 2019, Vol. 30
2019, Vol. 30 


b Institute of Drug Discovery Technology, Ningbo University, Ningbo 315211, China;
c Key Laboratory of Bioorganic Phosphorus Chemistry and Chemical Biology (Ministry of Education), Department of Chemistry, Tsinghua University, Beijing 100084, China
As written by Charles Darwin, "we could conceive in some warm little pond with all sorts of ammonia and phosphoric salts, light, heat, electricity etc. present, that a protein compound was chemically formed" [1]. Darwin expected that phosphate played an important role in the origin and evolution of life. Proteins, as one of the most important biological macromolecules in organism, contain a lot of amino acid residues. Therefore, one of the key questions for the origin of life is how peptides/proteins are formed from amino acids in the prebiotic environment.
Studies on the formation of probiotic peptides are mainly carried out by simulating the Hadean environment. Forsythe et al. discovered that the ester-mediated amide bond formation was driven by wet-dry cycles [2]. The relative abundance of 66% alanine was obtained by signal intensities of mass spectrometry and theoretical calculations, which is not an absolute conversion ratio of amino acid. Carbonyl sulfide (COS) was one of the key compounds in the hydrothermal area. Greenwald et al. reported that the polymerization of amino acids was continuously activated by COS [3]. According to Greenwald et al., amino acids were converted into peptides and amyloid fibers under the COSactivated process. Rabinowitz and his colleagues discovered the formation of dipeptides in an aqueous solution of amino acids and P3m under alkaline condition [4]. Yamagata et al. demonstrated that volcanic activities produced water-soluble phosphates and polyphosphates including P3m [5]. To date, many efforts have been put into the study on peptide formation under prebiotic conditions [6-12].
It is noted that several groups have reported interesting progress on the study on extraterrestrial life very recently. For example, the icy moons of Jupiter and Saturn presumably nourish chemical gradients at the stratum between crust and ocean, which is similar to the Earth. Therefore, icy moons might be a suitable environment for the process of life evolution [13, 14]. However, Pascal et al. hold a different view that it is difficult to find out the exact origin of life on icy moons merely based on the known conditions [15]. Therefore, evaluating the influences of new conditions on the origin of life on the earth will enlighten the extraterrestrial life exploration.
In 1981, Corliss proposed that life could be originated from the deep-sea hydrothermal vent [16]. Since then, numerous studies on the origin of life were based on this hypothesis [17-20]. In particular, the Lost City hydrothermal vent, where there are alkaline fluids (pH 9–11), might be appropriate as natural electrochemical reactors for driving the origin of life [18, 21]. High hydrostatic pressure, as one of the important environmental factors in hydrothermal vents, exerts profound effect on the physical properties of proteins and some other biomolecules [22-24]. But the effect of high hydrostatic pressure on the formation of prebiotic peptides remains unclear. Meanwhile, most of the submarine hydrothermal vents (e.g., mid-ocean ridge) were found 1–3 kilometers beneath the seafloor [25], where the pressure is up to 100–300 bar.
In this paper, we experimentally explored the properties of peptide formation under high hydrostatic pressures (300 bar). To this purpose, 0.1 mol/L P3m was respectively treated with 0.1 mol/L amino acids (Phe, Trp, Met, Val, Gly and Ala) under different pressure and temperature conditions to study the effect of different pressures on peptide formation.
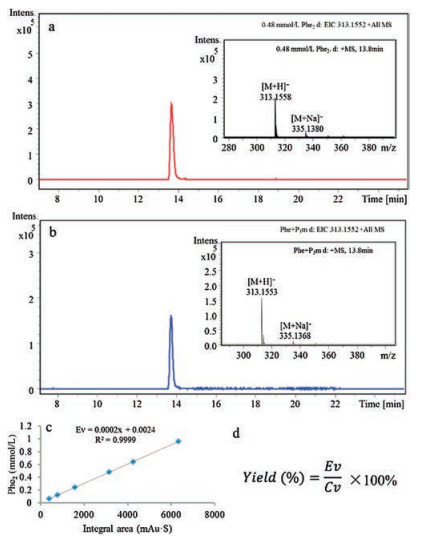
The dipeptide product (Phe2) was analyzed by HPLC-MS and identical to authentic dipeptide Phe2 (Scheme 1 and Fig. 1) [26, 27].

|
Download:
|
| Scheme 1. Reaction of amino acids with P3m for dipeptide synthesis. | |

|
Download:
|
| Fig. 1. Formation of Phe2 from a reaction mixture of Phe and P3m at alkaline aqueous solution (pH 10.7). (a) HPLC-MS spectrum of authentic Phe2 (0.48 mmol/L), (b) HPLC-MS spectrum of the sample, (c) and (d) the standard curve and the formula for quantifying yields of product Phe2. Ev: Experimental value; Cv: Calculated value. | |
In order to test effect of substrate concentration and Mg2+ on the dipeptide formation, reaction mixtures containing varied concentrations of Phe (10, 30, 50, 80 and 100 mmol/L) and 100 mmol/L P3m were incubated, under 1 bar and 300 bar, respectively, at room temperature in a aqueous solution with the pH 10.7 for 7 days. In another set of reactions, 2 mmol/L Mg2+ was added under the identical conditions. The results showed that the yield of the dipeptide Phe2 was increased with the increased concentrations of reactant. However, Mg2+ has no positive effect on the yields of dipeptide. So Mg2+ was not considered in later experiments (Fig. S1 in Supporting information).
Based on the above results, we treated 0.1 mol/L P3m with 0.1 mol/L amino acids of Phe, Trp, Met, Val, Gly and Ala, respectively, at 26℃ under a pressure of 1 bar and 300 bar. The produced dipeptides were confirmed and quantified through comparing with the standards by HPLC-MS (Figs. S2–S12 in Supporting information). The results showed that these amino acids were converted to corresponding dipeptides except for Val (Table 1 and Fig. S13 in Supporting information).
|
|
Table 1 Reaction conditions and dipeptide yields at 26℃ and 37℃ under different pressures. |

{kind=link}
{kind=link}
As shown in Table 1 and Fig. S13, for Phe, Trp and Met at 26℃, the dipeptide yields under 300 bar were higher than those under 1 bar, which were improved to 2.30, 1.45 and 1.33 times, respectively. However, the yields of dipeptide were almost the same for both Gly and Ala under different pressures.
It is worth noting that most of the dipeptide yields were less than 2% except for Gly and Ala (29.58% and 4.69%, respectively). To evaluate the effect of high hydrostatic pressure at higher temperatures, similar reactions were carried out at 37℃. Indeed, as the temperature increased, the dipeptide yields increased for all the five amino acids (Phe, Trp, Met, Gly and Ala). Meanwhile, for Val, the dipeptide Val2 was also detected and quantified by comparing with the Val2 standard using HPLC-MS (Figs. S7 and S8). The dipeptide yields for Trp2 and Met2 under 300 bar are increased about 17% and 10% higher than those under 1 bar, respectively. The dipeptide yields of Phe2, Val2, Gly2 and Ala2 showed no significant differences under these two pressures (Table 1 and Fig. S14 in Supporting information).
Hence, both temperature and pressure as the deep-sea environmental conditions should be considered simultaneously. As mentioned above, the results showed that the yields of dipeptides increase with raising pressure. Therefore, high hydrostatic pressure could be a much favored factor for prebiotic dipeptide formation.
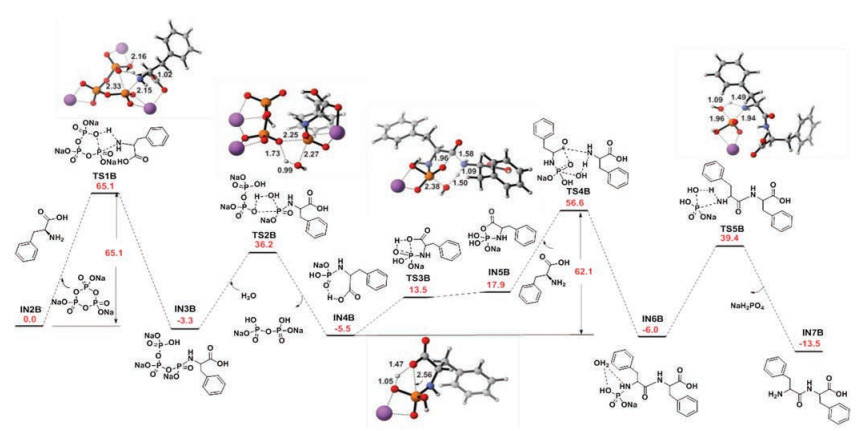
Based on density functional theory calculations at the M06-L/6- 311++G(2d, p)// B3LYP/6-31 G(d) level of theory in the gas phase, free energy profiles of the formation of dipeptides were produced (Figs. 2 and S15 in Supporting information). The rate-determining step of the reactions, Phe2 and Gly2, was found with an extremely high overall barrier of 65.1 kcal/mol and 56.2 kcal/mol, respectively. It suggested that the reactions were thermodynamically and kinetically unfavorable.

|
Download:
|
| Fig. 2. Free energy (kcal/mol) profile of the formation of Phe2. Bond lengths were expressed in terms of angstrom. | |
{kind=link}
These theoretical calculations were consistent with the experimental results above, i.e., the yields of dipeptide were low in the aqueous solution. Moreover, from the free energy profiles of Phe2 and Gly2, it could be found that the energy barriers of all steps of Phe2 formation were lower than that of the corresponding steps of Gly2 formation except for the rate-determining step. Perhaps this explained why the yield of Phe2 under 300 bar was higher than that under 1 bar at 26℃, whereas the yield of Gly2 was almost the same under different pressures. The reason is that the high hydrostatic pressure of 300 bar reduces the energy barrier of ratedetermining step of Phe2, and the energy barriers of other steps are relatively lower, resulting in a significant improvement of the final yield. However, with Gly2 at 300 bar, the energy barriers for other steps are still relatively higher, eventually leading to an insignificant increase in yield. We speculated that when the pressure is increased to a certain extent, the yields of dipeptide, such as Gly2, will also be significantly higher than those under the normal pressure of 1 bar.
The equilibrium constants of aqueous reactions are influenced by both pressure and temperature. The effect of pressure is manifested only when the pressure is great, and such conditions are met in the deep oceans [28]. Meanwhile, Hamed and Owen illustrated the effect of pressure and temperature on the △G (kJ/mol) of water ionization [29]. Therefore, we believed that high pressure increases the equilibrium constant of the reaction. At the same time, it is worth noting that since the pressure in our experiments is not too high, the equilibrium constant has only slightly increased, so that the yield of dipeptide only increases little comparing with that under atmospheric pressure. But it do indicate that the high hydrostatic pressure may serve as one of the key factors in prebiotic peptide synthesis under Hadean deep-sea environment.
In summary, our experimental results showed that high pressure, as a key factor of deep-sea environment conditions, indeed promoted the peptide formation in the reaction system of amino acids and P3m. High pressure could support the hypothesis that life came from the Hadean deep ocean. And it indicates that the influence of pressure should not be neglected while exploring the origin of life. Besides the model of amino acids and P3m, there are a series of ongoing studies on other models of dipeptide formation at high pressures.
AcknowledgmentsThis study was supported by the National Natural Science Foundation of China (Nos. 21778042 and 41576081), and the Fundamental Research Funds for the Central Universities (No. 20720160034). Thanks are also given to Dr. Y. Liu from Xiamen University, Prof. X. Xiao from Shanghai Jiao Tong University, and Dr. X. Tang and P. Huang from the Third Institute of Oceanography of the State Oceanic Administration (China). We are indebted to L. Q. Xu (Ningbo University) for many constructive comments on this manuscript.
Appendix A. Supplementary dataSupplementary material related to this article can be found, in the online version, at doi:https://doi.org/10.1016/j.cclet.2018.06.015.
| [1] |
J.B.S. Haldane, Ration. Annu. 148 (1929) 3-10. |
| [2] |
J.G. Forsythe, S.S. Yu, I. Mamajanov, et al., Angew. Chem. Int. Ed. 54 (2015) 9871-9875. DOI:10.1002/anie.201503792 |
| [3] |
J. Greenwald, M.P. Friedmann, R. Riek, Angew. Chem. Int. Ed. 55 (2016) 11609-11613. DOI:10.1002/anie.201605321 |
| [4] |
J. Rabinowitz, J. Flores, R. Krebsbach, G. Rogers, Nature 224 (1969) 795-796. DOI:10.1038/224795a0 |
| [5] |
Y. Yamagata, H. Watanabe, M. Saitoh, T. Namba, Nature 352 (1991) 516-519. DOI:10.1038/352516a0 |
| [6] |
X. Gao, Y. Liu, P.X. Xu, et al., Amino Acids 34 (2008) 47-53. DOI:10.1007/s00726-007-0599-8 |
| [7] |
A. Hill, L.E. Orgel, Helv. Chim. Acta 85 (2002) 4244-4254. DOI:10.1002/hlca.200290009 |
| [8] |
J. Rabinowitz, Helv. Chim. Acta 53 (1970) 1350-1355. |
| [9] |
J. Rabinowitz, H. Aioub, Helv. Chim. Acta 61 (1978) 1842-1847. |
| [10] |
Z.Z. Chen, Y.F. Tong, S.B. Chen, et al., Sci. Bull. 47 (2002) 1866-1870. DOI:10.1360/02tb9408 |
| [11] |
R. Hu, J. Tian, Y. Yin, Sci. China Chem. 49 (2006) 351-356. DOI:10.1007/s11426-006-2003-z |
| [12] |
W. Jia, Z.S. Zhang, X.W. Yu, et al., Sci. Bull. 56 (2011) 633-639. DOI:10.1007/s11434-010-4314-x |
| [13] |
N.G. Holm, C. Oze, O. Mousis, et al., Astrobiology 15 (2015) 587-600. DOI:10.1089/ast.2014.1188 |
| [14] |
R.T. Pappalardo, S. Vance, F. Bagenal, et al., Astrobiology 13 (2013) 740-773. DOI:10.1089/ast.2013.1003 |
| [15] |
R. Pascal, Astrobiology 16 (2016) 328-334. DOI:10.1089/ast.2015.1412 |
| [16] |
J.B. Corliss, J.A. Baross, S.E. Hoffman, Oceanol. Acta 4 (1981) 59-69. |
| [17] |
C. Huber, G. Wächtershäuser, Science 281 (1998) 670-672. DOI:10.1126/science.281.5377.670 |
| [18] |
D.S. Kelley, J.A. Karson, D.K. Blackman, et al., Nature 412 (2001) 145-149. DOI:10.1038/35084000 |
| [19] |
W. Martin, J. Baross, D. Kelley, M.J. Russell, Nat. Rev. Microbiol. 6 (2008) 805-814. DOI:10.1038/nrmicro1991 |
| [20] |
S.L. Miller, J.L. Bada, Nature 334 (1988) 609-611. DOI:10.1038/334609a0 |
| [21] |
B. Herschy, A. Whicher, E. Camprubi, et al., J. Mol. Evol. 79 (2014) 213-227. DOI:10.1007/s00239-014-9658-4 |
| [22] |
K. Heremans, Annu. Rev. Biophys. Bioeng. 11 (1982) 1-21. DOI:10.1146/annurev.bb.11.060182.000245 |
| [23] |
S. Kapoor, M. Berghaus, S. Suladze, et al., Angew. Chem. Int. Ed. 53 (2014) 8397-8401. DOI:10.1002/anie.v53.32 |
| [24] |
P.C. Michels, D.S. Clark, Appl. Environ. Microbiol. 63 (1997) 3985-3991. |
| [25] |
D.S. Kelley, J.A. Baross, J.R. Delaney, Annu. Rev. Earth Planet. Sci. 30 (2002) 385-491. DOI:10.1146/annurev.earth.30.091201.141331 |
| [26] |
C.M. Cheng, X.H. Liu, Y.M. Li, et al., Orig. Life Evol. Biosph. 34 (2004) 455-464. DOI:10.1023/B:ORIG.0000043122.97856.79 |
| [27] |
F. Ni, C. Fu, X. Gao, et al., Sci. China Chem. 58 (2015) 374-382. DOI:10.1007/s11426-015-5321-1 |
| [28] |
B.B. Owen, S.R. Brinkley, Chem. Rev. 29 (1941) 461-474. DOI:10.1021/cr60094a003 |
| [29] |
H.S. Harned, B.B. Owen, C.V. King, Electrochem. Soc. 106 (1959) 15C. |