 2019, Vol. 30
2019, Vol. 30 


b State Key Laboratory of Bioactive Substance and Function of Natural Medicines, Institute of Materia Medica, Chinese Academy of Medical Sciences and Peking Union Medical College, Beijing 100050, China
Benzylisoquinoline alkaloids represent one class of the most important alkaloids, which mainly exists in families of Ranunculaceae, Rutaceae, Menispermanceae, Papaveraceae, Berberidaceae etc. [1]. These compounds possess various bioactivities like antialzheimer, antihyperlipidemics, anti-HIV, antibacterial, anticancer, immunomodulatory and cholesterol lowering effect [2]. However, low bioavailability and acute toxicity influenced their therapeutic effects [3]. Glycosylation was an effective way to improve bioavailability and even enhance bioactivity of benzylisoquinoline alkaloids. For instance, glycosylation of tetrahydroprotoberberines could maintain and even enhance the inhibitory effects on tissue factor procoagulant activity [4]. Moreover, introducing sugar moieties to the 9–OH of berberine derivatives could dramatically improve the dissolution rate of the aglycon in plasma, thus increase its bioavailability [3a].
Enzymatic glycosylation catalyzed by glycosyltransferases (GTs) is generally recognized as a powerful approach for the synthesis of bioactive glycosides [5]. GTs are able to transfer sugar moieties from activated donors to acceptor molecules with regioselectivity and stereoselectivity, without protection/ deprotection of functional groups and use of toxic reaction agents [6]. To date, a number of GTs have been discovered, some of which have also been used as enzymatic tools in syntheis of diverse glycosides. An engineered oleandomycin GT (ASP OleD) from Streptomyces antibioticus exhibited unprecedented catalytic promiscuity and was able to recognize 71 diverse acceptors out of 137 drug-like compounds [5b]. Besides the bacteria, GTs from fungi and plant also showed broad substrate spectra and high catalytic efficiency. MhGT1 from Mucor hiemalis has the capability for regio- and stereospecific glucosylation of 72 structurally diver drug-like compounds [5f]. In our previous work, a promiscuous plant GT (UGT73AE1) from Carthamus tinctorius, which could catalyze to generate diverse O-, S- and N-glucosides was also discovered [5c]. However, few of the GTs have been used as enzymatic tools for glycosylation of benzylisoquinoline alkaloids. Thus, the discovery of GTs with catalytic promiscuity and novel specificity to diverse bioactive benzylisoquinoline alkaloids is of necessity in practice.
In this work, we aim to screen for GTs with catalytic promiscuity to various benzylisoquinoline alkaloids, which possess a basic isoquinoline unit, to expand the application of GTs in generating novel glycosides. Thus, three new GTs UGT90A14, UGT71AE1 and UGT84A33 from C. tinctorius were succesfully mined and employed as enzymatic tools in glycosylation of diverse bioactive benzylisoquinoline alkaloids.
C. tinctorius is a famous medicinal plant and a wide variety of structurally diverse glycosides have been isolated from its florets [7]. The diversity of glycosyated secondary metabolites implied the existence of various GTs, which inspired us to seek GTs with novel catalytic properties. In our previous work, UGT73AE1 with promiscuous O-glycosylation activity and UGT71E5 with permissive N-glycosylation have been obtained [5c, 5e]. Thus, to clone novel GTs from C. tinctorius, a degenerate PCR primer for 5' RACE was designed based on the conserved PSPG (plant secondary product glycosyltransferases) motif of five permissive plant GTs as shown in Fig. S1 (Supporting information) [5e]. Combined with 3' RACE, 17 new different C. tinctorius glycosyltransferase (CtGTs) genes were successfully cloned by RT-PCR amplification using the total RNA from florets of C. tinctorius as a template and heterologously expressed in Escherichia coli.
To screen the glycosylation capability of the recombinant CtGTs in vitro, UDP-glucose (UDP-Glc) along with 5-hydroxyisoquinoline (1), 6-hydroxyisoquinoline (2), 7-hydroxyisoquinoline (3) and 8-hydroxyisoquinoline (4), which are typical and basic structural units of benzylisoquinoline alkaloids, were used in enzymatic assays. The screening reactions (50 mmol/L Tris-HCl, pH 7.4; 0.5 mmol/L UDP-Glc; 0.25 mmol/L aglycons; 500 μg of crude CtGTs; 30 ℃, 6 h) were analyzed by HPLC-UV/MS (high-performance liquid chromatography with ultraviolet/mass spectrometry detection). Of the 17 recombinant CtGTs, only CtGT31, CtGT38 and CtGT42 showed glycosylation activity to all the four isoquinolines with relative high conversion rates (Figs. 1A and B). Control reactions lacking either enzyme or UDP-Glc confirmed that the reactions were dependent upon both the enzyme and the sugar donor.

|
Download:
|
| Fig. 1. The glycosylation of diverse hydroxyl isoquinolines catalyzed by the recombinant UGT84A33, UGT71AE1 and UGT90A14. (A) The structures of the hydroxyl isoquinolines used as acceptors for screening reactions; (B) Percent conversions of glucosylated products catalyzed by UGT84A33 (CtGT42), UGT71AE1 (CtGT38) and UGT90A14 (CtGT31), respectively; (C) SDS-PAGE analysis of the three purified recombinant CtGTs. | |
{kind=link}
The cDNA sequence of CtGT31 (1455 bp, GenBank accession number MH013340) contained an ORF encoding 484 amino acids and showed the highest identity (84%) to CtUGT25, a flavonoid GT from C. tinctorius [8]. CtGT38 (1446 bp, MH013341) contained an ORF encoding 481 amino acids and showed the highest identity (80%) to CtUGT3, a flavonoid GT from C. tinctorius [8]. CtGT42 (1473 bp, MH013342) contained an ORF encoding 490 amino acids and showed the highest identity (66%) to VLRSgt, a stilbenes, flavonoids and coumarins GT from Vitis labrusca [9]. CtGT31, CtGT38 and CtGT42 were respectively further named UGT90A14, UGT71AE1 and UGT84A33 according to the UGT Naming Committee [10]. Recombinant His6-UGT90A14, His6- UGT71AE1 and His6-UGT84A33 were purified using His-tag affinity chromatography and analyzed with SDS-PAGE (Fig. 1C).
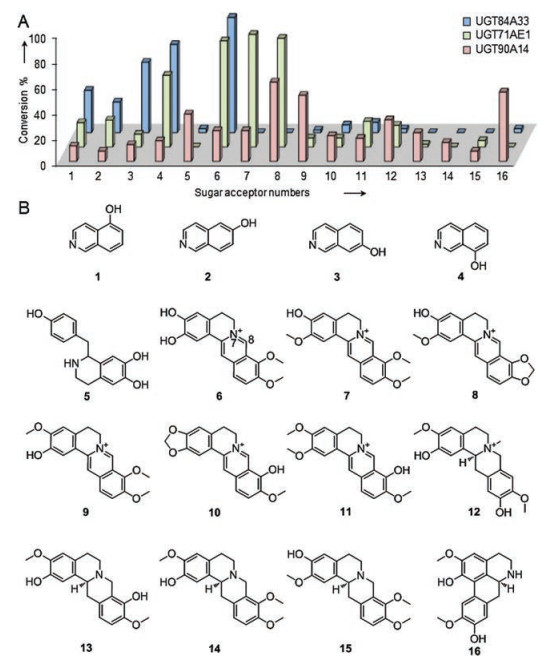
To investigate the catalytic promiscuity of UGT90A14, UGT71AE1 and UGT84A33, a benzylisoquinoline alkaloids library with several skeletons including benzyltetrahydroisoquinonline (demethylcoclaurine, 5), berberines (demethyleneberberine, 6; jatrorrhizine, 7; groenlandicine, 8; columbamine, 9; berberrubine, 10; palmaturbin, 11), protoberberines (phellodendrine, 12; scoulerine, 13; tetrahydrocolumbamine, 14; corypalmine, 15) and aporphine (norisoboldine, 16) was employed for the enzymatic assays.
Surprisingly, UGT90A14 was able to recognize all types of the benzylisoquinoline alkaloids (5–16) from the first pass analysis with HPLC-UV/MS, which suggest the incredible broad substrate spectrum of this permissive GT (Fig. 2 and Figs. S5–S20 in Supporting information). Although UGT71AE1 and UGT84A33 cannot accept all the aglycons, these two CtGTs showed much higher conversion rates to some members. For 6, UGT84A33 showed the highest glycosylation activity, while UGT71AE1 exhibited highest catalytic activity to 7, 8 and 11 (Fig. 2 and Figs. S10–S12, S15 in Supporting information). However, for 5, 9, 10 and 12-16, UGT90A14 was the perfect tool to implement the glycosylation (Fig. 2 and Figs. S9, S13, S14 and S16–S20 in Supporting information). Thus, the glycosylated modification was achieved with high efficiency by these three promiscuous GTs. In total, eight glycosylated products (6a'–9a', 10a–12a and 16a, structures are shown in Figs. S21–S52 in Supporting information) were prepared via the enzymatic scale-up reactions, seven (6a'– 9a', 11a, 12a and 16a) of which were novel compounds. During the purification process of the glycosylated products, we found the double bonds between N7 and C8 of berberines are easily reduced to dihydroberberines. Therefore, the glycosylated products of 6–9 were isolated as dihydroberberine glycosides (6a'–9a'), which was in agreement with the data of MS and NMR. The structures including the attached positions of the glucosyl moiety and anomeric stereochemistry were illuminated by HR-ESI-MS, 1H NMR, 13C NMR, HSQC and HMBC spectroscopic data analyses (Figs. S21–S52). For example, norisoboldine 9-O-β-D-glucoside (16a) gave an HR-ESI-MS ion peak at m/z 476.1901 [M + H]+, corresponding to a molecular formula of C24H30NO9 with one more glucosyl unit. In the 1H NMR and 13C NMR spectrum, the presence of the sugar moiety was suggested by proton signals 5.07 (d, 1H, J = 7.1 Hz, Glc-H1), 3.85–3.89 (m, 2H, Glc-H), 3.54 (s, 1H, overlapped, Glc-H), 3.53 (s, 1H, overlapped, Glc-H), 3.52 (s, 1H, overlapped, Glc-H), 3.48 (s, 1H, overlapped, Glc-H) and carbon signals 101.4 (Glc-1), 78.0 (Glc-5), 77.8 (Glc-3), 74.8 (Glc-2), 70.9 (Glc-4), 61.9 (Glc-6). The location of the sugar moiety was indicated by the HMBC correlaton from Glc-H1 to C-9. And the anomeric proton with large coupling constants (J = 7.1 Hz) suggested the β-configuration of glycosidic bond. Thererfore, the enzymatic product was unambiguously identified as norisoboldine 9-O-β-D-glucoside (16a). The stereoselectivities of UGT84A33, UGT71AE1 and UGT90A14 were suggested by the fact that all anomers of the glycosylated products are in the β-configuration due to anomeric protons with large coupling constants (J > 6.7 Hz, Table S2 in Supporting information).

|
Download:
|
| Fig. 2. Comparing the catalytic capabilities of UGT84A33, UGT71AE1 and UGT90A14 in glycosylation of benzylisoquinoline alkaloids. (A) Percent conversions of glycosylated products catalyzed by these three CtGTs. The library numbers are listed based on the structural scaffolds shown in part B; (B) The structures of the aglycons. The corresponding glycosylated products of 6–12 and 16 were prepared by scaled-up reactions and identified by HR-ESI-MS and NMR spectroscopic data analyses (Figs. S21–S52 in Supporting information). | |
{kind=link}
Additionally, we explored the capability of these three new GTs in recognizing diverse sugar donors including UDP-galactose, UDPglucuronic acid, UDP-N-acetylglucosamine, UDP-N-acetylgulosamine, and UDP-xylose, however, the results showed that UGT84A33, UGT71AE1 and UGT90A14were unable to recognize these five sugar donors.
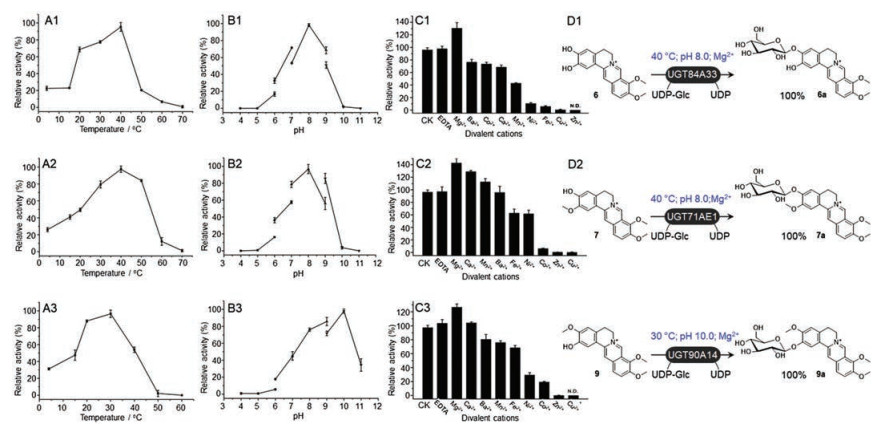
In order to improve the yields of the benzylisoquinoline alkaloid glucosides, the biochemical characteristics including temperatures, pH of buffers and divalent cations of UGT84A33, UGT71AE1 and UGT90A14, were investigated. The optimum temperature and pH of purified UGT84A33 were determined to be 40 ℃ and pH8.0 (Figs. 3A1 and B1).And those of UGT71AE1 were 40 ℃ and pH8.0 (Figs. 3A2and B2), while UGT90A14 showed the highest activity around 30 ℃ and pH 10.0 (Figs. 3A3 and B3). The three CtGTs were independent of metal ions, while Mg2+ could greatly enhance the activity (Figs. 3C1 –C3). Thus under the optimum reaction conditions of UGT84A33, UGT71 AE1 and UGT90A14, respectively, the conversion rates of 6, 7 and 9 were all improved to be 100% (Figs. 3D1–D3). Km values of UGT84A33, UGT71 AE1 and UGT90A14 toward 6, 7 and 9 were determined to be 266 μmol/L, 333 μmol/L and 112 μmol/L, respectively (Figs. S2–S4 in Supporting information).

|
Download:
|
| Fig. 3. Investigation of the effects of temperatures (A1–A3), pH of buffers (B1–B3) and divalent cations (C1–C3) to glycosylation activities of UGT84A33, UGT71AE1 and UGT90A14. Under the optimum of reaction conditions, the percent conversions of 6, 7 and 9 catalyzed by UGT84A33, UGT71AE1 and UGT90A14 were all improved to be 100% (D1–D3). | |
{kind=link}
In summary, three promiscuous GTs UGT90A14, UGT71 AE1 and UGT84A33 with capability of glycosylating benzylisoquinoline alkaloids were successfully screened from C. tinctoriuwas. The enzyme pool including UGT90A14, UGT71AE1 and UGT84A33 was developed as an enzymatic tool to efficiently synthesize diverse novel benzylisoquinoline alkaloids glycosides. Finally, 7 novel glycosylated benzylisoquinoline alkaloids along with a known one were obtained from the scale-up reactions. Furthermore, UGT90A14 exhibited robust promiscuity and stereoselectivity in glycosylation of structurally diverse derivatives of benzylisoquinoline alkaloids. Taken together, these three CtGTs constitute a powerful biocatalyst for efficient and stereoselective synthesis of novel glycosides. This study not only constitutes an enzymatic method to generate benzylisoquinoline alkaloid glycosides but also demonstrates the application prospect of plant GTs in synthesis of target natural and unnatural glycosides.
AcknowledgmentsThis work is financially supported by the National Natural Science Foundation of China (No. 81573317) and CAMS Innovation Fund for Medical Sciences (Nos. CIFMS-2016-I2M-3-012 and CIFMS-2016-I2M-2-002).
Appendix A. Supplementary dataSupplementarymaterial related to this article canbefound, in the online version, at doi: https://doi.org/10.1016/j.cclet.2018.05.010.
| [1] |
(a) L.Grycová, J.Dostál, R.Marek, Phytochemistry 68 (2007)150-175; (b) T.Hu, X.Zhang, S.Ma, et al., Chin. Chem. Lett. 20 (2009)955-957. |
| [2] |
(a) S.B.Hardik, S.Sudeep, M.Debashis, et al., Eur. J. Med. Chem. 46 (2011)1045-1049; (b) Y.Ding, X.Ye, J.Zhou, et al., Chin. J. Org. Chem. 32 (2012)677-685; (c) M.Bala, K.Pratap, P.K.Verma, et al., J. Ethnopharmacol.175 (2015)131-137; (d) H.A.Jung, B.Min, T.Yokozawa, et al., Biol. Pharm. Bull. 32 (2009)1433-1438. |
| [3] |
(a) Z.Chen, X.Ye, J.Yi, et al., Med. Chem. Res. 21 (2012)1641-1646; (b) M.M.Kheir, Y.Wang, L.Hua, et al., Food Chem. Toxicol. 48 (2010)1105-1110. |
| [4] |
H. Ge, J. Zhang, L. Chen, et al., Bioorgan. Med. Chem. 21 (2013) 62-69. DOI:10.1016/j.bmc.2012.11.002 |
| [5] |
(a) A.Minami, R.Uchida, T.Eguchi, et al., J. Am. Chem. Soc. 127 (2005)6148-6149; (b) R.W.Gantt, R.D.Goff, G.J.Williams, et al., Angew. Chem. Int. Ed. 47 (2008)8889-8892; (c) K.Xie, R.Chen, J.Li, et al., Org. Lett. 16 (2014)4874-4877; (d) K.Xie, Y.Zhang, R.Chen, et al., Tetrahedron Lett. 58 (2017)2118-2121; (e) K.Xie, R.Chen, D.Chen, et al., Adv. Synth. Catal. 359 (2017)603-608; (f) J.Feng, P.Zhang, Y.Cui, et al., Adv. Synth. Catal. 359 (2017)995-1006. |
| [6] |
(a) E.K.Lim, Chem. Eur. J. 11 (2005)5486-5494; (b) A.Luzhetskyy, C.Méndez, J.A.Salas, et al., Curr. Top. Med. Chem. 8 (2008)680-709; (c) R.W.Gantt, P.Peltier-Pain, J.S.Thorson, Nat. Prod. Rep. 28 (2011)1811-1853. |
| [7] |
(a) K.Kazuma, T.Takahashi, K.Sato, et al., Biosci. Biotechnol. Biochem. 64 (2000)1588-1599; (b) H.Yoon, H.Han, Y.J.Paik, Appl. Biol. Chem.50 (2007)175-178; (c) J.Jiang, J.He, Z.Feng, Org. Lett. 12 (2010)1196-1199. |
| [8] |
D. Guo, F. Liu, Y. Tu, et al., Plos One 11 (2016) e0158159. DOI:10.1371/journal.pone.0158159 |
| [9] |
D. Hall, V. De Luca, Plant J. 49 (2007) 579-591. DOI:10.1111/j.1365-313X.2006.02987.x |
| [10] |
(a) P.I.Mackenzie, I.S.Owens, B.Burchell, et al., Pharmacogenetics 7 (1997)255-269 (b) Homepage of the UGT Nomenclature Committee, (2018) http://www.flinders.edu.au/medicine/sites/clinical-pharmacology/ugt-homepage.cfm. |