 2019, Vol. 30
2019, Vol. 30 


b Department of Chemistry and Chemical Engineering, Beijing University of Technology, Beijing 100124, China
Organic phosphates are a large class of phosphates in organisms, including various nucleotides, nicotinamide adenine dinucleotide (NADH), flavin adenine dinucleotide (FAD), phospholipids [1]. These organic phosphate biomolecules (OPBs) have multiple roles and physiological functions in vivo, such as energy storage [2], signal transduction [3], DNA and RNA synthesis [4], as important coenzyme factors [5], regulating redox balance [6], maintaining cell growth and membrane size [7]. It is well known that monitoring of organic phosphate biomolecules has become increasingly popular in recent years due to their important roles [8, 9]. However, the wide variety of compositions and small differences in structure pose significant challenges for identifying and monitoring them, such as nucleotides and glycerophospholipids.
With the joint efforts of biologists and chemists, many advanced analytical techniques have been used to monitor these important OPBs to obtain valuable information, such as nuclear magnetic resonance spectroscopy (1H and 31P NMR), mass spectrometry, high-performance liquid chromatography, and electrochemical methods [8]. However, due to the complexity of operation and poor sensitivity, these methods are difficult to apply to in situ monitoring in cells or organisms. Fluorescence imaging technology has become a powerful analytical tool widely used in bioassays and clinical diagnostics due to its high resolution, realtime monitoring, good selectivity and sensitivity [10, 11]. More importantly, small molecule fluorescent probes have excellent advantages, such as tailored properties, biodegradability, and are widely used for reactive oxygen species, nitrogen, sulfur species (ROS, RNS, RNS), metal ions and enzymes in vitro or in vivo [12-17]. For organic phosphates that play an important physiological role in complex living systems, the use of fluorescent probes for in vitro or in vivo monitoring and imaging is a good choice. To date, a large number of fluorescent probes have been developed and reported for sensing and monitoring OPBs [8, 18].
Fluorescent probes for a certain type of nucleotide such as adenosine triphosphate (ATP) have been reviewed in the past decade [19]. It is necessary to systematically study the fluorescent probes of the entire OPBs, but as far as we know, there is no review report for the entire OPBs. In order to fully and systematically understand current progress, this review aims to provide a summary of the various OPBs small-molecule fluorescent probes reported from 2010 to 2019, and is divided into three categories based on the structures of OPBs: nucleotides (such as RTP, RDP, RMP and R = adenosine (A), guanosine (G), thymidine (T), cytidine (C), uridine (U)), nicotinamide adenine dinucleotide (NAD(P)H) and flavins nucleotide (FAD/FMN), as well as phospholipids (such as phosphatidylserine (PS)), and a brief introduction to the physiological functions of the respective OPBs. In this review, we pay close attention to the strategy of fluorescent probe design, the mechanism of recognition, and its biological applications, which will provide a systematic overview of fluorescent probes for OPBs and gain a deeper insight into biology from the current advances. Although other fluorescent probe techniques such as genetically encoded probes and nanoprobes have made good progress in the detection and imaging of OPBs [20-22], they are beyond the scope of this review.
2. General strategies for designing OPBs fluorescent probesIn view of the negative charge of OPBs, most probe designs are based on electrostatic interactions that are derived from the development of protein-binding receptors [23, 24]. And there are other probe design strategies adapted from pharmaceutical research and supramolecular chemistry, such as drug screening and host-guest construction [25, 26]. Therefore, we summarized the general strategies for constructing and designing fluorescent probes for OPBs: (1) The structural commonality of OPBs is negative phosphate, which mainly includes nucleotides and phospholipids, therefore, positively charged probes (such as metal complexes, positively charged receptors) are the best choice for combination; (2) Hydrogen bonding is another way of combination between phosphates and probes; (3) π-π Stacking between probe and nucleotide bases is the excellent idea to increase the selectivity of probes; (4) A new strategy of covalent connection based on special chemical reactions has emerged; (5) Distinct functions of NADH/NAD(P)H and FAD/FMN determine that their recognition can be achieved by redox reaction; (6) Multiple strategic synergistic interactions are more effective in selective differentiation and detection.
3. Fluorescent probes for nucleotides (RTP, RDP, RMP, R=A, G, T, C, U)Nucleotides are indispensable biomolecules in almost all biological processes and participate in multiple functions of organisms, ranging from structure, energy supply, metabolism, catalysis and signal regulation to genetics [27, 28]. Nucleotides are composed of heterocyclic nitrogenous bases (mainly pyrimidines and purines), sugars and phosphates, which further constitute the fundamental molecules of nucleic acids and monomers of RNA and DNA. The six-membered ring of pyrimidine mainly includes thymine (T), cytosine (C), and uracil (U), while purine includes adenine (A) and guanine (G). Among all nucleosides, ATP is the most widely studied by the fact that it is a universal energy source and is widely distributed in the body. Importantly, ATP also acts as an ion channel, a neurotransmitter and signaling molecule that regulates cellular movement throughout the organism. Guanine triphosphate (GTP) is another nucleotide with three phosphates groups. It is involved in the synthesis of RNA, participates in the citric acid cycle, and also provides energy for several metabolic reactions [29]. Similarly, uridine-based UTP, thymidine-based TTP and cytidine-based CTP are also important members of nucleotides. UTP is a substrate for RNA synthesis, and TTP and CTP act as substrates for DNA synthesis and are involved in DNA replication and cell division [30]. Adenosine diphosphate (ADP) is a product of ATP hydrolysis, and other diphosphate nucleotides including cytidine diphosphate (CDP), thymidine diphosphate (TDP), uridine diphosphate (UDP) and guanine diphosphate (GDP) are the substrates for the synthesis of corresponding triphosphate nucleotides. Monophosphate nucleotides including adenosine monophosphate (AMP), guanosine monophosphate (GMP), thymidine monophosphate (TMP), cytidine monophosphate (CMP), and uridine monophosphate (UMP) act as intermediates in the synthesis of nucleic acids. In addition to these nucleotides, other cyclic nucleotides include cyclic adenosine monophosphate (cAMP) and guanosine monophosphate (cGMP), which serve as second messengers of hormonal regulation as well as other physiological events [31]. Each specific nucleotide corresponds to a unique function and role, and participates in different signal regulation and related balances. Therefore, selective detection of various nucleotides and real-time monitoring of their levels is necessary to comprehensively understand the relationship between concentration changes and physiological effects. As mentioned above, the selective recognition of specific nucleotides from so many nucleotides is a huge challenge due to differences in nucleobase type and phosphate number, which has promoted a large number of researchers to focus on designing fluorescent probe for monitoring nucleotides.
3.1. Fluorescent probes for ATPDue to the importance of ATP in various biological processes, many fluorescent probes for recognition of ATP have been developed and reported based on the specific structure of ATP in vitro or in vivo, including electrostatic interaction between positively charged probes and negatively charged phosphates, formation of hydrogen bonds between probes with the oxygen atom of the phosphate, π-π stacking interaction between aromatic structures of probes and adenine, a covalent bonding between boronic acid and ribose and synergistic interaction by multiple ways between probes and ATP.
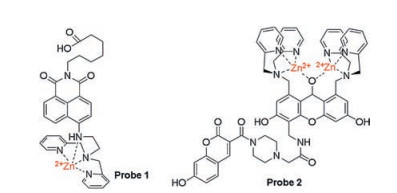
3.1.1. Recognition of ATP based on electrostatic interaction between positive and negative chargesIn the early days of ATP probe design, positively charged metal complexes were used as ATP recognition sites because they provided a stronger affinity for the three negatively charged phosphates of ATP. Fluorescent probes based on Zn(Ⅱ) complexes are more promising candidates than other metal-complexed probes. For example, fluorescent probes possessing dipicolylamine (DPA) unit for the detection of phosphate species, especially for ATP, are well known and have been reported by numerous researchers [32]. In 2010, Moro et al. developed probe 1 (Fig. 1) based on naphthalimide chemosensor with a Zn(Ⅱ)-DPA complex unit for ATP detection [33]. In order to immobilize the probe on a surface or biomolecule by a succinimidyl-ester reaction, a carboxylic group is introduced into the probe molecule. To the best of our knowledge, the introduction of DPA quenches the fluorescence of aminonaphthalimide due to deprotonation of the aromatic amine under alkaline media. After DPA coordinated with Zn2+, the fluorescence of aminonaphthalimide shows a slight increase for two reasons: a decrease in photoinduced electron transfer (PET) effect from DPA and a speed-up of the deprotonation of aromatic amines under neutral pH conditions via the electrostatic interaction of Zn2+-NH. After the gradual introduction of ATP, the interaction of Zn2+-NH through the binding of ATP to Zn(Ⅱ)-DPA is reduced, the fluorescence at 535 nm is increased by 2.3-fold even when the concentration of ATP is as low as 1 μmol/L.

|
Download:
|
| Fig. 1. The structures of fluorescent probes for recognition of ATP based on Zn(Ⅱ)-DPA complexes. | |
{kind=link}
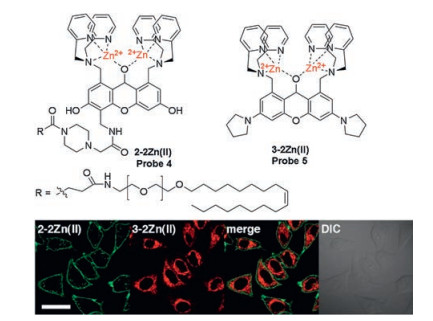
Hamachi's group has made excellent achievements in the design of ATP fluorescent probes for in vitro and cellular assays. Based on their previous binuclear zinc complexes as ATP recognition sites [34], they reported two new ATP ratiometric probes 2, 3 (Fig. 1, Scheme 1) based on fluorescence resonance energy transfer (FRET) mechanism, in which coumarin and xanthene are used as the donor and acceptor, respectively [35]. Both probes 2 and 3 have an obvious ratio response at the twoemission peak (R = Fxanthene/Fcoumarin), and the coordination between ATP and binuclear zinc complexes of the probes exhibits high FRET efficiency, 76% and 83%, respectively. These probes display significant dual-emission signal changes upon binding to ATP with strong affinity (Kapp ≈ 106-107 L/mol), which is amenable for accurate monitoring of ATP levels in living cells and various in vitro enzymatic reactions, such as the glycosyltransferase. By using the same respond mechanism, two new fluorescent probes 2-2Zn(Ⅱ) and 3-2Zn(Ⅱ) (probes 4, 5, Fig. 2) were synthesized and designed by the same group in 2012 to detect and multicolor display the dynamic changes of ATP in specific regions of living cells, such as plasma membrane and mitochondria [36].

|
Download:
|
| Scheme 1. The schematic illustration of the mechanism of probe 3 for recognition of ATP based on binuclear Zn(Ⅱ)-DPA complexes. | |
{kind=link}

|
Download:
|
| Fig. 2. Top: Molecular structure of 2-2Zn(Ⅱ) and 3-2Zn(Ⅱ) (probes 4 and 5). Bottom: Confocal micrographs of HeLa cells stained with 2-2Zn(Ⅱ) and 3-2Zn(Ⅱ). Scale bars: 20 μm. Copied with permission [36]. Copyright 2012, American Chemical Society. | |
{kind=link}
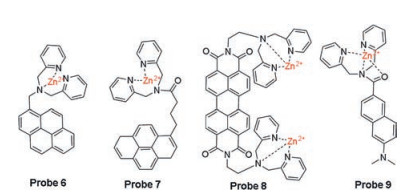
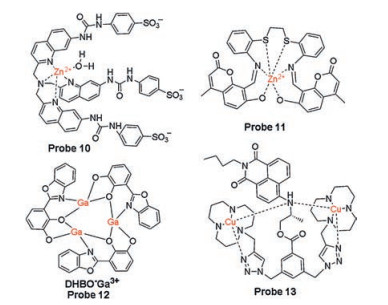
Based on the similar mechanism, two Zn(Ⅱ)-DPA complexbased pyrene excimers probes 6 and 7 (Fig. 3) were developed [37, 38]. Probe 6 has high selectivity to detect and distinguish ATP and UTP from other nucleotides in the solution of DMSO: HEPES (0.01 mol/L, pH 7.4, 90:10, v/v). Due to the length of linker between DAP-Zn complex and the chromophore pyrene, probe 7 is more selective for ATP than probe 6 and exhibits ratiometric fluorescence after responding to the nucleotide. Yan et al. reported a new perylene diimide skeleton probe 8 (Fig. 3) containing a Zn(Ⅱ)-DPA complex for specific detection of ATP rather than other phosphates, showing significant reduced absorption and considerable fluorescence enhancement [39]. For the advantageous features of two-photon microscopy (TPM) in biological applications, such as near infrared excitation and deep penetration depth, a two-photon fluorophore acedan functioned with a Zn(Ⅱ)-DPA complex is chosen to design a novel two-photon probe 9 (Fig. 3) for ATP detection [40]. Other Zn(Ⅱ) complexes with good ATP binding capacity were also selected for the development of ATP recognition sites for fluorescence probes 10 and 11 (Fig. 4), such as a hexadentate ligand-Zn(Ⅱ) complex based on thioether schiff base or a tripodal quinoline ligand-Zn(Ⅱ) complex [41, 42].

|
Download:
|
| Fig. 3. The structures of fluorescent probes for recognition of ATP based on Zn(Ⅱ)-DPA complexes. | |
{kind=link}

|
Download:
|
| Fig. 4. The structures of fluorescent probes for recognition of ATP based on other metal complexes. | |
{kind=link}
Although numerous of zinc(Ⅱ) complexes have been applied to the recognition of ATP based on electrostatic interaction, other metal complexes that with good affinity for triphosphate of ATP provide a brand-new approach to the design of ATP probes. According to their previous report, Ga3+ ions are easily coordinated with oxygen atoms on ligands, such as hydroxyphenyl structure. Xu et al. developed a new Ga(Ⅲ) complex as ATP probe 12 (Fig. 4) in 2015 [43]. This probe with a structure of a multi-membered array assembled between DHBO and Ga3+ ions, exhibits higher selectivity towards ATP with a fluorescence enhancement from other nucleoside in vitro or in vivo. The high selectivity of probe 12 for ATP over other nucleoside triphosphates (GTP, UTP, CTP) can be explained by two effects, including stronger electrostatic interaction between positively charged Ga3+ and phosphate group as well as the π-π stacking interaction between DHBO ligand and adenosine.
Interestingly, a Cu(Ⅱ)-complex based fluorescent probe 13 (Fig. 4) was reported by Lu et al. for detection of ATP in 2016 [44]. The addition of ATP to the probe solution generated a remarkable increase in fluorescence at physiological pH, while negligible fluorescence enhancement was observed in the presence of GTP and CTP. As described in literatures, a distinct difference between Cu(Ⅱ)-complex and Zn(Ⅱ)-complex based ATP fluorescent probes is ascribed to the concerted coordination of the triphosphate and adenosine segment of ATP to copper(Ⅱ) ions, which would be the decisive factors that determine the higher selectivity of Cu(Ⅱ)-complex based ATP probes.
Fluorescent probes based on metal complexes to recognize ATP have some drawbacks because these metal complexes may be toxic and non-metabolizable, or have significant suspicion with several pathological disorders, resulting in the rapid development of metal-free positively charged probes for specific binding phosphate anions of ATP. Li et al. synthesized a new metal-free probe DPTB-IMI-EG (probe 14, Fig. 5) with a electropositive di(1H-imidazol-1-yl)methane dication (IMI) as the triphosphate recognition site [45]. The probe exhibited specificity for ATP over ADP, AMP and other analytes. In the presence of ATP in an aqueous milieu, the electrostatic interaction between the probe and phosphate resulted in the formation of aggregates due to hydrophobic interaction, which gives DPTB-IMI-EG a considerable enhancementof photostability during the recognition process.Owing to the great improvement of photostability, this probe was successfully adopted to monitor and track the change and distribution of ATP with apyrase in NIH/3T3 fibroblast cells, which is a preferable imaging agent for cell imaging in confocal fluorescence microscopy and fluorescence lifetime imaging microscopy.

|
Download:
|
| Fig. 5. Top: Molecular structure of DPTB-IMI-EG (probe 14). Bottom: Confocal fluorescence images of live NIH/3T3 cells incubated with (A2-D2) or without (A1-D1) apyrase (1 U/mL) in DMEM for 60 min at 37 ℃ and then incubated with DPTB-IMI-EG and MDRF. (A) Green channel for DPTB-IMI-EG (probe 14). (B) Red channel for MDRF. (C) Panels A and B merged. (D) Optical image. Reproduced with permission [45]. Copyright 2014, Wiley Publishing Group. | |
{kind=link}
Ammonium is another candidate that can form a strong complex with the triphosphate segment of ATP. Bojtár and coworkers reported an ammonium modified pillararene complex DSS-AP6 (probe 15, Fig. 6) based on the fluorescent indicator displacement (FID) system to selectively recognize ATP by replacing with another competitive analyte [46]. During the recognition process, the chromophore dapoxyl sodium sulfonate (DSS) in the complex of ammonium-pillar [6] arene is replaced by ATP by electrostatic interaction, which contributes to the expected fluorescence quenching. Since the charge and length of the phosphate chains of ATP are most suitable compared to GTP, the probe exhibits a large signal ratio and a selective response to ATP among other nucleotides. An artificial nucleotide-binding peptide based naphthalimide fluorophore is utilized to design a biomolecule-based probe 16 (Fig. 7) for the detection of ATP, wherein a guanidiniocarbonyl pyrrole moiety can bind to triphosphate through electrostatic interactions and π-π interaction between the tryptophan moiety and adenine [47]. Successful application for intracellular ATP detection was also demonstrated by cellular imaging experiments.

|
Download:
|
| Fig. 6. The structures of DSS-AP6 (probe 15) for recognition of ATP based on the fluorescent indicator displacement (FID) system. | |
{kind=link}

|
Download:
|
| Fig. 7. Top: Molecular structure of peptidic probe 16. Bottom: Schematic representation for detection of ATP. Reproduced with permission [47]. Copyright 2017, Royal Society of Chemistry. | |
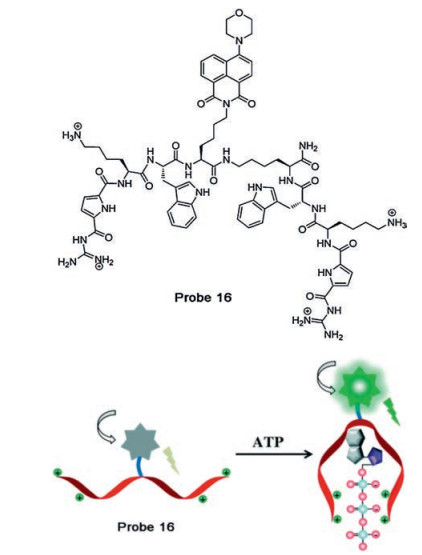{kind=link}
Along with the rapid interest in the use of aggregation-induced emission (AIE) probes, numerous examples have been reported to monitor reactive small molecules, enzyme, protein, DNA. Noguchi et al. developed a fluorescent probe 17 (Fig. 8) based on guanidinium modified tetraphenylethene (TPE) to discriminate ATP from ADP and AMP [48]. A 60-fold enhancement of blue emission was observed when ATP was added. During the recognition process, the positive and negative electrostatic interaction between guanidinium and phosphate anions contributes to the aggregation of TPE and the high signal-to-background ratio. Inspired by their seminal work, a TPE derivative TPPTPE (probe 18, Fig. 8) functioned with four pyridinium group was reported for the detection of ATP with a turn-on fluorescence signal in aqueous solution [49]. With excellent AIE properties, the probe was applied to monitor the activity of apyrase associated with ATP and image ATP in normal and cancer cells. Probe 19 (Fig. 8) is based on a thiazole orange derivative, another type of AIE molecule, which was reported by Deng and coworkers to show a turn-on fluorescent signal to ATP [50].

|
Download:
|
| Fig. 8. The structures of AIE fluorescent probes for recognition of ATP based on positively charged receptors. | |
{kind=link}
3.1.2. Recognition of ATP based on hydrogen bonds and π-π stacking interaction
Many fluorescent probes bearing amide, urea and azacrown have been invented because they can bind to phosphate anion by forming hydrogen bonds [9]. However, recognizing a nucleotide only by one hydrogen bonds interaction remains a huge challenge, especially in terms of selectivity. To the best of our knowledge, non-covalent interactions of aromatic rings have long been discovered and demonstrated in biological analysis, such as studies of the interactions of protein-protein and protein-receptor. Compared to other bases in the nucleotide, purines base more preferably forms stacking interaction with aromatic compounds, which dates back to the study of proteins and ATP complexes in 1996 [51]. Based on experimental and theoretical studies, Kataev et al. found that adenine and guanine have π-π interaction during nucleotide recognition. In addition, the cation-π interaction is unique to adenine [52]. Therefore, a number of ATP fluorescent probes based on hydrogen bonding interactions and unique stacking interaction have been developed and reported with intrinsic cationic chromophores, such as rhodamine B and rhodamine 6 G.
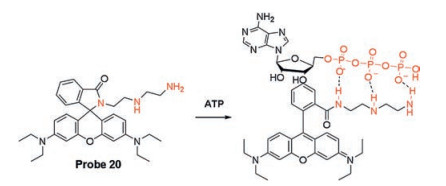
Based on the strategy of regulating fluorescence of rhodamine B via ring-opening of spirolactam, Li's research group designed and synthesized probe 20 (Scheme 2) using the polyamine groups to form hydrogen bonds with polyphosphates of ATP [53]. It was noted that the zero-background signal of probe itself due to the spirolactam structure. A remarkable enhancement of rhodamine B emission was observed in the presence of ATP, indicating that triphosphate of ATP promote the transformation of ring-opened amide from the spirolactam moiety. Meantime, probe 20 showed good selectivity for ATP rather than other phosphates due to π-π stacking interaction between adenine and xanthene. One year later, the same group reported a novel ratiometric fluorescent probe NR (probe 21, Fig. 9), a naphthalimide-rhodamine B compound for ATP detection based on the same strategy [54]. With the addition of ATP, the emission intensity ratio (I580/I530) of NR gradually increased with a large signal-to-background ratio of 31.8. The distance between ATP and NR was calculated with density functional theory (DFT) by Gaussian 03 package, indicating that there are dual interactions: hydrogen bonding interactions and π-π stacking interaction between them.

|
Download:
|
| Scheme 2. The possible mechanism of probe 20 for recognition of ATP based on hydrogen bonding and π-π stacking interactions. | |
{kind=link}

|
Download:
|
| Fig. 9. The structures of fluorescent probes for recognition of ATP based on diethylenetriamine as the reaction site. | |
{kind=link}
Similarly, Fang and coworkers reported a novel versatile fluorescent probe 22 (Fig. 9) for simultaneous detection of ATP and nitroreductase (NTR) with different responses, utilizing diethylenetriamine as the reaction site for ATP [55]. The dualfunction probe 22 exhibits different fluorescence responses to NTR, ATP, and NTR/ATP by collecting two emission peaks and adjusting the excitation wavelengths. After the addition of increasing concentrations (0–5 mmol/L) of ATP, a strong fluorescence at 580 nm was observed upon excitation at 540 nm, resulting in nearly 110-fold enhancement. Importantly, probe 22 was employed to reveal a decrease in the level of ATP during hypoxia, suggesting a possible relationship between NTR and ATP in living cells under hypoxia conditions.
ATP, a universal energy supply in cells, primarily produced in the mitochondria. It has been reported that the concentrations of ATP in mitochondria ranges from 0 to 5 mmol/L, which is closely related to many mitochondrial diseases. Therefore, it is particularly important and meaningful to develop probes suitable for monitoring the fluctuations of ATP levels in mitochondria. In 2017, a mitochondrial-targeted fluorescent Mito-Rh (probe 23, Fig. 9) was designed and synthesized for ATP by Li group, in which triphenylphosphonium (TPP) was used as a mitochondrial targeting group and diethylenetriamine was used as a reaction site [56]. The probe has a detection range of 0.1–10 mmol/L, which closely matches the actual concentration of ATP in mitochondria in cells. Confocal microscopy studies illustrated that Mito-Rh localizes in mitochondria with a Pearson's colocalization coefficient 0.95 and can monitor changes of ATP concentrations in real time.
Although ATP is produced primarily in mitochondria, lysosomal ATP also plays distinct roles in various physiological processes, such as membrane fusion, immunogenic cell death and apoptosis. To address this issue, Ahn et al. designed a ratiometric two-photon probe Lyso-ATP (probe 24, Fig. 10) inspired by rhodamine-based ATP probes for the detection of lysosomal ATP with a ratiometric fluorescent response in 2018 [57]. To mimic the pH range of lysosomes (4.5–5.5), they introduced a BDOIPY dye at the end of the tetramine chain on rhodamine 6 G and only detected ATP in the acidic pH range of the lysosome. In addition, they used an apyrase, which can hydrolyze ATP, to confirm the reversibility of Lyso-ATP and ATP in vitro, suggesting that it can quantitatively track changes in ATP at the cellular level. Two-photon cell imaging experiments indicated that lysosomal ATP was successfully ratiometric imaged and quantified during lysosomal membrane fusion.

|
Download:
|
| Fig. 10. Top: Molecular structure of probe Lyso-ATP (24). Bottom: Two-photon live-cell images of lysosomes that undergo (a) 'kiss-and-run' and (b) 'full-collapse fusion' processes, processed with pixel-topixel ratio images containing the quantitative information on ATP, and schematics of the two processes. HeLa cells incubated with Lyso-ATP (10 μmol/L) were used. The values indicate the emission intensity ratio values (IYellow/IBlue). Reproduced with permission [57]. Copyright 2018, Wiley Publishing Group. | |
{kind=link}
In 2018, Yoon group presented a new class of fluorescent probes 25, 26 and 27 (Fig. 11) decorated with thiourea on the rhodamine B core as a new recognition site for ATP detection through hydrogen bond and π-π stacking interaction [58]. Probe 25 exhibited 65-fold enhancement of fluorescence and a large binding constant of 5967 L/mol in vitro, and probe 26 was applied to monitor ATP in mitochondria in living cells. However, since the electron withdrawing group weakens the hydrogen bonding interactions, probe 27 showed poor sensitivity to ATP. The same research group later reported two additional ATP probes 28, 29 (Fig. 11) were subsequently reported by the same research team to selectively localize and detect ATP in mitochondria, lysosome, respectively [59].

|
Download:
|
| Fig. 11. The structures of fluorescent probes for recognition of ATP based on thiourea as the reaction site. | |
{kind=link}
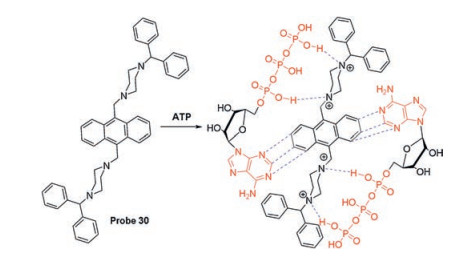
In addition to rhodamine B-based fluorescent probes, other probes have been designed to sense ATP based on hydrogen bonding and π-π stacking interaction. Misra et al. developed probe 30 (Scheme 3) to recognize ATP in live cells and aqueous medium [60]. A possible response mechanism is the concerted coordination of hydrogen bonds between triphosphate and piperazine resides and the π-π stacking interaction between anthracene and adenine.

|
Download:
|
| Scheme 3. The schematic illustration of the mechanism of probe 30 for recognition of ATP based on hydrogen bonding and π-π stacking interaction. | |
{kind=link}
3.1.3. Recognition of ATP based on synergistic interactions by multiple ways including covalent binding
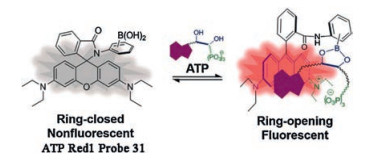
Given that the current fashion mechanisms for ATP recognition probes are mainly based on triphosphates and adenines in ATP structures, many probes limite the detection of ATP in living cells due to the challenges of selectivity and response range. In 2016, our group and collaborators designed and synthesized the probe ATP-Red 1 (probe 31, Fig. 12), through a multisite binding based on the vicinal diol of the pentose, adenine, and phosphates in the structure of ATP, which was the first proposed design ATP probe with covalent bond binding [61]. Probe ATP-Red 1 was prepared by introducing o-aminophenylboronic acid on the structure of rhodamine B and produced strong fluorescence in the presence of ATP, which was attributed to the synergistic interactions in multiple ways, including the covalent binding between the vicinal diol of a pentose and boronic acid, π-π stacking interaction between adenine and xanthene and the electrostatic interaction of triphosphate and amino. The absorption and emission spectra of ATP-Red 1 increase rapidly with titration of ATP from 0.5 mmol/L to 10 mmol/L, matching the intracellular ATP concentration range. With the selective and rapid response to ATP in vitro, ATP-Red 1 was successfully applied to monitor changes of ATP in mitochondria. Moreover, the applicability of probe ATP-Red 1 was verified by Qiao and co-workers for mitochondrial ATP detection and imaging [62].

|
Download:
|
| Fig. 12. The structure of ATP-Red1 (probe 31) and proposed mechanism for ATP sensing. Reproduced with permission [61]. Copyright 2016, Wiley Publishing Group. | |
{kind=link}
A similar strategy was adopted later by Xu et al. to report a mitochondria-targeted turn-on fluorescent probe Rh6 G-NH-PBA (probe 32, Fig. 13) to monitor ATP in living cells [63]. This probe utilizes rhodamine 6 G as a mitochondrial targeting fluorophore, and diethylenetriamine and phenylboronic acid (PBA) as recognition sites. The enhancement of fluorescence intensity of probe 32 toward ATP was observed to be approximately 51-fold in water at 1.0 mmol/L, while a 38-fold response to ADP was observed under the same condition. Rh6 G-NH-PBA was selectively located in mitochondria and displayed a decrease in red fluorescence in A375 cells when treated with apyrase. In addition to rhodamine B and rhodamine 6 G, some chromophores bearing boronic acid groups were also used for the detection of ATP in living cells. Ma et al. used this approach to design a new kind of AIE-active fluorescent probes TPA-PPA-1, TPA-PPA-2, TPA-PPA-3 (probes 33, 34, 35, Fig. 13) [64]. Both probes exhibited weak intramolecular charge transfer emission due to hydrogen bonding or proximityinduced quenching by aqueous molecules. Among them, TPA-PPA-3 showed low detection concentration and high selectivity towards ATP, as more binding sites resulted in more intramolecular rotations restriction. The application of TPA-PPA-3 for detection of ATP in HepG-2 cells was successfully carried out.

|
Download:
|
| Fig. 13. The structures of fluorescent probes for recognition of ATP based on synergistic interactions by multiple ways including covalent binding. | |
{kind=link}
3.2. Fluorescent probes for GTP
Besides ATP, GTP acts as a substrate for RNA synthesis, it is not only a vital kind of nucleotide, but also plays a critical role in providing energy source for protein synthesis, citric acid cycling and other metabolic processes [29]. In addition, the level of GTP in cells is closely related to the pathological state [30], thereby triggering the development of fluorescent probes to monitor it. The design theory of most fluorescent probes is based on guanosine base and triphosphate residue in GTP structure. On the one hand, it has been found that the ionic hydrogen bonding interaction between triphosphate and various receptors such as imidazolium, pyridinium is essential in the recognition process. On the other hand, the stacking interaction between guanosine and aromatic residue, the good electron-donating properties and the steric effect of guanosine give rise to the exceptional high quenching efficiency of fluorescence.
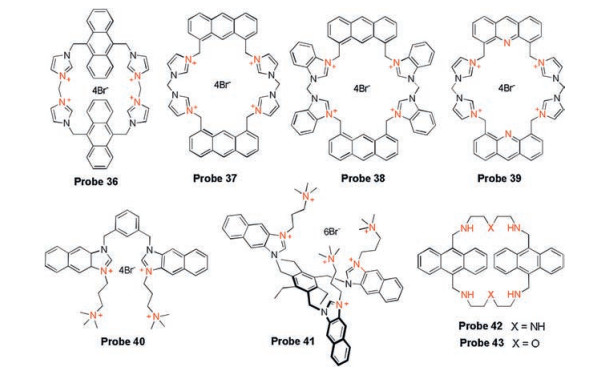
Based on the research of recognition mechanism through spectroscopy and NMR analysis as well as theoretical calculation, Kim group developed a series of fluorescent probes 36-39 (Fig. 14) for in vitro detection of GTP [65-68]. Probe 36 prepared by four imidazolium units, one anthracene and one cyclophane, showed monomer and excimer fluorescence emissions at 427 nm and 525 nm, respectively. Upon reaction with GTP, the fluorescence was significantly turned off due to the ionic hydrogen bond interactions between the GTP phosphate group and imidazolium protons, resulting in the separation of two anthracene excimers. Based on the same recognition mechanism, the same group developed probe 37 by modifying the imidazolium salt at different positions on anthracene, which is more selective for GTP than probe 36. They latter synthesized probe 38 based on benzimidazolium-anthracene cyclophane, showing effective fluorescence quenching effect for GTP in a physiological pH 7.4 aqueous solution. In 2014, anacridine fluorophore (probe 39) bearing animidazolium moiety was reported for GTP detection. In the presence of GTP, approximately 87% of the fluorescence was quenched, showing a higher quenching efficiency than probes 36-38 due to the aid of the electron-withdrawing nitrogen heteroatom of acridine in the ionic H-bonding interaction.

|
Download:
|
| Fig. 14. The structures of fluorescent probes for recognition of GTP. | |
{kind=link}
Several other research groups have also investigated GTP probes. Yoon group reported two probes 40, 41 (Fig. 14) based on bis- and tris-naphthoimidazolium derivatives [69]. Both probes exhibited a selective fluorescence quenching effect with GTP, which was attributed to the ionic hydrogen bonding interactions from imidazolium and phosphate groups. Kataev's group developed two probes 42, 43 (Fig. 14) based on anthracene modified with polycyclic amine, showing a turn-off response toward GTP [70]. You' group integrated a squaraines fluorophore with an bis-imidazolium-containing aniline derivatives to provide a sensitive colorimetric and fluorescent probe ImSQ8 (probe 44, Fig. 15) for GTP [71]. ImSQ8 possessing good solubility showed strong absorption and emission in HEPES buffer, while absorption and fluorescence spectra were reduced in the presence of GTP. This can be explained by the electrostatic interaction and ionic hydrogen bonding interactions between triphosphate and imidazolium by the aggregation of the dye on the GTP template, which forms the ImSQ8 and GTP host-guest model. And this hostguest interaction is used to detect alkaline phosphatase (ALP) by restoring color and fluorescence upon addition of ALP.

|
Download:
|
| Fig. 15. Proposed model of ImSQ88 (probe 44) for sensing GTP based on the hostguest interaction. Copied with permission [71]. Copyright 2014, Royal Society of Chemistry. | |
{kind=link}
In view of the fact that most GTP probes are based on turn-off fluorescence, Neelakandan et al. reported a novel cyclophanes CP-1 (probe 45, Fig. 16) containing a viologen receptor for selective turnon detection of GTP [72]. Based on the FID assay, CP-1 was combined with the fluorescent indicator 8-hydroxy-1, 3, 6-pyrene trisulfonate (HPTS) to construct the non-fluorescent complexes [CP-1·HPTS] for the determination of GTP with a 150-fold increase of HPTS fluorescence. The mechanism behind the recognition can be attributed to the concerted coordination of electrostatic interaction and π-π stacking interaction. A similar strategy was adopted by Karunakaran and coworkers to develop a fluorescent turn-on probe PyP·HPTS (probe 46, Fig. 16) to discriminate GTP from ATP in 2014 [73]. The complex [PyP-HPTS] was formed by complexation of the cationic porphyrins PyP with negatively charged HPTS, showing the quenching emission of HPTS from PyP. After the introduction of GTP, a remarkable green fluorescence corresponding to HPTS was observed.

|
Download:
|
| Fig. 16. The structures of fluorescent probes for recognition of GTP based on cationic receptors pyridinium. | |
{kind=link}
3.3. Fluorescent probes for ADP
ADP is one of the hydrolysis products of ATP and plays pivotal roles in oxidative phosphorylation, energy supply and metabolic inter-conversions. ADP is closely related to the significant biological reaction catalysed by ATPase and kinases [30]. In view of the biological importance of ADP, efforts have been made to design fluorescent probes to detect and discriminate it from ATP. In 2013, Feng's group found that mononuclear zinc complexes can be used as recognition sites for ADP detection. They developed a probe ZnL (probe 47, Fig. 17), appended two anthracene chromophores, showing a unique increased fluorescence response to ADP [74]. The unique selectivity of ZnL for ADP plays a synergistic role in the electrostatic interaction between Zn(Ⅱ)- diphosphate and the π-π stacking between the anthracene and adenine. Later, they reported another probe 1-2Zn (probe 48, Fig. 17), which has a binuclear zinc macrocyclic complex for recognizing ADP from other structurally similar phosphates, especially ATP and inorganic pyrophosphate (PPi) [75]. In view of the lack of application in cells of these two probes, probe 49 (Fig. 17) with a binuclear Zn(Ⅱ)-DPA complex and four NH2 groups was developed to detect ADP in living cells by the same group [76]. In the presence of ADP, a 133-fold fluorescence enhancement was observed, which was much higher than ATP (56-fold) and AMP (3.9-fold). It is attributed to multiple interactions, including coordination of metal-anion, π-π stacking interaction and hydrogen bonding interactions, where the hydrogen binding reduces the PET effect of the NH2 groups. Encouraged by their work, Lu's group reported an anthracene-based probe 50 (Fig. 17) for ADP detection with a similar response to ATP [77].

|
Download:
|
| Fig. 17. The structures of fluorescent probes for recognition of ADP based on zinc complexes and imidazolium. | |
{kind=link}
Duan's group reported a new recognition site, imidazolium podands for the design of ADP probes. They combined the naphthalimide dye and imidazolium moiety through a tripodral podands to construct probe TIA1 (probe 51, Fig. 17), which has high selectivity for fluorescence detection of ADP [78]. Upon the introduce of ADP, no change was observed in the absorbance spectrum, while a significant increase of fluorescence intensity was observed due to the PET effect. The hydrogen bonding interactions and the suitable lengths of the linker may be responsible for its selectivity to ADP over ATP and other nucleotides.
3.4. Fluorescent probes for other nucleotidesUTP and TTP are two nucleotides of triphosphate, which are key building blocks for the synthesis of RNA and DNA synthesis in vivo and are involved in various biological processes such as sugar and metabolic conversions. Seiji Shinkai's group reported probe PBG (probe 52, Fig. 18) decorated with a guanidinium-perylene based symmetric molecular receptor [79]. When UTP is added, the fluorescence of PBG is dramatic decreased due to ionic hydrogen bonding interactions between guanidinium and phosphate groups in UTP, resulting in the aggregation of perylene moieties. And probe PBG exhibited a high selectivity to UTP over other nucleotides, including CTP, GTP, ATP, UDP and UMP. In 2018, Kataev's group developed amacrocyclic-based TTP probe 53 (Fig. 18) with a turn-on fluorescence response [70]. Due to the elongated linker and the polycyclic amine on the macrocycle, probe 53 showed about 80% fluorescence increase compared to the other nucleotides.

|
Download:
|
| Fig. 18. The structures of fluorescent probes for recognition of other nucleotides. | |
{kind=link}
It is well known that many fluorescent probes for Hg2+ detection are based on high binding affinity between ligand thymidine and Hg2+ [80]. Inspired by this specific interaction, Chang and coworkers designed an Hg(Ⅱ) complex (probe 54, Fig. 19), a fluorescent probe for the detection of thymidine nucleotides (TTP, TDP, TMP) [81]. They synthesized a fluorophore with a thymine-like structure with high quantum yield. And the fluorescence of this compound was quenched by the formation of Hg(Ⅱ) complex. After the addition of thymidine polyphosphates, the fluorescence was rapidly restored by ligand exchange between thymidine and thymine analogue fluorophore, achieving the goal of selectively detecting thymidine nucleotides over other nucleotides.

|
Download:
|
| Fig. 19. Schematic illustration of the proposed mechanism of probe 54 for detection of thymidine derivatives. Reproduced with permission [81]. Copyright 2015, Elsevier Ltd. | |
{kind=link}
Monophosphate nucleotides (UMP, AMP, cAMP, GMP, TMP) are the fundamental units of nucleic acids synthesis and are the primary products of tri-/di-phosphate nucleotides, which participate in and regulate considerable physiological processes, such as metabolic processes, signal transduction and gene expression [30].
By utilizing the principle of complementary base pairing, Piantanida's group reported a novel UMP probe 55 (Fig. 20) based on bis-phenanthridinium adenine conjugates, taking advantage of recognizing a complementary nucleotide through hydrogen bonding from the nucleobase positioned within the hydrophobic cavity [82]. Upon addition of UMP, the fluorescence intensity of probe 55 declined gradually, which was the result of multiple interactions with UMP, including inter- and intra-molecular stacking interactions along with intermolecular hydrogen bonds between the probe and uracil. In addition, the selective recognition of UMP can be explained by the higher affinity of probe 55 for UMP over other nucleotide mono-phosphates.

|
Download:
|
| Fig. 20. The structures of fluorescent probes for recognition of monophosphate nucleotides. | |
{kind=link}
In 2013, Ghosh et al. established an ensemble (probe 56, Fig. 20) of pyridinium receptor and fluorescein dye for the selective detection of AMP over ATP or ADP by using FID assay. As an example, they utilized this probe to monitor AMP in A549 cell line [83]. In 2017, Sahoo and coworkers reported a new naphthol-based Rhodamine derivative NpRD (probe 57, Fig. 20) for the detection of adenosine 3', 5'-cyclic monophosphate (cAMP) with turn-on yellow fluorescence at 562 nm [84]. NpRD itself had no emission, and generated a strong fluorescence in the presence of AMP due to the opening of the spirolactam ring, which was attributed to the synergistic coordination of hydrogen bonding interactions and π-π stacking interaction. Similarly, this synergistic coordination was responsible for NpRD selectivity and differential detection of cAMP over ATP, GTP, GMP, CTP, UTP, TTP, inorganic phosphates and other anions. Moreover, NpRD was applied to measure cAMP in living cells and in human blood samples.
A new naphthyridine-based Gd(Ⅲ) complex Gd-ANAMD (probe 58, Fig. 20) was established by Duan's group for the detection of GMP in aqueous media and in vivo [85]. When GMP was introduced to the complex, strong multi-hydrogen bonds between naphthyridine and guanosine formed, resulting in coordination between monophosphate residue and lanthanide ion and the changes in fluorescence. In addition, Gd-ANAMD was used as a magnetic resonance imaging (MRI) agent due to the characteristics of the lanthanide metal. Using the same naphthyridine motif, Fang's group devised a GMP probe 59 (Fig. 20) bearing a guanidine group through the strong electrostatic interaction between the positively charged guanidinium and negatively charged monophosphate in an aqueous system [86]. In 2014, Zhao and coworkers reported a colorimetric and fluorescent probe 60 (Fig. 20) based on 1, 8-naphthalimide-based analogs for the selective detection of three mono-phosphates GMP, TMP, and UMP over other nucleotides [87]. And probe 60 was employed to detect these three mono-phosphates in C. elegans, respectively.
3.5. Challenges for detection of nucleotidesWhile using multiple strategies including electronic interaction, hydrogen bonding and stack interaction to design satisfactory probes, there are some challenges that need to be broken through: (1) Although improved selectivity is achieved by various approaches, the selectivity of probes are limited to recognize special nucleotide in living cells or biological environments due to various diversities and structural similarities, especially for nucleotides with low content. Moreover, several nucleotide probes have not received enough attentions, such as GTP, GDP, UDP, TDP and cGMP. (2) Although most probes based on metal complexes or positively charged ligands have been applied to monitor and image nucleotides in cells, their utility and toxicity remain problematic, such as applications in long-term cellular imaging experiments. In addition, the introduction of large amounts of positive charge into cells can disturb the cell balance, alter the cell membrane potential, and even rupture the cells [88, 89]. (3) On the one hand, most of the reported nucleotide probes are based on on-off of the fluorescence intensity of a single channel, which will be easily affected by a variety of factors, including probe concentrations, instrumental apparatus, and interference background. On the other hand, these reported probes have emission wavelengths mainly in the blue or visible regions, thus limiting their use for cells or in vivo due to the poor penetration depth and background fluorescence. Future research will focus on the development of ratiometric probes based on two-photon or near-infrared fluorescent probes. (4) Most of the reported probes have only used for in vitro spectral detection, but their detection limits and response concentration ranges do not match intracellular concentrations of nucleotides. Therefore, the concentration ranges and microenvironment within the cells should be considered in the future development of nucleotide probes.
4. Fluorescent probes for NAD(P)H and FAD (FMN)Nicotinamide adenine dinucleotide (NADH) and flavin adenine dinucleotide (FAD), the members of adenine nucleotides, are important coenzymes with unique biological functions and significant signal modulators [5]. Their structure consists of adenine moiety and ribosylated nicotinamide dinucleotide (NAD) or ribitolylated flavin adenine dinucleotide (FAD) via a pyrophosphate linking group. Among the nicotinamide adenine dinucleotide derivatives, two major categories have attracted widespread attention, that is, NADH and its phosphate form NADPH, and serve as two important coenzymes in various physiological activities, such as enzymatic reactions, cellular redox reactions and metabolic reactions [90]. Flavin adenine dinucleotide (FAD), flavin mononucleotide (FMN), riboflavin (RF) are the core members of the flavins family and play crucial roles in energy metabolism and redox balance, especially in biological electron transport process [91]. Given the critical importance of these molecules and related significant physiological events, there is an urgent need to design efficient fluorescent probes for their monitoring and imaging.
4.1. Fluorescent probes for NAD(P)HNAD derivatives (NADH, NADPH), electron and hydrogen atom carriers, are involved in many biological processes, including redox balance regulation, cellular energy metabolism, tricarboxylic acid (TCA) cycle, mitochondrial functions and immunological functions [90]. Recent studies have shown that their concentration levels are closely related to several diseases, such as Parkinson's disease, diabetes and cancer [92]. In addition, they are reported to be therapeutic agents for neurodegeneration treatment [93]. Based on their unique redox characteristics, a few fluorescent probes have been devised for their detection. Besides, several recognition mechanisms have been used, including hydrogen bonding interactions between phosphate groups and probes, metal-anion interactions, and combinations of pentoses with boric acid of probes.
In 2013, Komatsu and coworkers developed the first fluorescent probe UQ-Rh (probe 61, Scheme 4) for the detection of intracellular NAD(P)H using ubiquinone derivative as a new reaction site and rhodol as the fluorophore [94]. Based on the redox reaction between the quinone and NAD(P)H, it was observed that the intensity of UQ-Rh was reduced by the reduction product hydroquinone due to the PET mechanism. Significantly, a better response was observed after introducing a promoter of NAD(P)H (an artificial Ir complex) into the reaction system. As an example, this reaction systemwas successfully employed to detect the levels of NAD(P)H in HeLa cells.

|
Download:
|
| Scheme 4. The structures of fluorescent probes for recognition of NAD(P)H based on the redox reactions. | |
{kind=link}
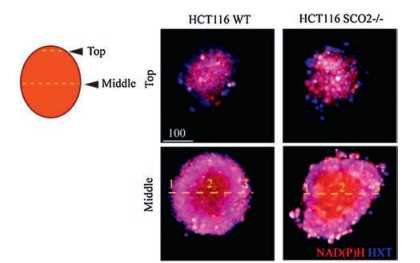
Inspired by the Alamar Blue bioassay for cell viability assessment, Fomin et al. reported a cyanine-based fluorescent probe 62 (Scheme 4) with a novel mechanism with two-acceptors [95]. A nearly 30-fold increase in fluorescence intensity was observed in the presence of NAD(P)H, due to the reduction of the quinolinium acceptors by NAD(P)H to dihydropyridine, forming a new π-conjugated donor-acceptor system of the cyanine dye. Probe 62 was applied to detect and image the generation of NAD(P) H, which assists with NAD(P)+ in vitro and in cells by glucose dehydrogenase (GDH) to oxidize glucose to gluconolactone. In particular, the authors demonstrated that the metabolic states were measured by visualizing the intracellular NAD(P)H in the hypoxic core of tumor spheroids with probe 62 (Fig. 21). Tang's group lateradopted the similar response site and developed a nearinfrared (NIR) emission fluorescent probe DCI-MQ (probe 63, Scheme 4) for the detection of NAD(P)H in vivo [96].

|
Download:
|
| Fig. 21. Fluorescence microscopy images NAD(P)H of different cross sections of spheroids stained with probe 62 (red) and marker of nuclei Hoechst 33, 342 (HXT, blue). Copied with permission [95]. Copyright 2016, American Chemical Society. | |
{kind=link}
In 2018, Mithun et al. developed a reversible fluorescent probe 64 (Scheme 4) based on benzo[a]phenoxazine dye for the detection of NAD(P)H for the first time [97]. When NAD(P)H was added, probe 64 was reduced and displayed a decrease in fluorescence intensity at 650 nm. Moreover, the original fluorescence of the probe itself was restored by the strong oxidant HClO, indicating the redox mechanism behind this reversible cycle.
It is well known that metal complexes and amide receptors are effective recognition sites for the recognition of phosphate compounds. A turn-off type probe FMA (probe 65, Fig. 22) based on fluorescein mercury acetate derivatives for the detection of NADH was reported by Han and coworkers, which depended on the metal-anion interactions formed by mercury and phosphate segments [98]. Due to these interactions, FMA exhibited a high selectivity for NADH over other biomolecules. Using a calix[4]arene derivative, a probe C4D (probe 66, Fig. 22) was designed for the detection of NADH in aqueous media, in which the three amide moieties act as receptors for binding to phosphate group of NADH [99]. The fluorescence intensity of C4D was observed to decrease significantly with increasing NADH, and the detection limit was 24.5 nmol/L. Moreover, the binding constant (Ka) of probe C4D to NADH was calculated to be 3.74 × 105 L/mol from the fluorescence titration spectra.

|
Download:
|
| Fig. 22. The structures of fluorescent probes for NAD(P)H based on other interactions. | |
{kind=link}
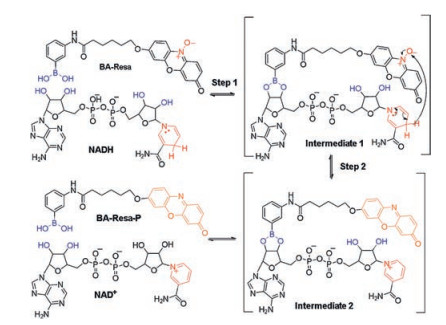
It is difficult to selectively detect NAD(P)H from a large number of reducing substances and phosphates in cells by only one kind of interaction. Inspired by the enzyme catalyzed reaction strategy, Chang's group designed two turn-on fluorescent probes BA-Resa and RA-Resa (probes 67, 68, Fig. 22) for sensing NADH in two steps: first shortening the distance between NADH and the probe through the binding of boronic acid and pentose, and then subjected to a redox reaction between them through a close spatial distance [100] (Scheme 5). This two-steps recognition process significantly increases the sensitivity and selectivity of the probe over NADPH and other phosphates, allowing BA-Resa to have a low detection limit of 0.087 μmol/L at low concentrations (0-1 μmol/L) for NADH. Since the pH condition of probe BA-Resa response was at 9.5, the probe RA-Resa was designed to have a better response to NADH under physiological conditions at pH 7.4, which was successfully applied to the measurement and imaging of NADH in living cells.

|
Download:
|
| Scheme 5. Proposed recognition mechanism of probe BA-Resa (probe 72) responding toward NADH. | |
{kind=link}
4.2. Fluorescent probes for FAD and FMN
FAD and FMN are essential coenzymes based on flavin in all eukaryotic enzyme catalysises. FAD is also a fundamental component of eosinophils and produces a series of reactive oxygen species, such as H2O2 and O2-, which produce phagocytosis [101]. FMN is a prosthetic group of various oxidoreductases involved in the process of oxidoreduction. Therefore, the biological concentration level of flavin is related to flavin-linked metabolism and various physiological symptoms [6, 91, 102]. Plenty of efforts by researchers have been devoted to the development of fluorescence detection systems for flavins. Both FAD and FMN structures contain an isoalloxazine ring with good fluorescence features, such as green emission and high quantum yield (Φ = 0.26) in water, which results in several fluorescence detection methods based on detection of changes in the isoalloxazine group. In addition, other strategies have been used to develop fluorescent probes for their recognition.
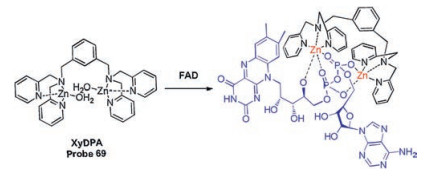
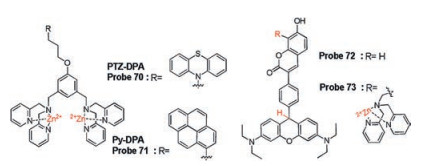
Continuous efforts of Hong's group have been devoted to developing and establishing fluorescence approaches for FAD and FMN. In the early stages, a new probe based on Zn(Ⅱ)-DPA complex, XyDPA (probe 69, Scheme 6), was reported to detect FAD due to strong metal-ionic interaction from diphosphate group of FAD and π-π stacking interaction between adenine and isoalloxazine ring [103]. And they found that XyDPA can split FAD into cFMN and AMP. Based on this result, they introduced a phenothiazine group into XyDPA to construct another probe PTZ-DPA (probe 70, Fig. 23), which can detect FAD by measuring the fluorescence of FMN due to the same splitting mechanism of XyDPA. Furthermore, due to the metal-ionic interaction of the mono-phosphate group of FMN and Zn complex, fluorescence quenching was observed by PET mechanism in the presence of FMN. These results enable PTZDPA to selectively distinguish between FMN and FAD through difference fluorescence changes. Probe PyDPA (probe 71, Fig. 23) with similar structure was employed to selectively distinguish NAD+/NADP+ from FAD/FMN through the changes of ratiometric fluorescence (Fexcimer/Fmonomer) and response time [104].

|
Download:
|
| Scheme 6. The recognition mechanism of XyDPA (probe 69) for FAD. | |
{kind=link}

|
Download:
|
| Fig. 23. The structures of fluorescent probes for recognition of FAD and FMN. | |
{kind=link}
Although autofluorescence of flavin is used to detect FAD and FMN, this approach is limited in living cells and tissues due to the short excitation and shallow penetration. The same group developed a new imaging method for flavin by photo-oxidation of probe 72 (Fig. 23) in 2016 [105]. Probe 72 was constructed by a bis(diethylamino)-[9H]xanthene ring and a coumarin chromophore via a phenyl linker, which exhibited remarkable new bands in both absorption and fluorescence spectra in the presence of 2.0 equivalents FMN upon photo-oxidation (365 nm, 6 W, 3 min). A pink color and red fluorescence can be explained by flavinmediated oxidation under UV light conditions. Due to this special reaction system, probe 72 exhibited high selectivity to FMN and a good detection limit of 5 × 10-7 mol/L. Confocal microscopy experiments demonstrated that probe 72 was capable of detecting mitochondrial flavin in cells and tissues. In order to significantly weaken the auto-oxidation of probe, the latter synthesized and developed probe 73 (Fig. 23) by modifying a Zn(Ⅱ)-DPA complex on coumarin segment [106]. Under ambient light conditions, the slowest rate of auto-oxidation was observed compared to probe 72 due to changes of surrounding electrostatic environment. Given the superior properties, probe 73 was successfully applied to eosinophil imaging and differential diagnosis of eosinophilia using human blood samples.
4.3. Challenges for detection of NAD(P)H and FAD (FMN)Although some progress has been made in identifying NAD(P)H and FAD (FMN) through smart strategies based on redox reactions or special interactions, there are still some challenges: (1) Only a few established probes were used for cell detection and imaging, while, most probes were not sufficient for practical applications in cellular environment, since their response concentration ranges and selectivity studies do not match intracellular concentrations. Faced with a large number of redox species in living organisms, such as oxidoreductase, small molecules and proteins with redox properties, it is necessary to enhance the redox mechanism based probe design by combining multiple methods, such as probe RA-Resa, preferably for the selective detection over other competitive interferent in living cells and in vivo. (2) The reaction processes involved in NAD(P)H and FAD (FMN) are dynamically changed in living cells, and only one reversible probe 64 has been reported in vitro, but cannot be used for monitoring intracellular NADH due to its poor sensitivity. Therefore, future efforts should focus on developing reversible probes to monitor their dynamics in living cells in real-time and selectively. (3) On the one hand, most synthesized probes are not able to quantify analyte levels, so it is necessary to develop ratiometric probes for better imaging results. On the other hand, due to the scarcity of recognition sites, many researchers are limited in developing fluorescent probes with excellent performance. Therefore, there is an urgent need to develop more recognition sites in future research. In addition to small molecule probes, advances in NAD(P)H and flavin derivatives have been made through genetically encoded probes and nanoprobes, but further research is needed. (4) Most reported probes are mainly used for NAD(P)H and FAD (FMN), but studies on other adenine nucleotides and dinucleotides have not received special attention, such as their derivatives NAD+/NAD(P)+.
5. Fluorescent probes for phospholipidsIn addition to phosphate-containing nucleotides, there is also a large class of organophosphates known as phospholipids, which are abundant and important in mammalian and bacterial membranes. The cell membrane composed of phospholipids separates intact cells from the external environment and maintains fundamental functions in such significant fields as membrane signal transduction, metabolism, and immune [107]. About 70% of membrane phospholipids are glycerophospholipids, which are very diverse due to cell types, different organelles and different types of inner and outer membranes [108, 109]. According to the polar heads on the glycerol backbone, glycerophospholipids mainly include six categories: phosphatidylserine (PS), phatidylcholine (PC), phosphatidylethanolamine (PE), phosphatidicacid (PA), phosphatidylglycerol (PG) and phospholipid cardiolipin (CL) [110].
5.1. Fluorescent probes for cell surface PSPS is the most abundant anionic phospholipid in the plasma membrane, and has become a hot research in the field of biochemistry due to its importance in the intracellular and extracellular context [111]. To the best of our knowledge, the extracellular membrane of healthy mammalian cells is electrically neutral because PS is exclusively on the inner membrane. However, PS is exposed to the cell exterior during cell death as a marker of early apoptosis [112]. In addition, the increase in negatively charged PS on the outer surface is also a marker of many physiological processes and disease states, such as hemostasis and phospholipidosis [113, 114]. Moreover, recent studies have shown that PS levels on cancer cell membrane are considered to be cancer therapeutic targets. Given the importance of PS in membranes, the lack of effective tools enables real-time monitoring of PS dynamics for studying cellular events and pathological states, triggering the rapid development of fluorescent probes for PS detection [115, 116]. In this part, we mainly focus on the design and recognition mechanism of PS probes, along with their biological applications. Considering the negative charge of PS, numerous fluorescent probes adopted metal complexes to devise recognition sites due to strong electrostatic interaction and other interactions.
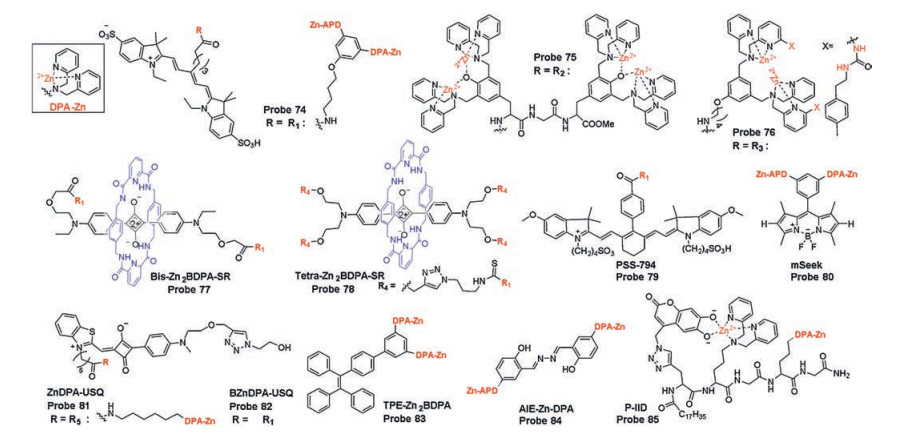
Based on their research of Zn(Ⅱ)-DPA complexes (Zn2BDPA) as PS probes in 2005, Smith's group developed two probes 74 and 75 (Fig. 24) based on CyAL-5 fluorophore and phenoxide-bridged Zn2BDPA structures. These probes selectively increase the target affinity of PS and reduce probe accumulation of unbound probes by enhancing the electrostatic interaction between PS and complex [117]. Among them, probe 74 showed better image contrast of dead and dying cells due to poor self-aggregation than probe 75. They also utilized a structural modification strategy to attach a urea functional group to Zn-BDPA core scaffold to get probe 76 (Fig. 24), to monitor PS by concerted coordination of electrostatic attraction, hydrogen bonding and hydrophobic insertion into membranes [118].

|
Download:
|
| Fig. 24. Fluorescent probes for recognition of PS based on Zn(Ⅱ)-DPA complexes. Inset: Structure of DPA-Zn complexes. | |
{kind=link}
Based on squaraine fluorophores, bis-Zn2BDPA-SR and tetraZn2BDPA-SR (probes 77, 78, Fig. 24) were developed from the same group for the detection of PS in different animal models with two or four appended Zn2BDPA units, respectively [119]. According to in vitro and in vivo studies, tetra-Zn2BDPA-SR (probe 78) displayed a higher selectivity for PS on the membrane exterior surface due to the high affinity between its large number of Zn2BDPA targeting group and negatively charged PS. Other probes PSS-794 and mSeek (probes 79, 80, Fig. 24) using Zn(Ⅱ)-DPA complexes as recognition group were developed and investigated by the same group for the PS monitoring and bacterial detection [120, 121]. In 2018, two probes ZnDPA-USQ and BZnDPA-USQ (probes 81, 82, Fig. 24) decorated with one or two Zn(Ⅱ)-DPA complexes on one solvatochromic squaraine dye were reported. Fluorescence and time-lapse microscopic imaging for the detection of PS exposed on the surface of dying cells were reported [122]. Compared with the previous reported PS probes, these two probes themselves exhibit weak fluorescence in bulk water due to self-aggregation, but the fluorescence intensity is significantly enhanced by a two-step binding process during the occurrence on membrane, that first releases free monomer and then binds to PS by electrostatic interaction. This way of interaction makes it possible with no wash before detection and imaging, providing a new idea for constructing PS activation probes.
Inspired by their groundbreaking work, Tang's group devised a PS probe TPE-Zn2BDPA (probe 83, Fig. 24)using tetraphenylethene (TPE) as the AIE reporter and Zn(Ⅱ)-DPA complexes as the recognition and targeting group [123]. TPE-Zn2BDPA can selectively detects and differentiates PS on the membrane surface in the early and late stages of apoptosis, so as to achieve the purpose of identifying different stages of cell apoptosis.
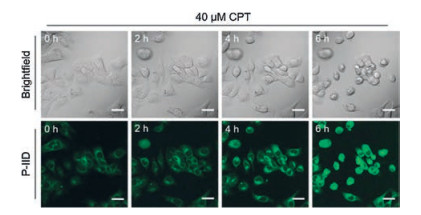
Similarly, a AIE fluorescent PS probe AIE-ZnDPA (probe 84, Fig. 24) with aggregation-induced emission and excited-state intramolecular proton transfer mechanism was reported by Liu's group, which was used to differentiate early and late stage of apoptosis [124]. In order to improve the high signal-to-noise ratio, Jolliffe and co-workers designed a new fluorescent PS probe P-IID (probe 85, Fig. 24) by intramolecular FID assay, which is displacement of two bindings: the binding of recognition group (Zn(Ⅱ)-DPA complexes) to the fluorescence indicator (6, 7-dihydroxycoumarin) and the analyte (cell surface PS) [125]. Due to the quenching effect, the probe itself has weak background fluorescence and shows a turn-on fluorescence in the presence of PS because of the strong interaction between Zn(Ⅱ)-DPA complex and the phosphate segment of PS. Cellular studies have shown that probe P-IID has rapid kinetics of binding to PS when labeling apoptotic cells without washing step, is more efficient, and avoids defects occurring on commercial probes AnV (Alexa Fluor 647). Time-lapse imaging experiments indicated that P-IID was successfully used for real-time imaging of cell apoptosis induced by drug camptothecin (CPT) (Fig. 25).

|
Download:
|
| Fig. 25. Time-lapse imaging of HCC-1806 cells treated with CPT (40 μmol/L) and stained with P-IID. Copied with permission [125]. Copyright 2018, Wiley Publishing Group. | |
{kind=link}
5.2. Challenges for detection of phospholipids
In addition to PS, detection of other phospholipid molecules (such as PC, PE, PA, PG, CL) based on probe design has rarely been reported, thus more effort is needed to develop probe designs. Overview of all reported small molecule fluorescent probes for PS detection, the recognition group based on Zn(Ⅱ)-DPA complexes limited the development of PS probe with higher performance, such as enhanced sensitivity and minimized background fluorescence of unbound probes [116]. For phosphates recognition, other sites have been widely used and reported, such as imidazolium, quaternary ammonium, amide and urea derivatives [8]. It is conceivable to combine these recognition sites with multiple interaction ways, such as hydrogen bonding, electrostatic interaction and covalent bonding. Despite various progresses achieved in the reported probes, these studies have focused on detecting PS on the cell surface during cell death with less interference because most phosphates is within cell. It is well known that PS is also exposed to the cell surface during blood clotting or other physiological processes, and these processes have not caused concern for research and testing. Unfortunately, there is no probe for quantitative detection and imaging of PS in cells. Ratiometric fluorogenic probes will be a good candidate. With the rapid development of organelle-targeting fluorescent probes [126], probes with an organelle membrane targeted group can be applied to distinguish and visualize various membranes surface PS.
Although several activated fluorescent probes are used for PS detection, the background fluorescence of probes cannot be eliminated, resulting a low signal-to-noise ratio. Numerous approaches based on zero background activation of fluorescent probes designs, such as Chang' group, developed an efficient background-free probe construction strategy in 2005 [127], which should be considered for PS detection in futurework. Sauer and coworkers reported a method that uses super-resolution imaging to visualize plasma membrane glycans and their distribution [128]. Therefore, visualization and analysis of the ultrastructural of PS can be achieved by using a fluorescent probe with a super-resolution fluorescence microscopy. To the best of our knowledge, probes can easily access the cell surface without the need of additional targeting strategy. If we want a more complete understanding of the distribution and function of cells under normal conditions, we need to develop probes to selectively detect native PS in cells. In addition tosmall-molecule probes for PS recognition, manystudies on fluorescent protein or genetically encoded sensors have recently been presented, but further investigations are need.
6. Summary and prospectsSmall fluorescent probes for the recognition of organic phosphates have achieved promising results in spectral analysis in vitro, cellular and intravital imaging. In this review, we have presented the progress of fluorescent probes for organic phosphates probes by our group and other groups from 2010 to the present, including probes for ATP, GTP, ADP, other nucleotides, NAD (P)H, FAD (FMN) and PS. We mainly summarize the general strategies for probe designs for selective and differential detection based on the structure of organic phosphates analytes, and achieve biometric information and tract the changes of their levels in different states. The challenges left by these advances point us to many future directions and efforts in designing and constructing organic phosphates probes: (1) Owing to the wide variety organic phosphates, selective detecting and differentiating them only throughsingle recognition site or mechanism is difficult toachieve. Our group or other groups utilized the synergistic interaction of multiple ways for the selective detection of ATP and NADH from other phosphates, which can be considered and adopted by researchers during the process of devising organic phosphates probes, and even other probes design. (2) Fluorescent probes and fluorescence imaging have been developed for decades and have been established for the detection and monitoringof various signal biomolecules in biological environments. For the design of organic phosphatesprobes, researchers can learn from the design of probes ROS/RNS/RSS, enzymes and complex molecules, such as utilizing different organelles, membranes, tumor targeting to detect different cellular surface PS; using weak acid and response sites for selective detection of organic phosphatesin lysosomalor tumor regions; making use of its special reaction and unique physiological functions for probes design. (3) Fluorescent probes based on fluorescence confocal imaging technique have not yet fully and factually obtained many biological informations, such as signaling pathways, dynamic changes and link related diseases at the cellular levels as well as intravital levels. Probes 58 and 14 combined the fluorescence imaging with MRI and fluorescence lifetime techniques, respectively. The fluorescent probes combined with multi-techniques can compensate for the defects of a certain technique and combine their respective advantages to obtain more comprehensive, complete and accurate information for us, which need to be exploited.
AcknowledgmentsThis work was financially supported by the National Natural Science Foundation of China (NSFC) (Nos. 21622504, 21735001, and 21877029), the Science and Technology Project of Hunan Province (No. 2017RS3019), and the Open Funding Project of the State Key Laboratory of Bioreactor Engineering.
| [1] |
C. Levinson, Biophys. Acta 120 (1966) 292-298. |
| [2] |
H.A. Lardy, H. Wellman, J. Biol. Chem. 195 (1952) 215-224. |
| [3] |
A.V. Gourine, E. Llaudet, N. Dale, et al., Nature 436 (2005) 108-111. DOI:10.1038/nature03690 |
| [4] |
A. Kornberg, CRC Crit. Rev. Biochem. 7 (1979) 23-43. DOI:10.3109/10409237909102568 |
| [5] |
A.A. Heikal, Biomark. Med. 4 (2010) 241-263. DOI:10.2217/bmm.10.1 |
| [6] |
C. Walsh, Acc. Chem. Res. 13 (1980) 148-155. DOI:10.1021/ar50149a004 |
| [7] |
T. Michigami, M. Kawai, M. Yamazaki, et al., Physiol. Rev. 98 (2018) 2317-2348. DOI:10.1152/physrev.00022.2017 |
| [8] |
A.M. Agafontsev, A. Ravi, T.A. Shumilova, et al., Chem.-Eur. J. 25 (2019) 2684-2694. |
| [9] |
A.E. Hargrove, S. Nieto, T. Zhang, et al., Chem. Rev. 111 (2011) 6603-6782. DOI:10.1021/cr100242s |
| [10] |
J.W. Lichtman, J.A. Conchello, Nat. Methods 2 (2005) 910-919. DOI:10.1038/nmeth817 |
| [11] |
T.B. Ren, W. Xu, W. Zhang, et al., J. Am. Chem. Soc. 140 (2018) 7716-7722. DOI:10.1021/jacs.8b04404 |
| [12] |
X. Chen, X. Tian, I. Shin, et al., Chem. Soc. Rev. 40 (2011) 4783-4804. DOI:10.1039/c1cs15037e |
| [13] |
D. Cheng, J. Peng, Y. Lv, et al., J. Am. Chem. Soc. 141 (2019) 6352-6361. DOI:10.1021/jacs.9b01374 |
| [14] |
H.W. Liu, L. Chen, C. Xu, et al., Chem. Soc. Rev. 47 (2018) 7140-7180. DOI:10.1039/C7CS00862G |
| [15] |
L.Y. Niu, Y.Z. Chen, H.R. Zheng, et al., Chem. Soc. Rev. 44 (2015) 6143-6160. DOI:10.1039/C5CS00152H |
| [16] |
S.R. Cuenca, V. Pellegrinelli, M. Campbell, et al., Prog. Lipid Res. 66 (2017) 14-29. DOI:10.1016/j.plipres.2017.01.002 |
| [17] |
J. Zheng, R. Yang, M. Shi, et al., Chem. Soc. Rev. 44 (2015) 3036-3055. DOI:10.1039/C5CS00020C |
| [18] |
Y. Zhou, Z. Xu, J. Yoon, Chem. Soc. Rev. 40 (2011) 2222-2235. DOI:10.1039/c0cs00169d |
| [19] |
Y. Wu, J. Wen, H. Li, et al., Chin. Chem. Lett. 28 (2017) 1916-1924. DOI:10.1016/j.cclet.2017.09.032 |
| [20] |
A.B. Smiraglia, M.S. Rana, C.E. Foley, et al., Nat. Methods 14 (2017) 1003-1009. DOI:10.1038/nmeth.4404 |
| [21] |
J.H. Kim, J.H. Ahn, P.W. Barone, et al., Angew. Chem. Int. Ed. 49 (2010) 1456-1459. DOI:10.1002/anie.200906251 |
| [22] |
R. Tao, Y. Zhao, H. Chu, et al., Nat. Methods 14 (2017) 720-728. DOI:10.1038/nmeth.4306 |
| [23] |
B.D. Kubena, H. Luecke, H. Rosenberg, et al., J. Biol. Chem. 261 (1986) 7995-7996. |
| [24] |
H. Luecke, F.A. Quiocho, Nature 347 (1990) 402-406. DOI:10.1038/347402a0 |
| [25] |
E. Kimura, M. Kodama, T. Yatsunami, J. Am. Chem. Soc. 104 (1982) 3182-3187. DOI:10.1021/ja00375a042 |
| [26] |
H.J. Schneider, T. Blatter, B. Palm, et al., J. Am. Chem. Soc. 114 (1992) 7704-7708. DOI:10.1021/ja00046a016 |
| [27] |
J.R. Hess, N.A. Greenberg, Nutr. Clin. Pract. 27 (2012) 281-294. DOI:10.1177/0884533611434933 |
| [28] |
F.B. Rudolph, J. Nutr. 124 (1994) 124S-127S. DOI:10.1093/jn/124.suppl_1.124S |
| [29] |
G. Li, V.B. Segu, M.E. Rabaglia, et al., Endocrinology 139 (1998) 3752-3762. DOI:10.1210/endo.139.9.6207 |
| [30] |
B. Alberts, A. Johnson, J. Lewis, et al., Molecular Biology of the Cell, 4th. ed., Garland Science, New York, 2002.
|
| [31] |
K.A. McDonough, A. Rodriguez, Nat. Rev. Microbiol. 10 (2011) 27-38. |
| [32] |
P.B. Dennis, A. Jaeschke, M. Saitoh, et al., Science 294 (2001) 1102-1105. DOI:10.1126/science.1063518 |
| [33] |
A.J. Moro, P.J. Cywinski, K.R. Susanne, et al., Chem. Commun. (Camb.) 46 (2010) 1085-1087. DOI:10.1039/B919661G |
| [34] |
O. Akio, T. Ippei, K. Takahiro, et al., J. Am. Chem. Soc. 130 (2008) 12095-12101. DOI:10.1021/ja803262w |
| [35] |
K. Yasutaka, K. Takahiro, O. Akio, et al., J. Am. Chem. Soc. 132 (2010) 13290-13299. DOI:10.1021/ja103615z |
| [36] |
Y. Kurishita, T. Kohira, A. Ojida, et al., J. Am. Chem. Soc. 134 (2012) 18779-18789. DOI:10.1021/ja308754g |
| [37] |
S. Marbumrung, K. Wongravee, V. Ruangpornvisuti, et al., Sens. Actuators B Chem. 171- 172 (2012) 969-975. |
| [38] |
Q.C. Xu, H.J. Lv, Z.Q. Lv, et al., RSC Adv. 4 (2014) 47788-47792. DOI:10.1039/C4RA07923J |
| [39] |
L. Yan, Z. Ye, C. Peng, et al., Tetrahedron 68 (2012) 2725-2727. DOI:10.1016/j.tet.2012.01.028 |
| [40] |
R.A. Sreenivasa, K. Dokyoung, N. Hyoseok, et al., Chem. Commun. (Camb.) 48 (2012) 3206-3208. DOI:10.1039/c2cc17629g |
| [41] |
S.J. Butler, Chemistry 20 (2015) 15768-15774. |
| [42] |
C. Patra, A.K. Bhanja, A. Mahapatra, et al., RSC Adv. 6 (2016) 6505-76513. |
| [43] |
L. Xiao, S. Sun, Z. Pei, et al., Biosens. Bioelectron. 65 (2015) 166-170. DOI:10.1016/j.bios.2014.10.038 |
| [44] |
Y.G. Gao, Q. Tang, Y.D. Shi, et al., Talanta 152 (2016) 438-446. DOI:10.1016/j.talanta.2016.02.040 |
| [45] |
X. Li, X. Guo, L. Cao, et al., Angew. Chem. Int. Ed. 53 (2014) 7809-7813. DOI:10.1002/anie.201403918 |
| [46] |
M. Bojtár, J. Kozma, Z. Szakács, et al., Sens. Actuators B:Chem. 248 (2017) 305-310. DOI:10.1016/j.snb.2017.03.163 |
| [47] |
D. Maity, M. Li, M. Ehlers, et al., Chem. Commun. (Camb.) 53 (2017) 208-211. DOI:10.1039/C6CC08386B |
| [48] |
T. Noguchi, T. Shiraki, A. Dawn, et al., Chem. Commun. (Camb.) 48 (2012) 8090-8092. DOI:10.1039/c2cc33262k |
| [49] |
G. Jiang, W. Zhu, Q. Chen, et al., Analyst (2017) 4388-4392. |
| [50] |
T. Deng, J.H. Chen, Y. Hui, et al., Sens. Actuators B:Chem. 209 (2015) 735-743. DOI:10.1016/j.snb.2014.12.008 |
| [51] |
S.L. Moodie, J.B. Mitchell, J.M. Thornton, J. Mol. Biol. 263 (1996) 486-500. DOI:10.1006/jmbi.1996.0591 |
| [52] |
E.A. Kataev, T.A. Shumilova, B. Fiedler, et al., J. Org. Chem. 81 (2016) 6505-6514. DOI:10.1021/acs.joc.6b01130 |
| [53] |
C.Y. Li, C.X. Zou, Y.F. Li, et al., Anal. Chim. Acta 795 (2013) 69-74. DOI:10.1016/j.aca.2013.07.065 |
| [54] |
J.L. Tang, C.Y. Li, Y.F. Li, et al., Chem. Commun. (Camb.) 50 (2014) 15411-15414. DOI:10.1039/C4CC08044K |
| [55] |
Y. Fang, W. Shi, Y. Hu, et al., Chem. Commun. (Camb.) 54 (2018) 5454-5457. DOI:10.1039/C8CC02209G |
| [56] |
K.Y. Tan, C.Y. Li, Y.F. Li, et al., Anal. Chem. 89 (2017) 1749-1756. DOI:10.1021/acs.analchem.6b04020 |
| [57] |
Y.W. Jun, T. Wang, S. Hwang, et al., Angew. Chem. Int. Ed. 130 (2018) 10299-10304. DOI:10.1002/ange.201804743 |
| [58] |
Y. Liu, D. Lee, W. Di, et al., Sens. Actuators B:Chem. 265 (2018) 429-434. DOI:10.1016/j.snb.2018.03.081 |
| [59] |
K.M.K. Swamy, S. Eom, Y. Liu, et al., Sens. Actuators B:Chem. 281 (2019) 350-358. DOI:10.1016/j.snb.2018.10.135 |
| [60] |
P. Srivastava, S.S. Razi, R. Ali, et al., Biosens. Bioelectron. 69 (2015) 179-185. DOI:10.1016/j.bios.2015.02.028 |
| [61] |
L. Wang, L. Yuan, X. Zeng, et al., Angew. Chem. Int. Ed. 128 (2016) 1805-1808. DOI:10.1002/ange.201510003 |
| [62] |
J. Qiao, C. Chen, D. Shangguan, et al., Anal. Chem. 90 (2018) 12553-12558. DOI:10.1021/acs.analchem.8b02496 |
| [63] |
Z. Xu, G. Zeng, Y. Liu, et al., Dye. Pigment. 163 (2019) 559-563. DOI:10.1016/j.dyepig.2018.12.035 |
| [64] |
H. Ma, M. Yang, C. Zhang, et al., J. Mater. Chem. B 5 (2017) 8525-8531. DOI:10.1039/C7TB02399E |
| [65] |
A. Nisar, S. Bahareh, I.S. Yoon, et al., Chem. Commun. (Camb.) 48 (2012) 2662-2664. DOI:10.1039/c2cc17145g |
| [66] |
A. Nisar, S. Bahareh, I.S. Yoon, et al., Org. Biomol. Chem. 11 (2013) 6407-6413. DOI:10.1039/c3ob41470a |
| [67] |
A. Nisar, S. Bahareh, G. Inacrist, et al., Org. Lett. 13 (2011) 5476-5479. DOI:10.1021/ol202183t |
| [68] |
M. Yousuf, N. Ahmed, B. Shirinfar, et al., Org. Lett. 16 (2014) 2150-2153. DOI:10.1021/ol500613y |
| [69] |
Z. Xu, N.R. Song, J.H. Moon, et al., Org. Biomol. Chem. 9 (2011) 8340-8345. DOI:10.1039/c1ob06344h |
| [70] |
A.M. Agafontsev, T.A. Shumilova, T. Rüffer, et al., Chem.-Eur. J. 25 (2019) 3541-3549. DOI:10.1002/chem.201806130 |
| [71] |
N. Wu, J. Lan, L. Yan, et al., Chem. Commun. (Camb.) 50 (2014) 4438-4441. DOI:10.1039/C4CC00752B |
| [72] |
P.P. Neelakandan, P.C. Nandajan, S. Baby, et al., Org. Biomol. Chem. 9 (2011) 1021-1029. DOI:10.1039/c0ob00673d |
| [73] |
S.C. Karunakaran, A.K. Paul, D. Ramaiah, RSC Adv. 4 (2014) 30644-30647. DOI:10.1039/C4RA04672B |
| [74] |
L. Shi, P. Hu, Y. Ren, et al., Chem. Commun. (Camb.) 49 (2013) 11704-11706. DOI:10.1039/c3cc47437b |
| [75] |
P. Hu, S. Yang, G. Feng, Org. Biomol. Chem. 12 (2014) 3701-3706. DOI:10.1039/C4OB00184B |
| [76] |
F. Huang, G. Hao, F. Wu, et al., Analyst 140 (2015) 5873-5876. DOI:10.1039/C5AN01291K |
| [77] |
M. Zhang, W.J. Ma, C.T. He, et al., Inorg. Chem. 52 (2013) 4873-4879. DOI:10.1021/ic302435g |
| [78] |
D. Wang, X. Zhang, C. He, et al., Org. Biomol. Chem. 8 (2010) 2923-2925. DOI:10.1039/c004148c |
| [79] |
B. Roy, T. Noguchi, D. Yoshihara, et al., Org. Biomol. Chem. 12 (2014) 561-565. DOI:10.1039/C3OB41586D |
| [80] |
B. Ma, F. Zeng, X. Li, et al., Chem. Commun. (Camb.) 48 (2012) 6007-6009. DOI:10.1039/c2cc32064a |
| [81] |
J. Jung, M.S. Eom, K. Baek, et al., Tetrahedron Lett. 56 (2015) 5847-5850. DOI:10.1016/j.tetlet.2015.08.067 |
| [82] |
L.M. Tumir, M. Grabar, S. Tomi c, et al., Tetrahedron 66 (2010) 2501-2513. DOI:10.1016/j.tet.2010.01.063 |
| [83] |
K. Ghosh, S.S. Ali, A.R. Sarkar, et al., Org. Biomol. Chem. 11 (2013) 5666-5672. DOI:10.1039/c3ob40833g |
| [84] |
S. Das, H.S. Sarkar, M.R. Uddin, et al., Chem. Commun. (Camb.) 53 (2017) 7600-7603. DOI:10.1039/C7CC02935G |
| [85] |
J. Kong, T. Liu, Y. Bao, et al., Talanta 117 (2013) 412-418. DOI:10.1016/j.talanta.2013.09.020 |
| [86] |
S.H. Lu, R. Phang, J.M. Fang, Org. Lett. 18 (2016) 1724-1727. DOI:10.1021/acs.orglett.6b00311 |
| [87] |
L.M. Zhang, L.E. Guo, X.M. Li, et al., Tetrahedron Lett. 55 (2014) 6131-6136. DOI:10.1016/j.tetlet.2014.09.074 |
| [88] |
L.F. Yousif, K.M. Stewart, K.L. Horton, et al., ChemBioChem 10 (2009) 2081-2088. DOI:10.1002/cbic.200900017 |
| [89] |
S. Zhang, T. Wu, J. Fan, et al., Org. Biomol. Chem. 11 (2013) 555-558. DOI:10.1039/C2OB26911B |
| [90] |
W. Ying, Front. Biosci. 11 (2006) 3129-3148. DOI:10.2741/2038 |
| [91] |
V. Massey, Biochem. Soc. Trans. 28 (2000) 283-296. |
| [92] |
E.J. Wood, Biochem. Mol. Biol. Educ. 32 (2010) 421-423. |
| [93] |
G.J.D. Birkmayer, W. Birkmayer, Acta Neurol. Scand. 80 (1989) 183-187. DOI:10.1111/j.1600-0404.1989.tb01800.x |
| [94] |
K. Hirokazu, S. Yutaka, O. Kotaro, et al., Angew. Chem. Int. Ed. 53 (2014) 3993-3995. DOI:10.1002/anie.201311192 |
| [95] |
M.A. Fomin, R.I. Dmitriev, J. Jenkins, et al., ACS Sens. 1 (2016) 702-709. DOI:10.1021/acssensors.5b00315 |
| [96] |
Y. Zhao, K. Wei, F. Kong, et al., Anal. Chem. 91 (2019) 1368-1374. DOI:10.1021/acs.analchem.8b03563 |
| [97] |
M. Santra, S. Sarkar, Y.W. Jun, et al., Tetrahedron Lett. 59 (2018) 3210-3213. DOI:10.1016/j.tetlet.2018.07.031 |
| [98] |
S.O. Jung, J.Y. Ahn, S. Kim, et al., Tetrahedron Lett. 51 (2010) 3775-3778. DOI:10.1016/j.tetlet.2010.05.044 |
| [99] |
R. Zadmard, P.A. Moghaddam, S. Darvishi, et al., Tetrahedron 73 (2017) 604-607. DOI:10.1016/j.tet.2016.12.053 |
| [100] |
L. Wang, J. Zhang, B. Kim, et al., J. Am. Chem. Soc. 138 (2016) 10394-10397. DOI:10.1021/jacs.6b05810 |
| [101] |
A.N. Mayeno, K.J. Hamann, G.J. Gleich, J. Leukoc. Biol. Suppl. 51 (1992) 172-175. DOI:10.1002/jlb.51.2.172 |
| [102] |
C.D.C. Chichi, F. Feillet, J.L. Guéant, et al., Am. J. Clin. Nutr. 71 (2000) 978-986. DOI:10.1093/ajcn/71.4.978 |
| [103] |
R.H. Woo, C.S. Jung, Y.S. Ho, et al., J. Am. Chem. Soc. 131 (2009) 10107-10112. DOI:10.1021/ja9018012 |
| [104] |
O. Jinrok, H.J. In, Org. Lett. 15 (2013) 1210-1213. DOI:10.1021/ol303403x |
| [105] |
D.N. Lee, E. Kim, J.H. Lee, et al., Chem. Commun. (Camb.) 52 (2016) 13487-13490. DOI:10.1039/C6CC07483A |
| [106] |
D.N. Lee, S. Bae, K. Han, et al., Chem.-Eur. J. 23 (2017) 16078-16084. DOI:10.1002/chem.201703466 |
| [107] |
L. Ulicna, D. Paprckova, V. Faberova, et al., Histochem. Cell Biol. 150 (2018) 245-253. DOI:10.1007/s00418-018-1690-9 |
| [108] |
B. Fadeel, D. Xue, Crit. Rev. Biochem. Mol. Biol. 44 (2009) 264-277. DOI:10.1080/10409230903193307 |
| [109] |
G. van Meer, D.R. Voelker, G.W. Feigenson, Nat. Rev. Mol. Cell Biol. 9 (2008) 112-124. DOI:10.1038/nrm2330 |
| [110] |
D. Hishikawa, T. Hashidate, T. Shimizu, et al., J. Lipid Res. 55 (2014) 799-807. DOI:10.1194/jlr.R046094 |
| [111] |
A.P. Demchenko, Exp. Oncol. 34 (2012) 263-268. |
| [112] |
S.H. Lee, X.W. Meng, K.S. Flatten, et al., Cell Death Differ. 20 (2013) 64-76. DOI:10.1038/cdd.2012.93 |
| [113] |
E.T. Baronas, J.W. Lee, C. Alden, et al., Toxicol. Appl. Pharmacol. 218 (2007) 72-78. DOI:10.1016/j.taap.2006.10.015 |
| [114] |
B.R. Lentz, Prog. Lipid Res. 42 (2003) 423-438. DOI:10.1016/S0163-7827(03)00025-0 |
| [115] |
J.G. Kay, G. Sergio, Sensors 11 (2011) 1744-1755. |
| [116] |
D.R. Rice, K.J. Clear, B.D. Smith, Chem. Commun. (Camb.) 52 (2016) 8787-8801. DOI:10.1039/C6CC03669D |
| [117] |
K.J. Clear, K.M. Harmatys, D.R. Rice, et al., Bioconjug. Chem. 27 (2016) 363-375. DOI:10.1021/acs.bioconjchem.5b00447 |
| [118] |
A.J. Plaunt, K.M. Harmatys, W.R. Wolter, et al., Bioconjug. Chem. 25 (2014) 724-737. DOI:10.1021/bc500003x |
| [119] |
B.A. Smith, K.M. Harmatys, S. Xiao, et al., Mol. Pharm. 10 (2013) 3296-3303. DOI:10.1021/mp300720k |
| [120] |
D.R. Rice, H. Gan, B.D. Smith, Photochem. Photobiol. Sci. 14 (2015) 1271-1281. DOI:10.1039/C5PP00100E |
| [121] |
B.A. Smith, S. Xiao, W. Wolter, et al., Apoptosis 16 (2011) 722-731. DOI:10.1007/s10495-011-0601-5 |
| [122] |
T.S. Jarvis, F.M. Roland, K.M. Dubiak, et al., J. Mater. Chem. B 6 (2018) 4963-4971. DOI:10.1039/C8TB01495G |
| [123] |
A.C.S. Leung, E. Zhao, R.T.K. Kwok, et al., J. Mater. Chem. B 4 (2016) 5510-5514. DOI:10.1039/C6TB01734G |
| [124] |
Q. Hu, M. Gao, G. Feng, et al., ACS Appl. Mater. Inter. 7 (2015) 4875-4882. DOI:10.1021/am508838z |
| [125] |
V.E. Zwicker, B.L. Oliveira, J.H. Yeo, et al., Angew. Chem. Int. Ed. 58 (2019) 3087-3091. DOI:10.1002/anie.201812489 |
| [126] |
W. Xu, Z. Zeng, J.H. Jiang, et al., Angew. Chem. Int. Ed. 55 (2016) 13658-13699. DOI:10.1002/anie.201510721 |
| [127] |
S.H. Alamudi, R. Satapathy, J. Kim, et al., Nat. Commun. 7 (2016) 11964. DOI:10.1038/ncomms11964 |
| [128] |
S. Letschert, A. Gohler, C. Franke, et al., Angew. Chem. Int. Ed. 53 (2014) 10921-10924. DOI:10.1002/anie.201406045 |