 2019, Vol. 30
2019, Vol. 30 



b School of Life Sciences, Kyungpook National University, Daegu 41566, Republic of Korea
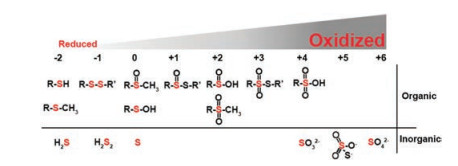
Sulfur is an important element in biological system. It has different oxidation states ranging from -2 to +6, such as hydrogen sulfide, elemental sulfur, thiol, thiolether, sulfinic acid, sulfonic acid, sulfate (Fig. 1) [1]. Thiols are important species among them and include protein thiols and low molecular weight thiols (LMWTs), such as cysteine (Cys), homocysteine (Hcy), and glutathione (GSH). It is well known that thiols (i.e., sulfhydryl) can react with reactive oxygen species (ROS), reactive nitrogen species (RNS), and reactive sulfur species (RSS) owing to its high reactivity under physiological environment [2]. The fluctuation of LMWTs is closely related with cancer [3], cardiovascular diseases [4], diabetes [5], and neurodegenerative diseases [6]. Protein thiols are more abundant than LMWTs within a cell (~70% of the total cellular thiols) and also play an important role in diverse physiological and pathological progresses [7-9].

|
Download:
|
| Fig. 1. Oxidation states of sulfur ranging from -2 to +6. | |
{kind=link}
Protein vicinal dithiols (PVD) refer to a pair of spatially adjacent Cys residues in a protein. This motif is widely present in many proteins and essential for maintaining proteins' structure. In addition, the conversion between vicinal dithiols and disulfides plays vital roles inmanybiological processes, such asmaintenanceof intracellular redox homeostasis and regulation of protein synthesis as well as posttranslational modifications [10, 11]. Accumulating studies have disclosed a linkage between the aberrant dithiols/ disulfides conversion and many diseases [12-16]. The important functions of PVD have stimulated researchers to develop various methods for selective detection of PVD [17-20].
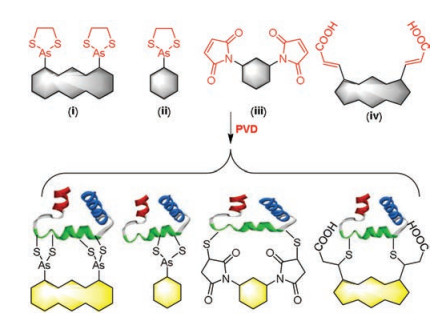
Fluorescent imaging has emerged as a powerful technique in various fields including chemical biological, clinical diagnosis and drug discovery [21, 22]. Especially, small molecule fluorescent probe has been a popular tool in determining multiple biological events due to their outstanding sensitivity, good biocompatibility and high temporal-spatial resolution [23-25]. In recent years, a large number of fluorescent probes have been applied to sense and analyze peptides or proteins that contain vicinal dithiol moiety. In this review, we summarized these fluorescent probes. According to their acceptor units, these fluorescent probes were classified into four types (Fig. 2): ⅰ) biarsenical probes, which comprise two space-closed dithiarsolanes as acceptor units; ⅱ) monoarsenical probes, which contain trivalent arsenic compound (one dithiarsolane or one p-aminophenylarsenoxide (PAO) moiety) as a specific acceptor; ⅲ) dimaleimide probes, where two maleimide groups are attached directly to the fluorophores; ⅳ) diacrylate probes, where two acrylic acid groups or acrylic ester groups are used as acceptor units. Through analyzing and evaluating these fluorescent probes in this review, we expect to further understand the structural factors that determine the properties of probes, and shed light in designing novel probes.

|
Download:
|
| Fig. 2. Overview of fluorescent probes for protein vicinal dithiols. | |
{kind=link}
2. Biarsenical probes
In order to study the structures and functions of a protein of interest (POI), various methods have been applied, such as gel electrophoresis [26, 27], X-Ray diffraction [28, 29], molecular simulation [30, 31], and nuclear magnetic resonance [32]. However, these methods are time-consuming and complex. In the past decades, fluorescent protein (FP) fused to the POI was used for noninvasive imaging in live cells and organisms. Nevertheless, the large size of FP might disturb the functions of POI [33]. For overcoming this disadvantage, a series of biarsenical probes specifically targeting the tetracysteine (TC) tag has been developed.
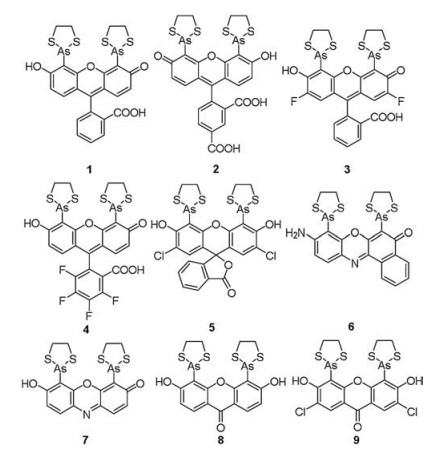
2.1. FlAsH and its analoguesIn 1998, Tsien et al. [34] designed and synthesized the first biarsenical probe (1) comprised fluorescein and two dithiarsolanes (Fig. 3). In addition, they designed a peptide domain with four cysteine residues at the i, i+1, i+4, and i+5 positions of an α-helix, which could bind to dithiarsolane moiety of probe 1. This binding elicits bright fluorescence (λex = 508 nm, λem = 528 nm, Φ = 0.49) and could be completely reversed by small molecule dithiols such as 2, 3-dimercaptopropanol (BAL) or 1, 2-ethanedithiol (EDT). This probe realized the fluorescent labeling of a TC motif appended to a green fluorescent protein (GFP) mutant in live cells.

|
Download:
|
| Fig. 3. Structures of probes 1–9. | |
{kind=link}
The non-specific fluorescence is increased when biarsenical probes bind to membranes and hydrophobic pockets of proteins, which results in high background fluorescence. In order to overcome this problem, Mayer et al. [35] developed a biarsenical multi-use affinity probe (2, Fig. 3). The obvious fluorescence signal of 2 towards peptide with TC moiety was observed (λex = 520 nm, λem = 534 nm) and the fluorescence intensity of complex reached a maximum around pH 7. Titration of probe 2 towards albumin had a little effect on fluorescence intensity, which suggested the probe had an enhanced signal-to-noise ratio.
The limited photostability and pH sensitivity of biarsenical probes based on fluorescein derivatives restricted the further application of fluorescent imaging technology. In 2006, JaresErijman et al. [36] designed the difluorinated derivatives (3) and tetrafluorinated derivatives (4) (Fig. 3). In the presence of a model sequence (FLNCCPGCCMEP, P12) with TC moiety, probe 3 exhibited higher absorbance (εmax = 65500 L mol-1 cm-1) and larger Stokes shift (λex = 500 nm, λem = 522 nm) compared to probe 1. The fluorescence intensity (λex = 528 nm, λem = 544 nm) of probe 4 and P12 was similar to that of probe 1. Moreover, phosphate buffers with pH ranging from 5.6 to 7.8 had a little effect on the absorption of probes 3 and 4. The complexes of the fluorinated dyes exhibited a corresponding brighter emission at lower pH. In addition, probes 3 and 4 could form an excellent FRET pair because they have a favorable spectral overlap. These properties are expected to facilitate studies of proteins in live cells.
In 2010, Otlewski et al. [37] also reported a new dichlorinated biarsenical probe (5, Fig. 3) based on 2, 7-dichlorofluorescein. Probe 5 caused an obvious fluorescence increment (λex = 520 nm, λem = 539 nm) after reacted with target peptide (AcFLNCCPGCCMEP-NH2, L1) with the TC moiety.
In 2001, Umezawa et al. [38] developed the first environmentsensitive fluorescent probe (6, Fig. 3) for specifically binding to TC moiety, which was an analogue of nile red with a biarsenical moiety. In the presence of TC moiety, a time-dependent fluorescence increase could be observed (λex = 550 nm, λem = 604 nm). This probe was used to label the recombinant calmodulin containing the TC motif (CaM-helix) inside live HEK 293 cells, and image the conformational change of CaM-helix upon [Ca2+] increases. Owing to different fluorescent intensity of this probe in various solvents, probe 6 had high potential for detecting the change of protein microenvironment and studying signal transduction in live cells.
Based on the above research, Tsien et al. [39] synthesized various analogues of FlAsH, including a resorufin analogue (7) and blue-fluorescing biarsenical probes (8 and 9) (Fig. 3). They designed a series of TC sequences and found that the binding was tightest for biarsenical probes in the presence of the sequence Cys-Cys-Pro-Gly-Cys-Cys, which suggested that the preferred conformation of TC sequence was a hairpin rather than α-helix. In addition, they observed that the most likely pairing for the two arsenics of FlAsH analogues was i, i+1 and i+4, i+5 positions in TC peptides. probe 7 was non-fluorescent in the free state, but rapidly displayed a red fluorescent signal (λex = 593 nm, λem = 608 nm) when bound with a TC-containing peptide. The binding of 8 and 9 with TC tag also displayed an obvious fluorescence signal enhancement (> 20-fold) (λex = 380 nm, λem = 430 nm). Furthermore, probe 9 may be used as a fluorescence resonance energy transfer (FRET) donor for GFP or yellow fluorescent protein (YFP) and for multicolor labeling with other biarsenical probe or autofluorescent proteins.
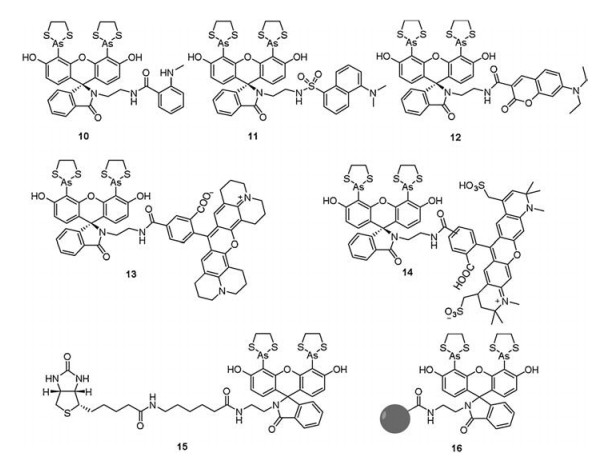
2.2. Fluorescein spirolactam biarsenical probesFrom the structures of various probes 1 and 7 derivatives, both fluorophores and the distance of arsenic moieties have limited the range of fluorescent labeling reagents that can be targeted to TC tags. In 2007, Miller et al. [40] adopted a modular approach that they linked a biarsenical fluorescein spirolactam (SplAsH) to several fluorophores for extending the scope of compatible dyes (Fig. 4), such as N-methylanthranilate (MANT; 10), dansyl (11), diethylamino-coumarin (DEAC; 12), X-rhodamine (ROX; 13), and Alexa-594 (14). In addition, probes 13 and 14 exhibited remarkably improved photostability compared with 1 and 7.

|
Download:
|
| Fig. 4. Structures of probes 10–16. | |
{kind=link}
In 2011, Ying et al. [41] synthesized a SplAsH derivative (15, Fig. 4) as a biarsenical affinity ligand to purify TC tagged proteins. The maximum absorbance of probe 15 was 242 nm with ε242 = 26000 L mol-1 cm-1 and there was no inherent fluorescence whether or not it was bound to the TC motif. Probe 15 exhibited improved photostability and more than 1000-fold affinity. Furthermore, SplAsH was conjugated with Affigel-10 to get the new non-fluorescent affinity matrix 16 for affinity purification of a TC-tagged and His6-tagged GFP (Fig. 4).
2.3. Carboxyfluorescein biarsenical probesFor developing novel genetically-encoded affinity tags to purify proteins, Cooke et al. [42] designed a novel affinity matrix based on the biarsenical fluorescein dye (17, Fig. 5A), which specifically recognized short α-helical peptides containing the TC sequence and produced fluorescence signal. The primary amine on the probe 17 reacted with N-hydroxysuccinamide (NHS) functionalized agarose beads, which gave a stable covalent linkage. Through purifying kinesin with TC peptide by biarsenical affinity chromatography, they found that this method could yield purer protein than the conventional nickel affinity chromatography of polyhistidine tagged kinesins.

|
Download:
|
| Fig. 5. (A) Structures of probes 17–30. (B) QD encapsulation and coupling to CrAsH. Copied with permission [61]. Copyright 2008, Journal of American Chemical Society. | |
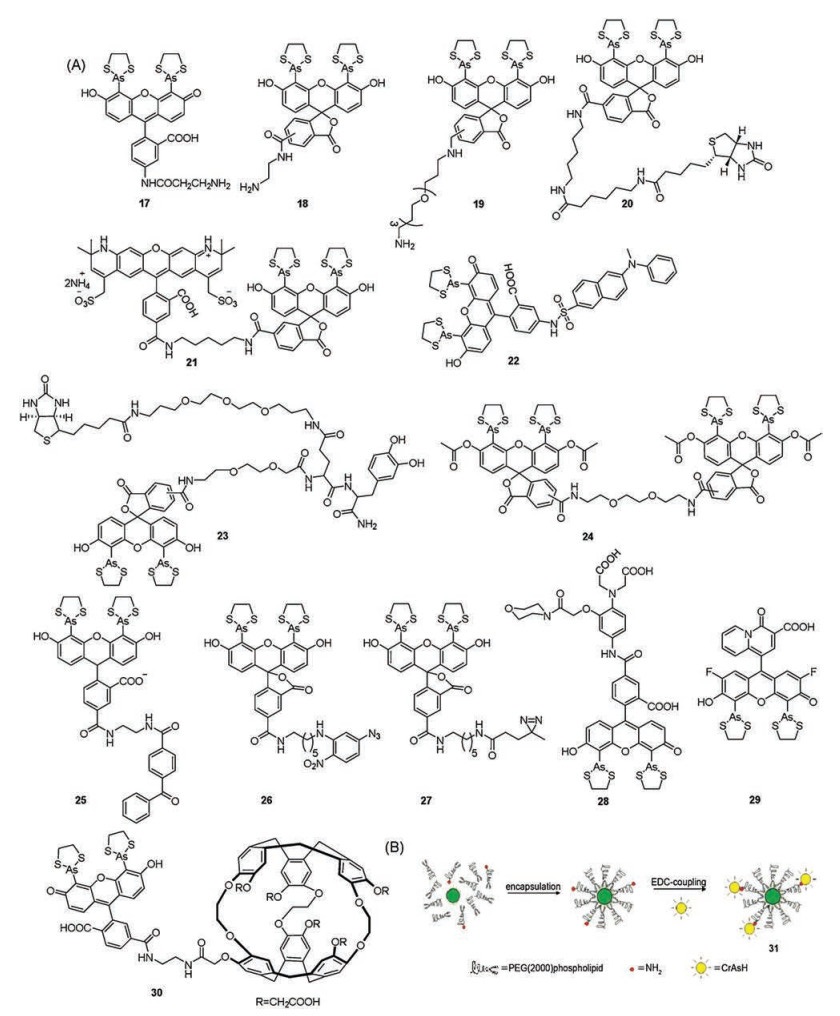{kind=link}
In 2005, Mayer et al. [43] also developed a biarsenical probe (18) immobilized on NHS-activated glass beads as a stationary phase for affinity purification of TC-tagged proteins (Fig. 5A). They isolated the Shewanella oneidensis RNA polymerase core enzyme with a TC-tagged RNA polymerase A as bait protein using this affinity probe resin. This one-step and non-denaturing procedure will be a useful method for protein binding partner identification with high yields enough to permit structural measurements of the intact complex.
In 2014, Triola et al. [44] developed a novel one-step method for protein immobilization onto biarsenical probe-modified surfaces. They designed and synthesized probe 19, and then glass slides activated with NHS were coated with the amine group of probe 19 to generate the biarsenical modified slides (Fig. 5A). Once the peptides or proteins containing TC sequence were spotted onto the modified slides, direct detection of immobilized peptides or proteins was successfully performed by determining the fluorescence intensity upon formation of the complex. Moreover, a TCcontaining prenyl-binding protein (PDEδ) was efficiently immobilized from cell lysates without prior purification, which suggested that this method expanded the repertoire of immobilization methods.
In 2008, Baron et al. [45] developed a novel technique, instant with dithiothreitol (DTT), EDT, and low temperature (IDEAL)- labeling, for rapid and specific labeling of TC-tagged cell surface proteins. Using this method, they observed that TC-tagged prion protein (PrP) converted from the cell surface normal isoform (PrPsen) to the disease-associated isoform (PrPres) in live cells. Furthermore, they designed and synthesized two new probes (20 and 21, Fig. 5A), which consisted of conjugates to Alexa Fluor dye and biotin, respectively, and possessed improved fluorescence properties, versatility of detection, and ease of affinity purification compared with probe 1. The two probes successfully labeled TCtagged amyloid precursor protein (APP) and imaged PrP endocytosis and ultrastructural localization. Therefore, IDEAL-labeling technique will be an important method for extracellular proteins and beyond microscopic imaging.
In order to develop a more sensitive protein conformation indicator, Umezawa et al. [46] designed and synthesized a novel probe (22, Fig. 5A) that conjugated mansyl chloride to the benzoic acid moiety of FlAsH. The extended benzoic acid moiety of this probe promoted the flexible interaction of the fluorophore with protein surfaces, which made the probe more sensitive to the protein conformational change of calmodulin attached to the TC moiety (λex = 480 nm, λem = 533 nm). Molecular calculation of probe 22 suggested that the sensitive fluorescence response towards TC moiety was due to the photoinduced electron transfer (PET) process.
The detection of protein–protein interactions is important for understanding various biological processes including the function of multi-protein complexes and the regulation of signal transduction networks [47, 48]. In 2007, Kodadek et al. [49] developed probe 23 that contained tethered biotin and dihydroxyphenylalanine (DOPA) residues to study protein-protein interactions (Fig. 5A). They linked the "bait" protein with a TC-containing peptide (FRP) for specifically binding with biarsenical moiety. Once the "bait" protein reacted with partner proteins, the DOPA residues was transformed into a reactive orthoquinone that could crosslink with nearby nucleophilic amino acids in the presence of sodium periodate. After crosslinking, the biarsenical-FRP complex could be dissociated by boiling in an excess of dithiol, which resulted in transferring the biotin to nearby partner proteins. This novel label transfer system will be a useful procedure for the characterization of protein-protein interactions in their native environments.
In 2011, Schultz et al. [50] developed probe 24 as a tool for studying protein–protein interactions (Fig. 5A). It comprised two biarsenical molecules via a flexible linker, and was acetylated to increase solubility during synthesis and to ensure permeability in cell applications. As a novel cross-linker molecule, probe 24 could specifically recognize and crosslink two interacting proteins each containing only a 12 amino acid peptide tag (P12). Furthermore, this binding could produce fluorescent signal and be disconnected by addition of BAL. Through fluorescent imaging and western blotting, they verified that probe 24 could crosslink TC-tagged proteins in vitro and in live cells.
In 2009, Mayer et al. [51] developed a new crosslinker targeted releasable affinity probe (25, Fig. 5A). The biarsenical moiety of probe 25 could selectively label an engineered TC-tagged site on a target protein in a complex mixture and the benzophenone subunit was photocrosslinked to capture binding partners. When the crosslinked complex reacted with excess dithiol, the crosslinker was released and transferred to the binding partner for efficient mass spectrometric identification of the crosslinking site. Owing to high photo-crosslinking yields and well cell-permeability of probe 25, it was capable to crosslink the TC-tagged α subunit of RNA polymerase in live S. oneidensis MR-1 bacterial cells and phospholamban in C2C12 myocyte cells. Therefore, the simple method will facilitate the high-throughput identification of protein complexes in different organisms.
The photoaffinity probe 25 usually needed to be irradiated for long times, which promoted non-specific labeling. In 2016, Georg et al. [52] designed and synthesized two new photo-crosslinking biarsenical probes (26 and 27, Fig. 5A). Probe 26 carried a phenylazide as the photo-crosslinker group, which produced a highly reactive nitrene species after irradiated by UV light. Probe 27 contained a diazirin moiety at C5 of the benzoate moiety, which yielded a reactive carbene species and long-lived electrophilic diazo compounds after irradiated by long wave UV light. All of reactive species could react with nucleophiles. In order to provide enhanced flexibility to react with binding partners, they designed the longer linker at C5 in both probes than probe 25. Furthermore, probes 26 and 27 could be used to label recombinant doublesex and Mab3-related transcription factor (DMRT1) containing the TC motif and produce fluorescent signal in nuclear. Therefore, the two photoaffinity probes will be expected to be useful tools for crosslinking DNA or proteins associated with the tagged protein and allowing identification of DNA regulatory sequences and protein partners.
Intracellular calcium ion (Ca2+) regulates various proteins functions and cellular activities [53, 54], so it is necessary to develop novel tools for detecting Ca2+. In 2007, Tsien et al. [55] developed a biarsenical Ca2+ indicator (28) to study the concentration variation of Ca2+ surrounding genetically targeted proteins (Fig. 5A). When the biarsenical moiety of probe 28 bound to TC peptide, there was a 4-fold fluorescent enhancement. O-Aminophenol-N, N, O-triacetic acid could be used as the Ca2+ binding site. After the complex bound to Ca2+, the fluorescence enhanced a further 10-fold (λem = 535 nm) with a Kd = 100 μmol/L for Ca2+. Furthermore, probe 28 could be used to label gap junctions and report the fluctuation of [Ca2+] after injury in HeLa cells expressing TC-tagged connexin 43. In addition, this probe showed a fluorescent response towards TC-tagged α1C L-type calcium channels in human embryonic kidney (HEK293) cells.
Magnesium ion (Mg2+) plays important roles in many physiological processes in live cells, but its mobilization and underlying mechanisms are still unknown [56]. For studying the dynamics of intracellular Mg2+ around particular proteins, target organelles, and Mg2+ transporters or channels, Oka et al. [57] designed and synthesized a novel fluorescent Mg2+ probe (29), which composed of Mg2+ report moiety (KMG-104) and a fluorescent labeling probe bound specifically to TC tag (Fig. 5A). probe 29 was non-fluorescent in the free state. However, there was a 10-fold fluorescent enhancement when 29 bound to both the TCtagged peptide and Mg2+ (λex = 490 nm, λem = 540 nm), and the probe was highly selective towards Mg2+ (Kd/Mg = 1.7 mmol/L, Kd/Ca ≫ 100 mmol/L). Through labeling TC-tagged mKeima localized to the mitochondrial intermembrane space with probe 29, they detected the diffusion of Mg2+ from the mitochondria to the cytoplasm induced by carbonyl cyanide p-(trifluoromethoxy) phenylhydrazone (FCCP) in HeLa cells. Therefore, this probe was useful for localized imaging and mechanism study of Mg2+.
Biarsenical probes have been used as effective tools for detecting and imaging TC-tagged protein. However, such probes show poor tissue permeability. In 2015, Rousseau et al. [58] designed and synthesized a doubly responsive probe (30) for providing in-depth anatomical and physiological information of target proteins on tissues (Fig. 5A). probe 30 comprised a probe 2 moiety for specifically binding with TC tag and a hexacarboxylic acid cryptophane-A derivative (MM-2) as a 129Xe NMR-based biosensor. MM-2 possesses biological ligand and can reversibly bind to xenon in solution [59]. When probe 30 bound to the TCtagged peptide, it exhibited a strong fluorescent signal (λex = 480 nm, λem = 535 nm) as well as a 129Xe NMR signal highly specific for caged xenon.
Although various biarsenical affinity probes have been developed, classical photobleaching of the organic dye still limits the widespread application of these probes. Semiconductor nanocrystals, also known as quantum dots (QDs), have exhibited high quantum yield, large molar extinction coefficient, tunable fluorescence emission and photostability [60]. In 2008, Dubertret et al. [61] designed and synthesized a new nanohybrid (31, Fig. 5B). This nanohybrid exhibited better photostability and higher selectivety with TC tags (λex = 350 nm, λem = 520 nm) compared to probe 2.
2.4. Cyanine dye biarsenical probesThe interatomic distance between the two arsenic moieties of various biarsenical derivatives is conserved (~6 Å), whichmakes the selective labeling of multiple proteins with different reports complicated. In 2007, Mayer et al. [62] synthesized a new red emission small molecule biarsenical multiuse affinity probe (32, Fig. 6). This probe comprised Cy3, a member of the cyanine dye family, as fluorophore and possessed large interatomic distance between the two arsenics in probe 32 (~14.5 Å). Furthermore, a complementary high-affinity binding sequence (CCKAEAACC, Cy3TAG) was coupled with probe 32, which accomplished within 15 s. In addition, this probe had absorbance and fluorescence emission maxima peak at 560 and 568 nm, and the emission spectrum of probe 1 overlapped with the absorption spectrum of probe 32, which could act as an acceptorof FRET to allow ratiometric measurementsofprotein association.Therefore, probe 32 couldbe a colored probe for directing different small binding moiety.

|
Download:
|
| Fig. 6. Structures of probes 32–34. | |
{kind=link}
In order to facilitate cellular delivery and quantitative imaging applications of probe 32, Squier et al. [63] designed and synthesized probe 33 with both an ethanedithiol cap and uncharged methoxy ester functionalized N-alkyl chains (Fig. 6). Carboxylic acid methyl ester was introduced to modify probe hydrophobicity. When the probe was delivered into cells, endogenous esterases were expected to hydrolyze the methyoxy esters for enhancing solubility of probe. This progress was likely to minimize hydrophobic interactions with non-target proteins. When probe 33 bound to RNA polymerase (RpoA*) with a TC sequence (Cy3TAG), there was a red shift in both absorbance and emission spectra (λex = 565 nm, λem = 579 nm) and higher fluorescence quantum yield (Φ = 0.21) compared to probe 32. probe 33 exhibited improved cell permeability apparent even in bacterial cells and enhanced photostability, which provided a general method to monitor protein localization and dynamics.
For further studying the structural arrangement and dynamics of subunits within macromolecular protein complexes, the superresolution imaging technology has been developed [64-66]. In 2012, Squier et al. [67] designed and synthesized a new superresolution imaging probe (34, Fig. 6). It was prepared by coupling unsymmetrical probe 32 and Cy5, which were separated by a linker (< 21 Å) through the indolium nitrogens and designed to produce FRET effect [68]. The close distance of Cy3 and Cy5 permitted rapid photo-switching, which could realize super-resolution visualization of tagged proteins. Once binding with the target proteins (RpoA*), there was a 20-fold fluorescent enhancement in the emission of Cy5 by the increase in the ratio of the emission of Cy5 (acceptor) relative to Cy3 (donor).
3. Monoarsenical probesBiarsencial probes could specifically recognize the TC sequence and be a tool for studying the structures and functions of POI with TC-tag. Some reviews about the application of biarsencial probes in protein research have been published [69, 70]. There are two pairs of vicinal dithiols moiety in TC sequence. Each dithiarsolane moiety in biarsenical probe binds with a pair of vicinal dithiols moiety. So the biarsenical probe could be thought of as a tool for detecting the sequence with two pairs of vicinal dithiols structure. However, TC sequence is less content in living system and it needs to be fused into the target protein, which is a complex process. Moreover, PVD could be thought of as a part of TC sequence and plays a critical role in biological system. Therefore, monoarsenical probes that contained one dithiarsolane or one PAO moiety as specific acceptor for sensing PVD have obtained remarkable progress.
3.1. Conventional monoarsenical probesIn 2011, Xu et al. [71] developed a highly selective and cellpermeable fluorescent probe (35) for rapidly detecting and imaging of vicinal dithiols in proteins (Fig. 7). This probe comprised naphthalimide as a fluorophore and one dithiarsolane as a specific acceptor (λex = 460 nm, λem = 538 nm). probe 35 was used to label the reduced Thioredoxin (rTrx) containing one pair of vicinal dithiols and produced fluorescent signal by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE), so they speculated that the exchange of EDT in the dithiarsolane of probe 35 with PVD induced covalent-bond formation between vicinal dithiols moiety and the probe. Through imaging probe 35-labeled Chang liver cells, they observed that the PVD was mainly localized in the mitochondria. After the cells were pretreated with different redox reagents such as DTT or diamide, cellular PVD showed different fluorescent response through probe 35-labeling. So this probe will be useful for the proteomic-scale identification of PVD under physiological and pathological conditions.

|
Download:
|
| Fig. 7. Structures of probes 35–42. | |
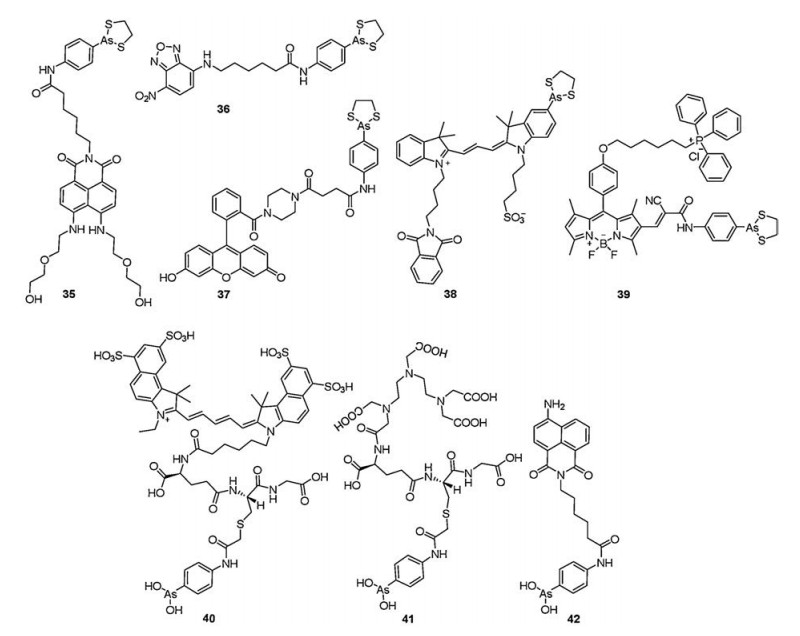{kind=link}
In order to offer new tools for the potential identification and visualization of vicinal dithiols existing in different regions of proteins in live cells, Zhu et al. [72] developed a conjugated approach to expand detectable tags (dithiarsolane) to nitrobenzoxadiazole (NBD), fluorescein, naphthalimide, and biotin for the synthesis of a series of probes. Especially, probes 36 (λex = 495 nm, λem = 553 nm) and 37 (λex = 500 nm, λem = 523 nm) may make in situ imaging of vicinal thiols of different protein regions (Fig. 7). This versatile chemical strategy affords a convenient method for the in situ imaging of PVD and the proteomics study of various disease-related proteins with a vicinal dithiol motif.
Squier et al. [73] had synthesized a thiol-reactive affinity probe (38, Fig. 7) for permitting quantitative measurements of changes in the redox state of cellular vicinal dithiols. It was consisted of photostable and red emission environment-insensitive cyanine and dithiarsolane moiety. probe 38 was used to label an engineered protein (i.e., C4-CaM) with proximal introduced cysteines (i.e., EEQIAE11 to CCQICC11) and reduced bovine serum albumin (rBSA) using SDS-PAGE. Obvious red fluorescent signal could be observed after the tagged engineered protein expressed in E. coli (i.e., C4- CaM) reacted with probe 38, which verified the cell-permeability of probe 38 and the ability of detecting redox-active vicinal dithiols in live bacterial cells. Furthermore, when increasing the levels of oxygen and light in the photosynthetic microbe Synechococcus sp. PCC7002, weak fluorescence intensity was observed following the separation of cellular lysates by SDS-PAGE. This result suggested that the reduced state of PVD greatly decreased under oxidative stress.
In 2018, Kim et al. [74] developed a smart fluorescent probe 39 containing one dithiarsolane as a specific receptor and 4- hydroxyphenyl-tetramethyl boron-dipyrromethene (BODIPY) as a fluorophore (λex = 513 nm, λem = 534 nm, Fig. 7). In addition, probe 39 was conjugated to triphenylphosphonium group via an alkyl spacer targeted the mitochondria. The probe could be used for specifically labeling and imaging PVD on mitochondrial membrane. After imaging the oligomeric Aβ1-42-pretreated HeLa cells with this probe, the decreased fluorescence intensity was observed, which resulted from the increasement of oxidized forms of mitochondrial PVD. So probe 39 will be a super-resolution imaging reagent of PVD on mitochondrial membrane.
Cell death is an important characteristic of biological phenomenon and plays a significant role under normal physiological condition and in disease [75, 76]. In 2011, Hogg et al. [77] produced a peptide trivalent organoarsenical compound, 4-(N-(S-glutathionylacetyl)amino)phenylarsonous acid (GSAO), and they linked GSAO with different reporter groups (i.e., fluorophores, biotin, and radioisotopes) through the γ-glutamyl amine for selectively labeling dying and dead cells. The most abundant molecular chaperone of the eukaryotic cytoplasm, the 90-kDa heat shock protein (Hsp90) containing a highly conserved Cys-Cys motif in the C-terminal domain, could selectively bind to GSAO. When the integrity of the plasma membrane was destroyed, the abundance of Hsp90 in the cytosol made a mass of GSAO conjugates rapidly accumulate in apoptotic cells and allowed for superior detection and resolution of cell death. Especially, probes 40 and 41 could image cell death in cultured cells and in murine tumors ex vivo and in situ (Fig. 7). In a word, GSAO conjugates could be used as a celldeath imaging reagents based on their chemical stability and selectivity to Hsp90.
Based on the above design strategy, Fang et al. [78] synthesized probe 42 linking PAO with naphthalimide (Fig. 7). probe 42 was used to label the selenocysteine-deficient thioredoxin reductase (U498C TrxR) containing vicinal dithiols moiety and the complex exhibited fluorescent signal by SDS-PAGE.
3.2. Ratiometric monoarsenical probesMost monoarsenical probes contained strong fluorescence background resulted from unreacted or nonspecifically bound probes. In contrast, ratiometric fluorescent probes can eliminate most nonspecific fluorescent intensity by self-calibration of two emission peaks [79, 80]. In 2014, Xu et al. [81] developed the first ratiometric fluorescent probe (43) based on FRET mechanism for specifically detecting PVD (Fig. 8). probe 43 contained a coumarin and a naphthalimide for fluorescence signal transduction. In the absence of PVD, the fluorescent signal of the probe presented the emission of naphthalimide (λem = 540 nm) when excited at 405 nm. After selective exchange of 2, 3-dimercaptopropanol moiety of probe 43 with vicinal dithiols moiety, PVD was labeled by coumarin moiety and the complex presented the emission of coumarin (λem = 470 nm). probe 43 was used to image endogenous PVD in live MCF-7 cells and trace the changes of PVD under different cellular redox states. In addition, the mitochondria distribution of endogenous PVD was clearly visualized in live cells through colocalization experiments and ratiometric images of this probe. Therefore, this ratiometric fluorescence probe 43 offered a noninvasive tool for trapping and quantifying dynamic changes of endogenous PVD in live cells.

|
Download:
|
| Fig. 8. Reaction mechanism of probe 43 for detecting PVD. Reproduced with permission [81]. Copyright 2014, Journal of American Chemical Society. | |
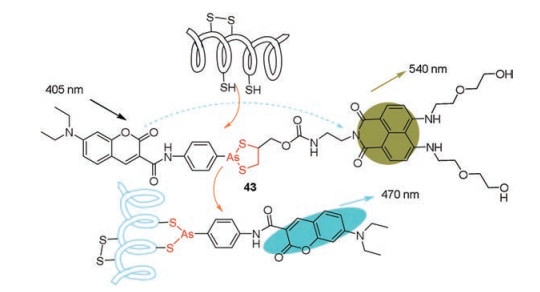{kind=link}
3.3. Environment-sensitive monoarsenical probes
Although ratiometric fluorescence probe 43 could overcome high fluorescent background, the probe still exhibited long response time (>60 min), short wavelength emission and moderate fluorescence intensity changes (~6 fold). In addition, the environmentsensitive fluorescent probe as a novel probe design strategy for detecting biomacromolecule has been developed [82-86]. In 2015, Yang et al. [87] firstly designed and synthesized a red emission and environment-sensitive fluorescent probe (44) for the rapid detection of PVD (Fig. 9). The probe contained 2-(4-dimethylaminophenyl)-4-(2-carboxyph-enyl)-7-diethylamino-1-benzopyrylium as the environment-sensitive fluorescence reporter with red emission and dithiarsolane as the specific ligand. In the absence of PVD, probe 44 showed negligible fluorescent intensity in buffer. However, once the trivalent arsenical of the probe specifically reacted with vicinal dithiols moiety, the fluorophore entered into the hydrophobic protein domain, which caused the strong red fluorescent signal (λex = 621 nm, λem = 651 nm). The fluorescence enhancement of the probe could be explained by reducing the charge transfer between the fluorophore and the polar media and restricting the intramolecular rotations via aggregation simultaneously. Therefore, probe 44 was applied for imaging PVD in live SMMC-7721 cells without tedious washing steps.

|
Download:
|
| Fig. 9. Structures of environment-sensitive monoarsenical probes (probes 44–48). | |
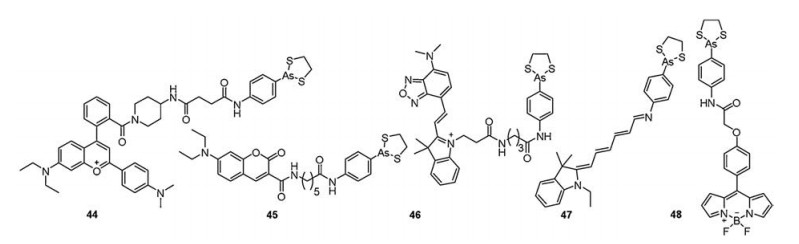{kind=link}
In 2016, Yang et al. [88] developed probe 45 as a new fluorescent probe for sensing PVD based on the modulation of intramolecular charge transfer (ICT) to twisted intramolecular charge transfer (TICT) conversion process (Fig. 9). 7-Diethylamincoumarin, which was a solvatochromic dye, as the fluorescence reporter and dithiarsolane as the specific ligand were introduced into probe 45. The probe existed both as ICT and TICT excited states upon photoexcitation, and it featured a broad emission peak at 550 nm. Since the hydrophobic protein domain with the low polarity environment, the ICT-TICT conversion was restricted. Hence, the emission peak at 468 nm increased dramatically upon addition of PVD, concomitant with a decrease of TICT emission peak at 550 nm. The emission ratio (I468/I550) of the probe essentially reached a maximum after 20 min. Furthermore, this probe was used for ratiometric fluorescent imaging of PVD in live L929 cells and probe 45-labeled proteins were mainly distributed in the mitochondria of live SMMC-7721 cells by fluorescence colocalization experiments.
In 2017, Jiang et al. [89] developed a novel fluorescent probe (46) using an engineered benzoxadiazole fluorophore selectively targeting for mitochondria, and exhibited deep red emission (630 nm), high environmental sensitivity, instantaneous responsibility (within 30 s) toward PVD (Fig. 9).
In 2018, Yang et al. [90] reported a far-red fluorescence turn-on probe (47) contained a merocyanine Schiff base and dithiarsolane for sensing PVD based on a pKa shift mechanism (Fig. 9). Under physiological environment (pH 7.4), probe 47 was predominantly present as the Schiff base (SB) form (pKa = 4.8), which was absent of a positive change, and the probe had a relatively short absorbance wavelength and was essentially non-fluorescent. Upon selectively binding to PVD, probe 47 entered into the binding domain of PVD, which had a considerable effect on the electronic charge distribution of the probe. It was present as protonated Schiff base (PSB) form (pKa = 7.1) in the low polarity environment of protein. The pKa shift, which led to red shift and a significant increasement in fluorescence intensity (λem = 657 nm), achieved the purpose of sensing PVD. After imaging the endogenous PVD in SMMC-7721 cells with probe 47, the cells displayed a bright red fluorescence signal.
Autophagy is a general cellular progress in eukaryotes, which could catabolize misfolded proteins, recycle components, or produce energy. Autophagy of the endoplasmic reticulum (ER), termed reticulophagy, which contributes to the protein quality control process and the accumulation of unfolded or misfolded proteins in the ER and leads to ER stress [91]. In 2019, Kim et al. [92] developed an environment-sensitive, dual-functional molecular rotor (48) for studying the change of viscosity in ER stress (Fig. 9). It contained a BODIPY-based fluorescent rotor as a microviscosity sensor that could determine local viscosity and a dithiarsolane moiety for selectively binding to PVD (λex = 488 nm, λem = ~515 nm). With the predominantly localization in the ER, probe 48 could initiate ER stress and induce reticulophagy by causing the accumulation of misfolded or unfolded proteins after 1 h incubation. Fluorescence lifetime imaging (FLIM) of the probe showed the viscosity increased in the ER during the fusion of the ER with the lysosome in the course of reticulophagy. Therefore, probe 48 would be a tool for specifically measuring viscosity changes associated with reticulophagy.
4. Dimaleimide probesBiarsenical probes and monoarsenical probes have been developed based on the binding of trivalent arsenical compounds to the paired thiols of proteins. However, these compounds possess cytotoxicity. It is known that the maleimide unit is a welldocumented thiols receptor and there are many fluorescent sensors that used maleimide as receptor to react with thiols [93, 94]. In addition, these probes containing maleimide moiety exhibit high biocompatibility and low toxicity. So a series of dimaleimide probes has been designed and synthesized for specifically labeling vicinal dithiols moiety in proteins.
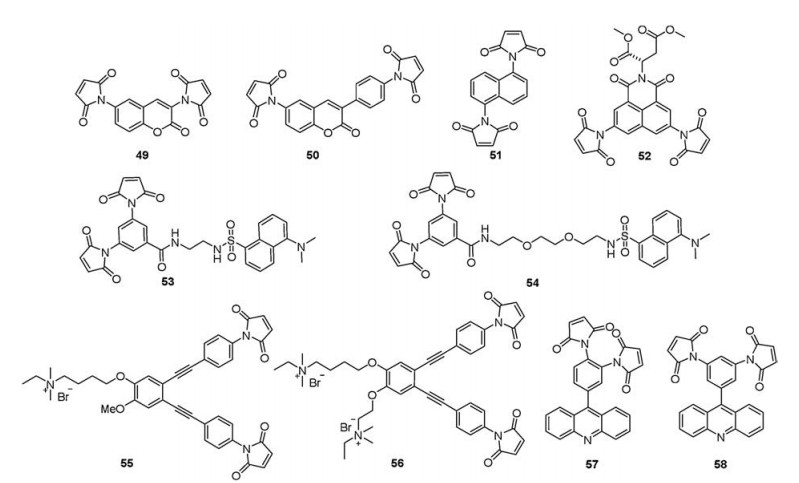
4.1. Maleimide groupsIn 2004, Keillor et al. [95] developed a series of dimaleimide probes (49–52) that two maleimide groups attached directly to fluorophores (Fig. 10). Through studies of kinetic and spectral characterization, they verified that these dimaleimide probes generated fluorescence signal when the probes reacted with thiols and formed the dithiolated adduct. The probe 50 displayed negligible fluorescence signal after reacting with 1 equiv. of mutant Trx (Cys35→Ala, C35A-Trx) by SDS-PAGE. However, obvious fluorescent bands were observed after a recombinant α- helical protein (diCys-Fox) bearing two cysteine residues in positions reacted with probe 50 or 52 (1 equiv.) by SDS-PAGE. These results demonstrated the efficacy of the covalent fluorescent labeling reaction and the feasibility of the application of this method to labeling specific proteins.

|
Download:
|
| Fig. 10. Structures of probes 49–58. | |
{kind=link}
In 2007, Keillor et al. [96] developed a new convergent synthetic route that allowed fluorophores to be attached to a dimaleimide moiety via a linker in a modular fashion. They designed and synthesized dimaleimide and dansyl fluorophore derivatives (53 and 54) via a linker to explore the mechanism of dimaleimide quenching (Fig. 10). Furthermore, they verified that a PET quench mechanism played a key role in quenching the dimaleimide dansyl system. This quenching mechanism will provide a theoretical guide for designing fluorescent labeling reagents of proteins.
In 2013, Ma et al. [97] designed and synthesized two novel fluorescent probes (55 and 56, Fig. 10). The two probes comprised the V-shape oligo(o-phenylene-ethynylene) (OPE) backbone as fluorophore and two maleimide groups at the distance of about 1 nm. In addition, quaternary ammonium salts were introduced to provide water solubility. They displayed obvious fluorescence signal towards PVD (λex = 340 nm, λem = ~410 nm). Moreover, probe 56 exhibited higher selectivity towards PVD owing to better solubility.
In 2015, Tang et al. [98] developed two active-site-matched fluorescent probes (57 and 58, Fig. 10) comprised two maleimide groups with different distances. The probes could match two adjacent active cysteine residues in peptides or proteins. Spectroscopic properties of probes to vicinal-dithiols-containing peptides and proteins, such as WCGPCK (W-6), WCGGPCK (W-7), and WCGGGPCK (W-8), rBSA, and rTrx, were measured respectively.Probe 57 displayed shorter response time (1 min) and higher binding sensitivity with the above-mentioned peptides and proteins, especially with W-7 and rTrx (λex = 360 nm, λem = 460 nm), which because the shorter distance between the two maleimide groups in probe 57 provided a better match with the model peptides. Furthermore, probe 57 was successfully used to rapidly and directly detect endogenous PVD in live HepG2 cells.
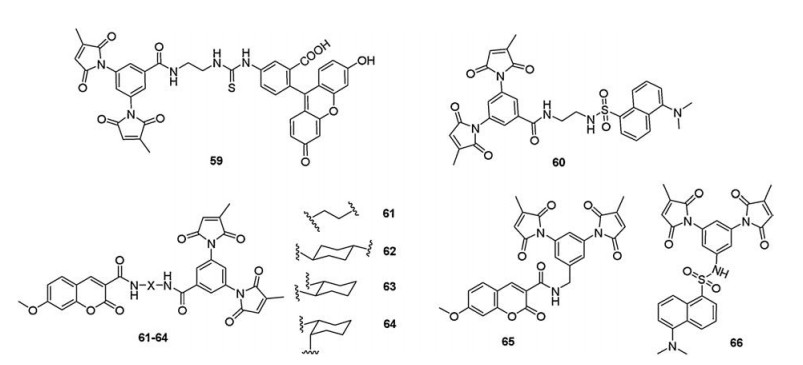
4.2. Methylmaleimide groupsIn 2010, Keillor et al. [99] prepared new di-(3-methylmaleimide) probes (59 and 60, Fig. 11) and designed a novel peptide sequence (dC10α) presenting two adjacent cysteine residues separated by two turns of an α-helix. Furthermore, they constructed the fusion protein (MBP-dC10α) through genetically fusing dC10α sequence to maltase binding protein (MBP) for structural and kinetic characterization. Probe 59 had a high fluorescence enhancement (FE = 5.8) upon reaction with two equivalents of thiol in aqueous media (λex = 494 nm, λem = 517 nm) and showed even greater kinetic selectivity for reaction with the MBP-dC10α (>131-fold) compared to GSH. Moreover, probe 59 was used to selectively label epidermal growth factor receptors (EGFR) bearing the dC10α in HEK 293 cells.

|
Download:
|
| Fig. 11. Structures of probes 59–66. | |
{kind=link}
Previous studies have shown that fluorophore and dimaleimide groups within a certain distance displayed lower fluorescence background via a PET mechanism. These compounds could be used to label specific protein and needed not to be washed out. In order to study the impacts of central spacer moiety on the quenching efficiency, Keillor et al. [100] synthesized a series of coumarinbased dimaleimide probes (61–65, Fig. 11), which contained coumarin as fluorophore and dimaleimide as quenched group with different central spacer moiety. Through exploring the fluorometric characterization of probes (61–65) with two equivalents of 2- mercaptopropionic acid (MPA), probe 65 had the highest fluorescence change (FE = 6.40) (λex = 347 nm, λem = 404 nm). They further carried out the molecular modeling and verified the dimaleimide-to-fluorophore distance was negative correlated with the fluorescence enhancement of probes upon reaction with MPA. Based on the results obtained from the series of coumarin fluorophore, they designed another "spacerless" probe (66, Fig. 11) containing dansyl as an appropriate fluorophore and a dimaleimide aniline moiety directly attached to dansyl without an intermediate spacer moiety. Fluorescence intensity of probe 66 (~530 nm) increased by a factor of more than 300-fold upon reaction with MPA.
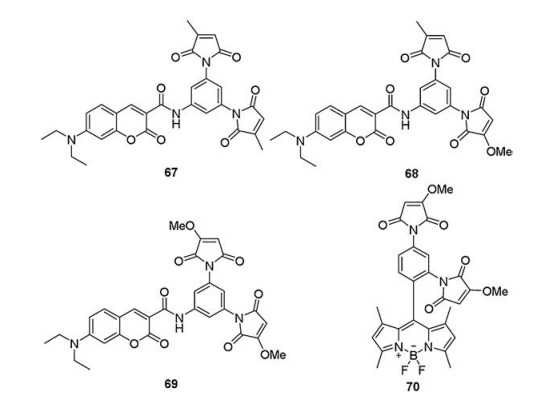
4.3. Methoxymaleimide groupsIn 2014, Keillor et al. [101] developed two novel dimaleimide probes (67 and 68, Fig. 12). probe 68 contained 7-diethylamino coumarin as the fluorophore and two different dimaleimide moieties. It exhibited a better selectivity with MBP-dC10α compared to probe 67, which suggested that the methoxymaleimide moiety has higher reactivity with the dC10α tag. Inspired by these results, they designed the probe 69 bearing two methoxymaleimide moieties, which induced obvious fluorescence signal (λex = 440 nm, λem = 485 nm) after reacted with MBP–dC10α (Fig. 12). HEK 293 T cells transfected with mNeptune–dC10α showed strong cyan fluorescence after incubated with the probe 69 and without any washing steps. The results confirmed that probe 69 was highly selective for dC10α and could be used as a non-wash labeling reagent in live cells.

|
Download:
|
| Fig. 12. Structures of probes 67–70. | |
{kind=link}
Both the dansyl and coumarin groups previously reported fluorophores require excitation with UV or blue light, raising questions regarding the risk of cellular photodamage. In 2018, Keillor et al. [102] designed and synthesized probe 70 with longer emission wavelength (~525 nm) (Fig. 12). It comprised the BODIPY sketelon and two methoxymaleimide groups, at both the ortho position and the para position of the meso-phenyl ring. The probe exhibited a remarkable fluorescence enhancement (FE = ~800) and high rate constant (868 ± 118 L mol-1 min-1) towards the MBPdC10α. probe 70 could be used to selectivelyand robustlylabelMBPdC10α in the complex biological mixture of cell lysate via SDS-PAGE. Furthermore, HeLa cells expressing histone-H2B-dC10α exhibited strong green fluorescence signal after incubated with probe 70.
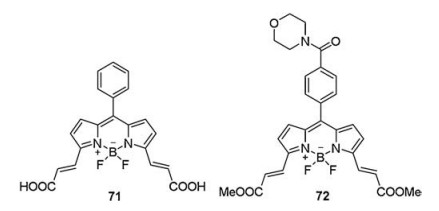
5. Diacrylate probesSite-specific labeling of target proteins with photophysical reporter probes allows numerous in vivo studies of protein functions. Fluorophores possessed dimaleimide groups have been sufficiently developed. Inspired by these dimaleimide probes, Chang et al. [103] designed and synthesized BODIPY-diacrylate probes (71 and 72, Fig. 13) for targeted proteins in live cells. Probe 71 comprised BODIPY-derivative and two acrylic acid groups. The space between the two acrylic groups was similar to the distance separating the two reactive cysteines in the peptide tag. probe 71 exhibited high selectivity towards a model peptide (P1, it had two cysteines at i and i+4 positions in an α-helix and was named as RC tag) and produced obvious fluorescent signal (λex = 480 nm, λem = 530 nm). In addition, they synthesized probe 72 consisted of two acrylic ester groups and morpholine at the para-position of the phenyl ring, which was used to specifically label RC-tagged Cherry and showed an apparent green fluorescence band (λex = 488 nm, λem = 526 nm) by SDS-PAGE. Furthermore, they used probe 72 as a fluorescent labeling reagent to image the RC2 tagged (the dimer of RC tag) recombinant proteins expressed in 293A cells. Dimeric peptide tag RC2 and its partner compound 72 enriched the current protein labeling methods, especially the optical imaging of specific target protein in live cells.

|
Download:
|
| Fig. 13. Structures of probes 71 and 72. | |
{kind=link}
6. Conclusions and outlook
The fluorescent labeling of POI for studying protein structures and functions have aroused increasing interest over the past decade, owing to their potential in monitoring various physiological and pathological activities, and illuminating the molecular mechanisms of different biological events. The most commonly applied methods genetically fused to POI for specific protein fluorescent labeling include the fluorescent protein [33], recognition tag [104, 105], and unnatural amino acid [106]. These methods have significantly facilitated our understandings on biological system and undoubtedly become powerful methods for POI. Among various recognition tags, TC tag or vicinal dithiols moiety in protein exhibits less steric hindrance and better accessibility in living system compared to other recognition tags, such as SNAP-tag [107], Halo-tag [108], or PYP-tag [109], which because the considerable size (ca. 14–30 kDa) of these tags poses a risk for steric perturbation of the POI.
Based on the above, numerous small molecule fluorescent probes for labeling PVD have been developed in recent years. In this review, we have summarized the representative fluorescent probes specifically recognizing and labeling the vicinal dithiol moiety in proteins. These probes are consisted of diverse fluorophores and acceptor moieties, which provide effective tools for detecting vicinal dithiols in living system. They are classified into four types according to different acceptor moieties. Biarsenical probes could specifically bind with TC tag and be widely applied in protein research, which play important roles in biotech applications as switches, linkers and sensing tools. Monoarsenical probes have been developed into the common tools for studying PVD in biological systems. In addition, various fluorescent mechanisms have made the application of monoarsenical probes enriched and expanded. However, the toxicity of organoarsenic probes and the low content of TC sequence in biological system have limited their further application in protein research. Dimaleimide and diacrylate probes exhibit low toxicity and high selectivity towards vicinal dithiols moiety. However, the imaging of depth is still limited because the short emission wavelength of dimaleimide and diacrylate probes. Without doubt, these fluorescent probes in this review have pushed forward the development of protein research and detection of vicinal dithiols. By analyzing and evaluating photoproperties and structures of these probes, it is observed that the same acceptor moiety accommodated to different fluorophores will exhibit enhanced photoproperties, which can further understand the relationship of structures and functions and provide the purposeful design strategy of novel probes for detecting vicinal dithiols.
Although existing small molecule fluorescent probes of PVD have made a great effect on protein study, several obstacles still exist for the practical applications, the current challenges as well as the potential future directions are listed as below: (1) In this review, these fluorescent probes are classified into four types according to their acceptor units. Following the disulfide rebridging reagents developing [110, 111], the novel fluorescent probes based on other electrophile (such as bissulfones, dibromopyridazinediones, alkynes) as acceptor site may be designed for vicinal dithiols. Through developing novel acceptor units, the application scope of the probe will be extended and the physiological activities and pathological changes of biological system will be better interpreted and understood. (2) The conversion between vicinal dithiols and disulfides participates in many bioactivities and the aberrant dithiols/disulfides conversion is closely related to diverse major diseases. Research models are important for shedding light onto the molecular mechanisms of different biological phenomena. In this review, we could find some research models, such as reticulophagy model, prion protein endocytosis model, cell-death model. However, other models of different important biological activities should also been focused. For example, 6-hydroxydopamine (6-OHDA)-treated PC12 cells, a well-established Parkinsonism model [112]. It is well known that the loss of protein thiols was closely associated with the Parkinsonism disease [113], however, there is no direct experimental data that the level of PVD decreases in Parkinsonism model. So detecting the state of PVD in disease models with novel fluorescent probes is urgently needed in the future. (3) The optical properties of fluorescent probe have an effect on their purpose and function. In this review, the large aromatic structures in most probes may cause fluorophore aggregation through π-π stacking, which can not only hamper the cellular permeability and subcellular localization, but also quench the fluorescence of probes. In addition, these probes exhibit one photon performance, which is limited to cell imaging and is not useful for deep-tissue imaging due to the shallow depth of tissue penetration (less than 100 mm) of the short excitation light [114]. Therefore, the development of novel probes with aggregation induced emission (AIE) or two photon performance is highly desired in the future work.
In summary, great efforts have been made to develop fluorescent probe of PVD during the past decade, and these probes have accelerated protein research. Although there are still many obstacles and challenges remained to be conquered, it is hoped that more novel fluorescent probes with higher performance will be emerged in the future.
AcknowledgmentsThe financial supports from the National Natural Science Foundation of China (Nos. 21572093 & 21778028) and the 111 Project are greatly acknowledged.
| [1] |
N. Lau, M.D. Pluth, Curr. Opin. Chem. Biol. 49 (2019) 1-8. |
| [2] |
C.E. Paulsen, K.S. Carroll, Chem. Rev. 113 (2013) 4633-4679. DOI:10.1021/cr300163e |
| [3] |
M.P. Gamcsik, M.S. Kasibhatla, S.D. Teeter, O.M. Colvin, Biomarkers 17 (2012) 671-691. DOI:10.3109/1354750X.2012.715672 |
| [4] |
Y. Ozkan, E. Ozkan, B. Simsek, Int. J. Cardiol. 82 (2002) 269-277. DOI:10.1016/S0167-5273(02)00010-4 |
| [5] |
I.C. West, Diabetic Med. 17 (2000) 171-180. DOI:10.1046/j.1464-5491.2000.00259.x |
| [6] |
B.D. Paul, J.I. Sbodio, S.H. Snyder, Trends Pharmacol. Sci. 39 (2018) 513-524. DOI:10.1016/j.tips.2018.02.007 |
| [7] |
R.E. Hansen, D. Roth, J.R. Winther, Proc. Natl. Acad. Sci. U. S. A. 106 (2009) 422-427. DOI:10.1073/pnas.0812149106 |
| [8] |
J.D. Short, K. Downs, S. Tavakoli, R. Asmis, Antioxid. Redox Signal. 25 (2016) 816-835. DOI:10.1089/ars.2016.6697 |
| [9] |
T.K. Lin, G. Hughes, A. Muratovska, et al., J. Biol. Chem. 277 (2002) 17048-17056. DOI:10.1074/jbc.M110797200 |
| [10] |
K.M. Cook, P.J. Hogg, Antioxid. Redox Signal. 18 (2013) 1987-2015. DOI:10.1089/ars.2012.4807 |
| [11] |
C.R. Borges, D.F. Lake, Antioxid. Redox Signal. 21 (2014) 392-395. DOI:10.1089/ars.2014.5946 |
| [12] |
X.W. Zhang, X.J. Yan, Z.R. Zhou, et al., Science 328 (2010) 240-243. DOI:10.1126/science.1183424 |
| [13] |
N. Houstis, E.D. Rosen, E.S. Lander, Nature 440 (2006) 944-948. DOI:10.1038/nature04634 |
| [14] |
J. Sugatani, M.E. Steinhelper, K. Saito, et al., J. Biol. Chem. 262 (1987) 16995-17001. |
| [15] |
C. Gitler, M. Mogyoros, E. Kalef, Methods Enzymol. 233 (1994) 403-415. DOI:10.1016/S0076-6879(94)33047-6 |
| [16] |
A. Nott, P.M. Watson, J.D. Robinson, et al., Nature 455 (2008) 411-415. DOI:10.1038/nature07238 |
| [17] |
R. Requejo, E.T. Chouchani, A.M. James, et al., Arch. Biochem. Biophys. 504 (2010) 228-235. DOI:10.1016/j.abb.2010.09.003 |
| [18] |
O. Rudyk, P. Eaton, Redox Biol. 2 (2014) 803-813. DOI:10.1016/j.redox.2014.06.005 |
| [19] |
R.D. Hoffman, M.D. Lane, J. Biol. Chem. 267 (1992) 14005-14011. |
| [20] |
R. Moaddel, A. Sharma, T. Huseni, et al., Bioconjugate Chem. 10 (1999) 629-637. DOI:10.1021/bc9801575 |
| [21] |
J.W. Lichtman, J.A. Conchello, Nat. Methods 2 (2005) 910-919. DOI:10.1038/nmeth817 |
| [22] |
J.A. Cotruvo, A.T. Aron, et al., Chem. Soc. Rev. 44 (2015) 4400-4414. DOI:10.1039/C4CS00346B |
| [23] |
X. Li, X. Gao, W. Shi, H. Ma, Chem. Rev. 114 (2014) 590-659. DOI:10.1021/cr300508p |
| [24] |
B. Li, Z. He, H. Zhou, et al., Chin. Chem. Lett. 28 (2017) 1929-1934. DOI:10.1016/j.cclet.2017.08.055 |
| [25] |
L. Yu, Y. Qiao, L. Miao, et al., Chin. Chem. Lett. 29 (2018) 1545-1559. DOI:10.1016/j.cclet.2018.09.005 |
| [26] |
Ünlü M., M.E. Morgan, J.S. Minden, Electrophoresis 18 (1997) 2071-2077. DOI:10.1002/elps.1150181133 |
| [27] |
D.W. Cleveland, S.G. Fischer, M.W. Kirschner, U.K. Laemmli, J. Biol. Chem. 252 (1977) 1102-1106. |
| [28] |
H. Frauenfelder, G.A. Petsko, D. Tsernoglou, Nature 280 (1979) 558-563. DOI:10.1038/280558a0 |
| [29] |
G.A. Petsko, D. Ringe, Annu. Rev. Biophys. Bioeng. 13 (1984) 331-371. DOI:10.1146/annurev.bb.13.060184.001555 |
| [30] |
A.D. MacKerell, D. Bashford, M. Bellott, et al., J. Phys. Chem. B 102 (1998) 3586-3616. DOI:10.1021/jp973084f |
| [31] |
A. Neumaier, SIAM Rev 39 (1997) 407-460. DOI:10.1137/S0036144594278060 |
| [32] |
D.S. Wishart, B.D. Sykes, F.M. Richards, J. Mol. Biol. 222 (1991) 311-333. DOI:10.1016/0022-2836(91)90214-Q |
| [33] |
K.M. Dean, A.E. Palmer, Nat. Chem. Biol. 10 (2014) 512-523. DOI:10.1038/nchembio.1556 |
| [34] |
B.A. Griffin, S.R. Adams, R.Y. Tsien, Science 281 (1998) 269-272. DOI:10.1126/science.281.5374.269 |
| [35] |
H. Cao, B. Chen, T.C. Squier, M.U. Mayer, Chem. Commun. (2006) 2601-2603. |
| [36] |
C.C. Spagnuolo, R.J. Vermeij, Jares-Erijman E.A., J. Am. Chem. Soc. 128 (2006) 12040-12041. DOI:10.1021/ja063212q |
| [37] |
A. Pomorski, J. Otlewski, A. Krezel, Chembiochem 11 (2010) 1214-1218. DOI:10.1002/cbic.200900768 |
| [38] |
J. Nakanishi, T. Nakajima, M. Sato, et al., Anal. Chem. 73 (2001) 2920-2928. DOI:10.1021/ac001528p |
| [39] |
S.R. Adams, R.E. Campbell, L.A. Gross, et al., J. Am. Chem. Soc. 124 (2002) 6063-6076. DOI:10.1021/ja017687n |
| [40] |
A.K. Bhunia, S.C. Miller, Chembiochem 8 (2007) 1642-1645. DOI:10.1002/cbic.200700192 |
| [41] |
L.Q. Ying, B.P. Branchaud, Bioconjugate Chem. 22 (2011) 987-992. DOI:10.1021/bc200038t |
| [42] |
K.S. Thorn, N. Naber, M. Matuska, et al., Protein Sci. 9 (2000) 213-217. |
| [43] |
Uljana Mayer M., L. Shi, T.C. Squier, Mol. BioSyst. 1 (2005) 53-56. DOI:10.1039/b500950b |
| [44] |
Schulte-Zweckel J., F. Rosi, D. Sreenu, et al., Chem. Commun. 50 (2014) 12761-12764. DOI:10.1039/C4CC04120H |
| [45] |
Y. Taguchi, Z.D. Shi, B. Ruddy, et al., Mol. Biol. Cell 20 (2009) 233-244. DOI:10.1091/mbc.e08-06-0635 |
| [46] |
J. Nakanishi, M. Maeda, Y. Umezawa, Anal. Sci. 20 (2004) 273-278. DOI:10.2116/analsci.20.273 |
| [47] |
D.M. Spencer, T.J. Wandless, S.L. Schreiber, G.R. Crabtree, Science 262 (1993) 1019-1024. DOI:10.1126/science.7694365 |
| [48] |
E.J. Licitra, J.O. Liu, Proc. Natl. Acad. Sci. U. S. A. 93 (1996) 12817-12821. DOI:10.1073/pnas.93.23.12817 |
| [49] |
B. Liu, C.T. Archer, L. Burdine, et al., J. Am. Chem. Soc. 129 (2007) 12348-12349. DOI:10.1021/ja072904r |
| [50] |
A. Rutkowska, C.H. Haering, C. Schultz, Angew. Chem. Int. Ed. 50 (2011) 12655-12658. DOI:10.1002/anie.201106404 |
| [51] |
P. Yan, T. Wang, G.J. Newton, et al., Chembiochem 10 (2009) 1507-1518. DOI:10.1002/cbic.200900029 |
| [52] |
S.S. Syeda, D. Rice, D.J. Hook, et al., Arch. Pharm. 349 (2016) 233-241. DOI:10.1002/ardp.201500440 |
| [53] |
G.J. Augustine, F. Santamaria, K. Tanaka, Neuron 40 (2003) 331-346. DOI:10.1016/S0896-6273(03)00639-1 |
| [54] |
H. Parnas, L. Segel, J. Dudel, I. Parnas, Trends Neurosci. 23 (2000) 60-68. DOI:10.1016/S0166-2236(99)01498-8 |
| [55] |
O. Tour, S.R. Adams, R.A. Kerr, et al., Nat. Chem. Biol. 3 (2007) 423-431. DOI:10.1038/nchembio.2007.4 |
| [56] |
N. Grimsley, T. Hohn, J.W. Davies, B. Hohn, Nature 325 (1987) 177-179. DOI:10.1038/325177a0 |
| [57] |
T. Fujii, Y. Shindo, K. Hotta, et al., J. Am. Chem. Soc. 136 (2014) 2374-2381. DOI:10.1021/ja410031n |
| [58] |
N. Kotera, E. Dubost, G. Milanole, et al., Chem. Commun. 51 (2015) 11482-11484. DOI:10.1039/C5CC04721H |
| [59] |
M.M. Spence, S.M. Rubin, I.E. Dimitrov, et al., Proc. Natl. Acad. Sci. U. S. A. 98 (2001) 10654-10657. DOI:10.1073/pnas.191368398 |
| [60] |
X. Michalet, F. Pinaud, L. Bentolila, et al., Science 307 (2005) 538-544. DOI:10.1126/science.1104274 |
| [61] |
E. Genin, O. Carion, B. Mahler, et al., J. Am. Chem. Soc. 130 (2008) 8596-8597. DOI:10.1021/ja802987q |
| [62] |
H. Cao, Y. Xiong, T. Wang, et al., J. Am. Chem. Soc. 129 (2007) 8672-8673. DOI:10.1021/ja070003c |
| [63] |
N. Fu, Y. Xiong, T.C. Squier, Bioconjugate Chem. 24 (2013) 251-259. DOI:10.1021/bc300619m |
| [64] |
G.H. Patterson, Lippincott-Schwartz J., Science 297 (2002) 1873-1877. DOI:10.1126/science.1074952 |
| [65] |
B. Huang, W. Wang, M. Bates, X. Zhuang, Science 319 (2008) 810-813. DOI:10.1126/science.1153529 |
| [66] |
R.M. Dickson, A.B. Cubitt, R.Y. Tsien, et al., Nature 388 (1997) 355-358. DOI:10.1038/41048 |
| [67] |
N. Fu, Y. Xiong, T.C. Squier, J. Am. Chem. Soc. 134 (2012) 18530-18533. DOI:10.1021/ja308503x |
| [68] |
C.R. Sabanayagam, J.S. Eid, A. Meller, J. Chem. Phys. 122 (2005) 061103. DOI:10.1063/1.1854120 |
| [69] |
A. Pomorski, A. Krezel, Chembiochem 12 (2011) 1152-1167. DOI:10.1002/cbic.201100114 |
| [70] |
R.A. Scheck, A. Schepartz, Acc. Chem. Res. 44 (2011) 654-665. DOI:10.1021/ar2001028 |
| [71] |
C. Huang, Q. Yin, W. Zhu, et al., Angew. Chem. Int. Ed. 50 (2011) 7551-7556. DOI:10.1002/anie.201101317 |
| [72] |
C. Huang, Q. Yin, J. Meng, et al., Chem.-Eur. J. 19 (2013) 7739-7747. DOI:10.1002/chem.201300567 |
| [73] |
N. Fu, D. Su, J.R. Cort, et al., J. Am. Chem. Soc. 135 (2013) 3567-3575. DOI:10.1021/ja3117284 |
| [74] |
Z. Yang, D.H. Kang, H. Lee, et al., Bioconjugate Chem. 29 (2018) 1446-1453. DOI:10.1021/acs.bioconjchem.8b00128 |
| [75] |
M. Dahmoun, K. Boman, S. Cajander, et al., J. Clin. Endocrinol. Metab. 84 (1999) 1737-1743. |
| [76] |
C.B. Thompson, Science 267 (1995) 1456-1462. DOI:10.1126/science.7878464 |
| [77] |
D. Park, A.S. Don, T. Massamiri, et al., J. Am. Chem. Soc. 133 (2011) 2832-2835. DOI:10.1021/ja110226y |
| [78] |
Y. Liu, D. Duan, J. Yao, et al., J. Med. Chem. 57 (2014) 5203-5211. DOI:10.1021/jm500221p |
| [79] |
X. Zhang, Y. Xiao, X. Qian, Angew. Chem. Int. Ed. 47 (2008) 8025-8029. DOI:10.1002/anie.200803246 |
| [80] |
Z. Zhou, M. Yu, H. Yang, et al., Chem. Commun. (2008) 3387-3389. |
| [81] |
C. Huang, T. Jia, M. Tang, et al., J. Am. Chem. Soc. 136 (2014) 14237-14244. DOI:10.1021/ja5079656 |
| [82] |
A.S. Klymchenko, Acc. Chem. Res. 50 (2017) 366-375. DOI:10.1021/acs.accounts.6b00517 |
| [83] |
J. Mei, N.L. Leung, R.T. Kwok, et al., Chem. Rev. 115 (2015) 11718-11940. DOI:10.1021/acs.chemrev.5b00263 |
| [84] |
D. Su, C.L. Teoh, L. Wang, et al., Chem. Soc. Rev. 46 (2017) 4833-4844. DOI:10.1039/C7CS00018A |
| [85] |
Z. Guo, A. Shao, W.H. Zhu, J. Mater. Chem. C 4 (2016) 2640-2646. DOI:10.1039/C5TC03369A |
| [86] |
J. Gong, P. Wei, Y. Su, et al., Chin. Chem. Lett. 29 (2018) 1493-1496. DOI:10.1016/j.cclet.2018.05.043 |
| [87] |
Y. Wang, X.F. Yang, Y. Zhong, et al., Chem. Sci. 7 (2016) 518-524. DOI:10.1039/C5SC02824H |
| [88] |
Y. Wang, Y. Zhong, Q. Wang, et al., Anal. Chem. 88 (2016) 10237-10244. DOI:10.1021/acs.analchem.6b02923 |
| [89] |
F. Liu, H.J. Liu, X.J. Liu, et al., Anal. Chem. 89 (2017) 11203-11207. DOI:10.1021/acs.analchem.7b03724 |
| [90] |
S. Zhang, G. Chen, Y. Wang, et al., Anal. Chem. 90 (2018) 2946-2953. DOI:10.1021/acs.analchem.7b05429 |
| [91] |
A.C. Kimmelman, E. White, Cell Metab. 25 (2017) 1037-1043. DOI:10.1016/j.cmet.2017.04.004 |
| [92] |
Y. He, J. Shin, W. Gong, et al., Chem. Commun. 55 (2019) 2453-2456. DOI:10.1039/C9CC00300B |
| [93] |
K. Renault, J.W. Fredy, P.Y. Renard, C. Sabot, Bioconjugate Chem. 29 (2018) 2497-2513. DOI:10.1021/acs.bioconjchem.8b00252 |
| [94] |
X. Chen, Y. Zhou, X. Peng, J. Yoon, Chem. Soc. Rev. 39 (2010) 2120-2135. DOI:10.1039/b925092a |
| [95] |
S. Girouard, M.H. Houle, A. Grandbois, et al., J. Am. Chem. Soc. 127 (2005) 559-566. DOI:10.1021/ja045742x |
| [96] |
J. Guy, K. Caron, S. Dufresne, et al., J. Am. Chem. Soc. 129 (2007) 11969-11977. DOI:10.1021/ja0738125 |
| [97] |
F. Gao, H. Chen, S. Xu, et al., Talanta 116 (2013) 508-513. DOI:10.1016/j.talanta.2013.07.032 |
| [98] |
X. Pan, Z. Liang, J. Li, et al., Chem.-Eur. J. 21 (2015) 2117-2122. DOI:10.1002/chem.201405349 |
| [99] |
J. Guy, R. Castonguay, Campos-Reales Pineda N.B., et al., Mol. BioSyst. 6 (2010) 976-987. DOI:10.1039/b918205e |
| [100] |
K. Caron, V. Lachapelle, J.W. Keillor, Org. Biomol. Chem. 9 (2011) 185-197. DOI:10.1039/C0OB00455C |
| [101] |
Y. Chen, C.M. Clouthier, K. Tsao, etal., Angew.Chem.Int.Ed. 53 (2014) 13785-13788. DOI:10.1002/anie.201408015 |
| [102] |
Y. Chen, K. Tsao, S.L. Acton, J.W. Keillor, Angew. Chem. Int. Ed. 130 (2018) 12570-12574. DOI:10.1002/ange.201805482 |
| [103] |
J.J. Lee, S.C. Lee, D. Zhai, et al., Chem. Commun. 47 (2011) 4508-4510. DOI:10.1039/c1cc10362h |
| [104] |
L.W. Miller, Y. Cai, M.P. Sheetz, V.W. Cornish, Nat. Methods 2 (2005) 255-257. DOI:10.1038/nmeth749 |
| [105] |
A. Gautier, A. Juillerat, C. Heinis, et al., Chem. Biol. 15 (2008) 128-136. DOI:10.1016/j.chembiol.2008.01.007 |
| [106] |
I. Nikic, T. Plass, O. Schraidt, et al., Angew. Chem. Int. Ed. 53 (2014) 2245-2249. DOI:10.1002/anie.201309847 |
| [107] |
S. Leng, Q. Qiao, L. Miao, et al., Chem. Commun. 53 (2017) 6448-6451. DOI:10.1039/C7CC01483J |
| [108] |
G.V. Los, L.P. Encell, M.G. McDougall, et al., ACS Chem. Biol. 3 (2008) 373-382. DOI:10.1021/cb800025k |
| [109] |
Y. Hori, T. Norinobu, M. Sato, et al., J. Am. Chem. Soc. 135 (2013) 12360-12365. DOI:10.1021/ja405745v |
| [110] |
S.L. Kuan, T. Wang, T. Weil, Chem.-Eur. J. 22 (2016) 17112-17129. DOI:10.1002/chem.201602298 |
| [111] |
G. Liu, J. Hu, S. Liu, Chem.-Eur. J. 24 (2018) 16484-16505. DOI:10.1002/chem.201801290 |
| [112] |
S. Peng, B. Zhang, X. Meng, et al., J. Med. Chem. 58 (2015) 5242-5255. DOI:10.1021/acs.jmedchem.5b00410 |
| [113] |
G. Hu, B. Zhang, P. Zhou, et al., J. Mater. Chem. B 7 (2019) 2696-2702. DOI:10.1039/C8TB03101K |
| [114] |
H.M. Kim, B.R. Cho, Chem. Rev. 115 (2015) 5014-5055. DOI:10.1021/cr5004425 |