et al., IEEE T. Nano Biosci. 11 (2012) 15-21. DOI:10.1109/TNB.2011.2159621 |
| [33] |
K. Alheim, C. Andersson, S. Tingsborg, et al., Proc. Natl. Acad. Sci. U. S. A. 88 (1991) 9302-9306. DOI:10.1073/pnas.88.20.9302 |
| [34] |
M.P. Prabhakaran, J.R. Venugopal, S. Ramakrishna, Biomaterials 30 (2009) 4996-5003. DOI:10.1016/j.biomaterials.2009.05.057 |
| [35] |
J. Huang, L. Lu, X. Hu, et al., Neurorehabil. Neural Repair 24 (2010) 736-745. DOI:10.1177/1545968310368686 |
| [36] |
J. Zhang, M. Li, E.T. Kang, K.G. Neoh, Acta Biomater. 32 (2016) 46-56. DOI:10.1016/j.actbio.2015.12.024 |
| [37] |
X. Zhou, A. Yang, Z. Huang, et al., Colloids Surf. B:Biointerfaces 149 (2017) 217-225. DOI:10.1016/j.colsurfb.2016.10.014 |
| [38] |
J.D. Foley, E.W. Grunwald, P.F. Nealey, C.J. Murphy, Biomaterials 26 (2005) 3639-3644. DOI:10.1016/j.biomaterials.2004.09.048 |
| [39] |
J. Zeng, Z. Huang, G. Yin, et al., Colloids Surf. B:Biointerfaces 110 (2013) 450-457. DOI:10.1016/j.colsurfb.2013.05.012 |
| [40] |
J. Ding, J. Zhang, J. Li, et al., Prog. Polym. Sci. 90 (2019) 1-34. DOI:10.1016/j.progpolymsci.2019.01.002 |
| [41] |
J. Li, W. Xu, D. Li, et al., ACS Nano 12 (2018) 6685-6699. DOI:10.1021/acsnano.8b01729 |
| [42] |
J. Li, W. Xu, J. Chen, et al., ACS Biomater. Sci. Eng. 4 (2018) 2026-2036. DOI:10.1021/acsbiomaterials.7b00605 |
| [43] |
J. Li, X. Feng, J. Shi, et al., ACS Omega 3 (2018) 2715-2723. DOI:10.1021/acsomega.8b00090 |
| [44] |
S. Li, S. Dong, W. Xu, et al., Adv. Sci. 5 (2018) 1700527. DOI:10.1002/advs.201700527 |
| [45] |
A. Yang, Z. Huang, G. Yin, X. Pu, Colloids Surf. B:Biointerfaces 134 (2015) 469-474. DOI:10.1016/j.colsurfb.2015.07.028 |
| [46] |
J.Y. Lee, C.A. Bashur, A.S. Goldstein, C.E. Schmidt, Biomaterials 30 (2009) 4325-4335. DOI:10.1016/j.biomaterials.2009.04.042 |
| [47] |
J. Xie, M.R. Macewan, S.M. Willerth, et al., Adv. Funct. Mater. 19 (2009) 2312-2318. DOI:10.1002/adfm.200801904 |
| [48] |
H. Aubin, J.W. Nichol, C.B. Hutson, et al., Biomaterials 31 (2010) 6941-6951. DOI:10.1016/j.biomaterials.2010.05.056 |
| [49] |
U. Malhotra, S. Maity, A. Chatterjee, J. Appl. Polym. Sci. 132 (2015) 41336. |
| [50] |
X. Feng, J. Li, X. Zhang, et al., J. Control. Release 302 (2019) 19-41. DOI:10.1016/j.jconrel.2019.03.020 |
| [51] |
X. Chen, X. Zhuang, C. Fu, et al., Curr. Pharm. Design 21 (2015) 1960-1966. DOI:10.2174/1381612821666150302152420 |
| [52] |
Z. Jin, T. Zheng, E. Alarçin, et al., Small 13 (2017) 1701949. DOI:10.1002/smll.201701949 |
| [53] |
F. Yang, R. Murugan, S. Wang, S. Ramakrishna, Biomaterials 26 (2005) 2603-2610. DOI:10.1016/j.biomaterials.2004.06.051 |
| [54] |
J.M. Corey, D.Y. Lin, K.B. Mycek, et al., J. Biomed. Mater. Res. A 83 (2007) 636-645. |
| [55] |
G. Bartosz, Biochem. Pharmacol. 77 (2009) 1303-1315. DOI:10.1016/j.bcp.2008.11.009 |
| [56] |
S.R. Heidemann, Int. Rev. Cytol. 165 (1996) 235-296. DOI:10.1016/S0074-7696(08)62224-X |
| [57] |
N. Patel, M.M. Poo, J. Neurosci. 2 (1982) 483-496. DOI:10.1523/JNEUROSCI.02-04-00483.1982 |
| [58] |
Y. Sun, H. Shan, J. Wang, et al., J. Biomed. Nanotech. 15 (2019) 939-950. DOI:10.1166/jbn.2019.2745 |
| [59] |
S. Wang, Z. Zhang, D. Jiang, et al., Polymers 8 (2016) 200. DOI:10.3390/polym8050200 |
| [60] |
Z. Jin, T. Zheng, E. Alarçin, et al., Small 13 (2017) 1701949. DOI:10.1002/smll.201701949 |
| [61] |
J. Zhang, H. Liu, J.X. Ding, et al., RSC Adv. 5 (2015) 32604-32608. DOI:10.1039/C5RA05530J |
| [62] |
D.J. Goldberg, D.W. Burmeister, J. Cell Biol. 103 (1986) 1921-1931. DOI:10.1083/jcb.103.5.1921 |
| [63] |
D. Johnson, A. Lanahan, C.R. Buck, et al., Cell 47 (1986) 545-554. DOI:10.1016/0092-8674(86)90619-7 |
| [64] |
P. Lamoureux, S.R. Heidemann, N.R. Martzke, K.E. Miller, Dev. Neurobiol. 70 (2010) 135-149. DOI:10.1002/dneu.20764 |
| [65] |
P.R. Chen, M.H. Chen, F.H. Lin, W.Y. Su, Biomaterials 26 (2005) 6579-6587. DOI:10.1016/j.biomaterials.2005.03.037 |