 2019, Vol. 30
2019, Vol. 30 


b Key Laboratory of Environment-Friendly Chemistry and Application of Ministry of Education, Laboratory of Biochemistry, College of Chemistry, Xiangtan University, Xiangtan 411105, China
Multi-walled carbon nanotubes (MWCNTs) might be used in many commercial products, such as supercapacitors, automobiles and packaging [1-3]. Meanwhile, MWCNTs could be used as novel nanomaterials (NMs) in biomedicine because of their unique physicochemical properties. For example, extensive studies have shown that MWCNTs as well as other carbon based NMs are promising nanoplateforms for cancer therapy [4]. MWCNTs have also been shown great potential as biosensors to quickly and legitimately detect biomolecules [5]. Owing to their extraordinary properties such as high efficient drug loading capacity, MWCNTs could be used as nanocarriers to deliver bioactive compounds and drugs [6]. However, to ensure the safe use of MWCNTs, it is necessary to understand the potential toxicity of MWCNTs to human endothelial cells, since these applications of MWCNTs could inevitably increase the contact of NMs to human blood vessels [7, 8].
Recently, we found that MWCNTs with short length were significantly less toxic to human umbilical vein endothelial cells (HUVECs) compared with the ones with long length, which suggested that short MWCNTs might be safer nanocarriers [9]. However, it should be pointed out that pristine MWCNTs are not commonly used as nanocarriers, as nanocarriers are normally surface-functionalized with chemical groups, for instance hydroxyl groups and carboxyl groups, to improve their biocompatibility and capacity to conjugate molecules [4, 10]. But the biocompatibility of hydroxylated MWCNTs (h-MWCNTs) and carboxylated MWCNTs (c-MWCNTs) to human endothelial cells was less studied, although some studies showed previously that h-MWCNTs and cMWCNTs were more biocompatible to liver cells compared with pristine MWCNTs [11, 12]. Meanwhile, previous studies, particularly in vitro studies, might overlook the importance of taking the disease-associated conditions into account. For example, ER stress is an adaptive response to restore homeostasis of ER, and prolonged ER stress has been implicated in human diseases, like cancer and vascular diseases [13]. Therefore, to better understand the potential adverse effects of MWCNTs as nanocarriers for disease therapy, the diversity of endothelial cells under ER stress conditions should be considered [7, 8].
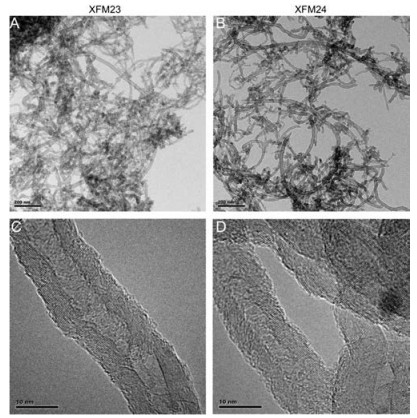
Therefore this study stressed HUVECs by pre-treatment with ER stress inducer thapsigargin (TG), and investigated the toxicity of hMWCNTs and c-MWCNTs to HUVECs both under normal and stressed conditions. The h-MWCNTs (code XFM23) and c-MWCNTs (code XFM24) were purchased from Nanjing XFNANO Materials Tech Co., Ltd. Some of the physicochemical properties, including purity, contents of functional groups, primary size, specific surface area, tap density, true density, and electric conductivity, have been measured by the supplier and were summarized in Table S1 in Supporting information. The contents of hydroxyl groups of XFM23 and carboxyl groups of XFM24 were measured by XPS by the supplier as 1.76 wt% and 1.23 wt%, respectively. The TEM images from this study (Fig. 1) suggested that XFM23 and XFM24 had similar morphologies. The hydrodynamic size and polydispersity index (PDI) of XFM23 and XFM24 in water and medium were also similar, but XFM23 had low levels of Zeta potential, whereas XFM24 were negatively charged both in water and medium (Table 1 and Fig. S1 in Supporting information).

|
Download:
|
| Fig. 1. The TEM images of XFM23 (h-MWCNTs) (A, C) and XFM24 (c-MWCNTs) (B, D). | |
|
|
Table 1 The average hydrodynamic size, Zeta potential and PDI of XFM23 (h-MWCNTs) and XFM24 (c-MWCNTs) in water and medium.a |

{kind=link}
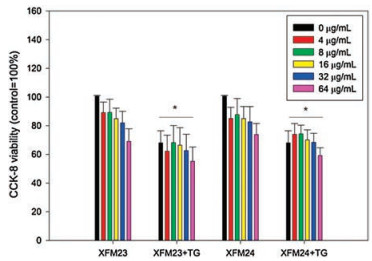
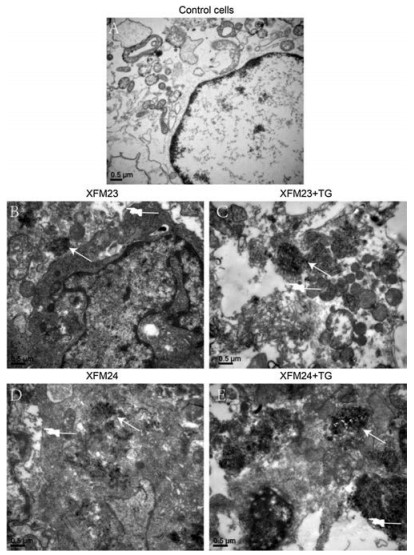
The cytotoxicity of h-MWCNTs and c-MWCNTs to HUVECs with or without TG pre-treatment was evaluated by CCK-8 assay. As shown in Fig. 2, there was a modest decrease of cellular viability following exposure to various concentrations of MWCNTs, but ANOVA indicated no significant effect of concentrations (P > 0.05). There was a single-factor effect of TG (P < 0.01), but ANOVA suggested no interaction between TG and MWCNTs (P > 0.05). To further investigate the ultrastructural changes of HUVECs due to exposure, transmission electron microscope (TEM) was used. The representative TEM images indicated that control cells had clear ultrastructures (Fig. 3A). However, HUVECs exposed to XFM23 (Fig. 3B) and XFM24 (Fig. 3D) showed ultrastructural changes. For example, the cells became relatively dark and appeared to have vacuolation (fingers in Figs. 3B and D). The intracellular vacuolation was more obvious in cells pre-treated with TG (Figs. 3C and E). The TEM images also indicated the internalization of MWCNTs as agglomerates and/or aggregates (arrows in Figs. 3B and D). TG pretreated HUVECs appeared to internalize more MWCNTs (Figs. 3C and E), although it could not be quantitatively measured.

|
Download:
|
| Fig. 2. The cellular viability of HUVECs. HUVECs were pre-incubated with or without 500 nmol/L TG for 1 h, and then exposed to various concentrations of XFM23 (h-MWCNTs) and XFM24 (c-MWCNTs) for 24 h. The cellular viability was measured by CCK-8 assay. Data represent mean ± SEM of three independent experiments (n = 3 for each). * P < 0.01, compared with control, ANOVA. | |
{kind=link}

|
Download:
|
| Fig. 3. The TEM images of HUVECs. HUVECs were first incubated with medium with or without 500 nmol/L TG for 1 h, and then exposed to 64 μg/mL XFM23 (h-MWCNTs) or XFM24 (c-MWCNTs) for 24 h. HUVECs not treated with TG or MWCNTs were served as control. All the samples were then imaged by TEM to indicate the ultrastructural changes of cells and internalization of MWCNTs. (A) control cells; (B) HUVECs exposed to XFM23 only; (C) HUVECs first treated with TG and then exposed to XFM23; (D) HUVECs exposed to XFM24 only; (E) HUVECs first treated with TG and then exposed to XFM24. The arrows indicate the MWCNTs internalized into HUVECs, and the fingers indicate intracellular vacuolation. | |
{kind=link}
The release of cytokines into cell culture medium and THP-1 adhesion to HUVECs was measured to indicate inflammatory responses. For IL-6 (Fig. 4A), only TG pre-treatment (P < 0.01), but not MWCNTs (P > 0.05), significantly promoted the release of IL-6. For sICAM-1 (Fig. 4B), its release was significantly promoted after exposure to 64 μg/mL XFM23 (P < 0.01) or XFM24 (P < 0.05). For sVCAM-1 (Fig. 4C), its release was significantly promoted after exposure to 16 μg/mL XFM23 (P < 0.05), 64 μg/mL XFM23 (P < 0.01), or 64 μg/mL XFM24 (P < 0.05). Pre-treatment with TG did not significantly influence MWCNT-induced sICAM-1 or sVCAM-1 release (P > 0.05). Moreover, ANOVA indicated no interaction between MWCNTs and TG on the release of IL-6, sICAM-1 or sVCAM-1 (P > 0.05). For THP-1 monocyte adhesion (Fig. 4D), exposure to 64 mg/mL XFM23 or XFM24 modestly promoted THP-1 adhesion onto HUVECs, but ANOVA indicated no effect of concentrations of MWCNTs (P > 0.05). After pre-treatment with TG, all the concentrations of XFM23 and XFM24 significantly promoted THP-1 adhesion (P < 0.01).

|
Download:
|
| Fig. 4. The release of IL-6 (A), sICAM-1 (B), sVCAM-1 (C), and the adhesion of THP-1 monocytes onto HUVECs (D). HUVECs were pre-incubated with or without 500 nmol/L TG for 1 h, and then exposed to various concentrations of XFM23 (h-MWCNTs) and XFM24 (c-MWCNTs) for 24 h. The concentrations of cytokines released into supernatants were determined by ELISA. THP-1 adhesion was measured by using a fluorescent probe CellTracker Green. Data represent mean ± SEM of three independent experiments (n = 3 for each). * P < 0.05, compared with control, ANOVA (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article). | |
{kind=link}
All of the data presented above indicate that the toxicity of XFM23 and XFM24 up to 64μg/mL was relativelymodest. The concentrations of MWCNTs were selected according toourprevious study [9], as these concentrations might be reached in vivo under biomedical uses. According to the supplier, the length of the MWCNTs used in this study ranges from 0.5 μm to 2 μm, and we recently found that pristine MWCNTs with this length were less toxic to HUVECs compared with MWCNTs with length of 10 μm to 30 μm [9]. These data are alsoin agreementwith previous studies showing thatfibers with relatively short length, either functionalized or non-functionalized, were generally less toxic to endothelial cells [14-17].
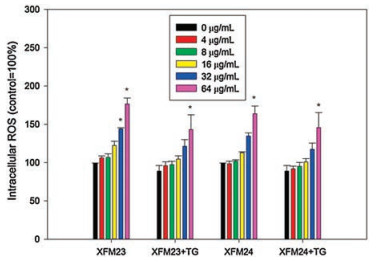
Intracellular ROS was measured to indicate oxidative stress induced by MWCNTs under different conditions. As shown in Fig. 5, exposure to MWCNTs was associated with dose-dependent increase of intracellular ROS, and significantly increased intracellular ROS was observed after exposure to 32 μg/mL XFM23 (P < 0.05), 64 μg/mL XFM23 (P < 0.01) or 64 μg/mL XFM24 (P < 0.01). Pre-treatment with TG did not further enhance MWCNT-induced ROS, and 64 μg/mL XFM23 or XFM24 (P < 0.05) significantly promoted intracellular ROS in TG pre-treated HUVECs.

|
Download:
|
| Fig. 5. The increase of intracellular ROS in HUVECs. HUVECs were pre-incubated with or without 500 nmol/L TG for 1 h, and then exposed to various concentrations of XFM23 (h-MWCNTs) and XFM24 (c-MWCNTs) for 24 h. The intracellular ROS was determined by using a fluorescent probe DCFH-DA. Data represent mean ± SEM of three independent experiments (n = 3 for each). * P < 0.05, compared with control, ANOVA. | |
{kind=link}
This study stressed HUVECs with ER stress inducer TG to investigate a combined effect of TG pre-treatment and MWCNTs. The concentration of TG was selected according to previous studies, which showed that 24 h exposure to 500 μmol/L could efficiently induce pronounced ER stress in vitro [18, 19]. Here the exposure time was shortened to 1 h in order to evaluate if MWCNTs could promote the toxic effects of TG, since convincing data showed that NM exposure could link ER stress to the adverse effects of NMs [20]. To indicate the activation of ER stress-apoptosis pathway, the typical ER stress-apoptosis genes and proteins were measured by real time RT-PCR and Western blot, respectively. The endpoints included mRNA levels of DDIT3 (DNA damage-inducible transcript 3), XBP-1s (spliced X-box binding protein 1) and ATF6 (activating transcription factor 6), as well as protein levels of chop (C/EBP homologous proteins) and p-chop. The mRNA levels of apoptotic gene CASP3 (caspase 3) and protein levels of pro-casp3 and cleaved casp3 were also measured by real time RT-PCR and Western blot, respectively. For the expression of DDIT3 (Fig. 6A), exposure to XFM24 led to approximately 80% increase of DDIT3 expression compared with control. TG pre-treatment significantly promoted the expression of DDIT3 (P < 0.01), and XFM23 and XFM24 exposed TG pre-treated HUVECs had significantly higher expression of DDIT3 (P < 0.01). For the expression of XBP-1s (Fig. 6B), the expression was significantly elevated in XFM23 or XFM24 exposed HUVECs as well as TG pretreated HUVECs (P < 0.01 for all). Moreover, the expression of XBP-1s in XFM24 exposed TG pre-treated HUVECs was significantly higher than that in XFM24 exposed cells (P < 0.01). For the expression of ATF6 (Fig. 6C), its expression was only significantly promoted by TG pre-treatment (P < 0.01) but not MWCNTs (P > 0.05). Also, the expression of ATF6 in MWCNT exposed TG pre-treated HUVECs was significantly higher than that in MWCNT exposed cells (P < 0.01). For the expression of CASP3 (Fig. 6D), its expression was only significantly promoted in TG pre-treated cells or MWCNT exposed TG pre-treated cells (P < 0.01). For the protein levelsof p-chop, chop (Fig. 6E) and pro-casp3 (Fig. 6F), the ratio of p-chop/chop was only modestly increased by XFM24 exposure. Cleaved casp3 proteins were not observed in any of the group (Fig. S2 in Supporting information).

|
Download:
|
| Fig. 6. The mRNA levels of DDIT3 (A), XBP-1s (B), ATF6 (C), CASP3 (D) and protein levels of chop, p-chop (E) and pro-casp3 (F). HUVECs were pre-incubated with or without 500 nmol/L TG for 1 h, and then incubated without (control) or with 64 μg/mL XFM23 (h-MWCNTs) and XFM24 (c-MWCNTs) for 24 h. The expression of DDIT3, XBP-1s, ATF6 and CASP3 was determined by real time RT-PCR. Data represent mean ± SEM of three independent experiments (n = 3 for each). The protein levels of p-chop, chop and procasp3 were measured by Western blot (the unedited Western blot images are shown in Fig. S2). * P < 0.05, compared with control; # P < 0.05, compared between cells with or without TG pre-treatment, ANOVA. | |
{kind=link}
The above results suggested that pre-treatment with TG significantly induced toxic effects to HUVECs probably through the activation of ER stress-apoptosis pathway. However, ANOVA indicated no interaction between TG pre-treatment and MWCNTs on all of the endpoints (P > 0.05). In our previous study, it was shown that the presence of TG did not significantly promote MWCNT-induced monocyte adhesion to HUVECs [9]. In another study, the toxicity of polymeric micelles was not enhanced by coexposure of TG [21]. However, the cytotoxicity of ZnO NMs to THP- 1 macrophages was significantly promoted with the presence of TG [22]. Therefore, it is possible that the influence of TG on the toxicity of NMs is dependent on the compositions of NMs. This might be related with the potential of NMs to induce ER stress. Metal based NMs, such as ZnO NMs, have been shown to induce a pronounced ER stress both in vivo and in vitro [23, 24]. In contrast, NMs with tubular structures, such as MWCNTs and anodic alumina nanotubes, generally induced more modest ER stress responses [25, 26], and therefore activation of ER stress by TG might have little to no effect to alter the toxicity of MWCNTs. However, this conclusion has to be confirmed in the future by comparing more types of NMs.
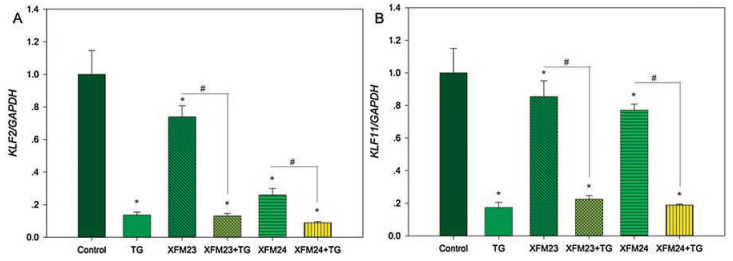
This study also measured the expression of KLF2 (Kruppel-like factor 2) and KLF11 by real time RT-PCR. KLF belong to the transcript factors that could regulate pluripotency [27] and vascular homeostasis [28]. In our recent study, KLF4 expression could be induced by metal-based NPs in human vascular smooth muscle cells, which suggested that NP exposure might modulate KLF expression [29]. In this study, the expression of KLF2 (Fig. 7A) was significantly decreased after exposure to 64 μg/mL XFM23 or XFM24 (P < 0.05). TG pre-treatment also significantly decreased the expression (P < 0.01), and the expression of KLF2 was significantly lower in MWCNT exposed TG pre-treated HUVECs compared with that in HUVECs only exposed to MWCNTs (P < 0.01). Similar trend was observed for KLF11 (Fig. 7B). KLF11 expression was significantly lowered after exposure to 64 μg/mL XFM23 (P < 0.05) or XFM24 (P < 0.01), and its expression was further significantly lowered by TG pre-treatment (P < 0.01).

|
Download:
|
| Fig. 7. The expression of KLF2 (A) and KLF4 (B). HUVECs were pre-incubated with or without 500 nmol/L TG for 1 h, and then incubated without (control) or with 64 μg/mL XFM23 (h-MWCNTs) and XFM24 (c-MWCNTs) for 24 h. The expression of KLF2 and KLF4 was determined by real time RT-PCR. Data represent mean ± SEM of three independent experiments (n = 3 for each). *P < 0.05, compared with control; #P < 0.01, compared between cells with or without TG pre-treatment, ANOVA. | |
{kind=link}
KLF2 has been well-studied as a vasoprotective transcription factor that could positively up-regulate the generation of vascular signaling molecules, such as NO [30, 31]. KLF11 is less studied, but some studies indicated KLF11 could also be protective to endothelial cells by the inhibition of inflammation [29, 32]. Therefore, the pronounced down-regulation of KLF2 and KLF11 by TG pre-treatment might render HUVECs more sensitive to MWCNT exposure. This could be evidenced by that MWCNTexposed HUVECs exhibited ultrastructural changes, which appeared to be more obvious with TG pre-treatment (Fig. 3). Again, the results showed no synergistic effect of TG pre-treatment and MWCNTs on KLF expression, that MWCNTs did not further alter the effects of TG.
In summary, the results from this study showed that up to 64 μg/mL h-MWCNTs and c-MWCNTs only induced modest toxic effects to HUVECs. Pre-treatment with TG induced toxic effects that are likely associated with ER stress, but a synergistic effect of TG pre-treatment and MWCNTs was not observed. This indicated that the toxicity of h-MWCNTs and c-MWCNTs might not be exacerbated under ER stressed conditions. Nevertheless, many different types of NMs based on inorganic materials, such as Au nanorods [33], carbon nanodots [34] and quantum dots [35], could be used for biomedical purposes. It is still suggested that their toxicity to endothelial cells, particularly under stressed conditions, should be carefully evaluated to understand their potential adverse effects during biomedical uses.
AcknowledgmentsThis work was financially supported by College Outstanding Young and Middle-aged Innovative Research Team Project of Hubei Province (No. T201717), the Health and Family Planning Commission Scientific Research Project of Hubei Province (No. WJ2017Z027).
Appendix A. Supplementary dataSupplementary material related to this article can be found, in the online version, at doi:https://doi.org/10.1016/jcclet.2018.12.011.
| [1] |
L. Liu, Z. Niu, J. Chen, Chin. Chem. Lett. 29 (2018) 571-581. DOI:10.1016/j.cclet.2018.01.013 |
| [2] |
T. Kuang, L. Chang, F. Chen, et al., Carbon 105 (2016) 305-313. DOI:10.1016/j.carbon.2016.04.052 |
| [3] |
D. Fu, T. Kuang, Y.C. Yen, et al., J. Cell Plast. 53 (2017) 273-285. DOI:10.1177/0021955X16651253 |
| [4] |
J. Saleem, L. Wang, C. Chen, Adv. Healthc. Mater. (2018) e1800525. |
| [5] |
S. Alim, J. Vejayan, M.M. Yusoff, A.K.M. Kafi, Biosens. Bioelectron. 121 (2018) 125-136. DOI:10.1016/j.bios.2018.08.051 |
| [6] |
M. Mohajeri, B. Behnam, A. Sahebkar, J. Cell. Physiol. 234 (2018) 298-319. |
| [7] |
Y. Cao, Y. Gong, L. Liu, et al., J. Appl. Toxicol. 37 (2017) 1359-1369. DOI:10.1002/jat.v37.12 |
| [8] |
M.I. Setyawati, C.Y. Tay, D. Docter, R.H. Stauber, D.T. Leong, Chem. Soc. Rev. 44 (2015) 8174-8199. DOI:10.1039/C5CS00499C |
| [9] |
J. Long, Y. Xiao, L. Liu, Y. Cao, Int. J. Nanobiotechnology Pharm. 15 (2017) 80. DOI:10.1186/s12951-017-0318-x |
| [10] |
P.T. Wong, S.K. Choi, Chem. Rev. 115 (2015) 3388-3432. DOI:10.1021/cr5004634 |
| [11] |
Z. Liu, Y. Liu, D. Peng, J. Biomed. Mater. Res. A 103 (2015) 2770-2777. DOI:10.1002/jbm.a.v103.8 |
| [12] |
Z. Liu, Y. Liu, D. Peng, Environ. Toxicol. Pharmacol. 47 (2016) 124-130. DOI:10.1016/j.etap.2016.09.013 |
| [13] |
L. Ozcan, I. Tabas, Annu. Rev. Med. 63 (2012) 317-328. DOI:10.1146/annurev-med-043010-144749 |
| [14] |
S. Tang, Y. Tang, L. Zhong, et al., J. Appl. Toxicol. 32 (2012) 900-912. DOI:10.1002/jat.v32.11 |
| [15] |
Y. Cao, N.R. Jacobsen, P.H. Danielsen, et al., Toxicol. Sci. 138 (2014) 104-116. DOI:10.1093/toxsci/kft328 |
| [16] |
D.A. Donkor, X.S. Tang, Biomaterials. 35 (2014) 3121-3131. DOI:10.1016/j.biomaterials.2013.12.075 |
| [17] |
A. Celluzzi, A. Paolini, D'Oria V., et al., Int. J. Nanomed. Nanosurg. 13 (2017) 1-18. DOI:10.2147/IJN |
| [18] |
R. Ying, X.Q. Wang, Y. Yang, et al., Life Sci. 144 (2016) 208-217. DOI:10.1016/j.lfs.2015.11.025 |
| [19] |
H. Yoshino, Y. Kumai, I. Kashiwakura, Mol. Med. Rep. 15 (2017) 2867-2872. DOI:10.3892/mmr.2017.6298 |
| [20] |
Y. Cao, J. Long, L. Liu, et al., Life Sci. 186 (2017) 33-42. DOI:10.1016/j.lfs.2017.08.003 |
| [21] |
F. Liu, H. Huang, Y. Gong, et al., Chem. Biol. Interact. 263 (2017) 46-54. DOI:10.1016/j.cbi.2016.12.014 |
| [22] |
G. Chen, Y. Shen, X. Li, et al., Environ. Toxicol. Pharmacol. 50 (2017) 103-110. DOI:10.1016/j.etap.2017.01.020 |
| [23] |
R. Chen, L. Huo, X. Shi, et al., ACS Nano 8 (2014) 2562-2574. DOI:10.1021/nn406184r |
| [24] |
X. Yang, H. Shao, W. Liu, et al., Toxicol. Lett. 234 (2015) 40-49. DOI:10.1016/j.toxlet.2015.02.004 |
| [25] |
S. Chang, X. Zhao, S. Li, et al., Ecotoxicol. Environ. Saf. 161 (2018) 569-577. DOI:10.1016/j.ecoenv.2018.06.025 |
| [26] |
Y. Wang, G. Kaur, A. Zysk, et al., Biomaterials 46 (2015) 117-130. DOI:10.1016/j.biomaterials.2014.12.008 |
| [27] |
M.K. Farrugia, D.B. Vanderbilt, M.A. Salkeni, J.M. Ruppert, Cancer Res. 76 (2016) 1677-1682. DOI:10.1158/0008-5472.CAN-15-1806 |
| [28] |
M. Wang, Q. Yang, J. Long, et al., Int. J. Nanomed. Nanosurg. 13 (2018) 8037-8049. DOI:10.2147/IJN |
| [29] |
Y. Fan, H. Lu, W. Liang, et al., J. Mol. Cell Biol. 9 (2017) 352-363. DOI:10.1093/jmcb/mjx037 |
| [30] |
H. Chu, H. Li, X. Guan, et al., Mol. Med. Rep. 17 (2018) 5708-5714. |
| [31] |
X. Wang, Z. Wu, Y. He, et al., Mol. Immunol. 101 (2018) 245-250. DOI:10.1016/j.molimm.2018.07.008 |
| [32] |
C. Glineur, B. Gross, B. Neve, et al., Arterioscler. Thromb. Vasc. Biol. 33 (2013) 621-628. DOI:10.1161/ATVBAHA.112.300665 |
| [33] |
F. Zhu, G. Tan, Y. Jiang, Z. Yu, F. Ren, Biomater. Sci. 6 (2018) 2905-2917. DOI:10.1039/C8BM00691A |
| [34] |
Y. Yang, X. Wang, G. Liao, et al., J. Colloid Interface Sci. 509 (2018) 515-521. DOI:10.1016/j.jcis.2017.09.007 |
| [35] |
P. Zhao, Q. Xu, J. Tao, et al., Wiley interdiscip, Rev. Nanomed. Nanobiotechnol. (2018) e1483. |