 2018, Vol. 29
2018, Vol. 29 



b School of Life Sciences and Technology Center for Protein Sciences, Tsinghua University, Beijing 100084, China
Protein synthesis is a fundamental process for all living organisms, which is carried on the ribosome and assisted by several protein factors, including four well-known translational GTPases (trGTPases). They are initiation factor 2 (IF2), elongation factor Tu (EF-Tu), elongation factor G (EF-G), and release factor 3 (RF3) [1, 2]. In addition, there are a number of noncanonical trGTPases identified in bacteria, whose physiological functions and molecular mechanisms are largely unknown [3].
Elongation factor 4 (EF4, also known as LepA) is one of the poorly-understood noncanonical trGTPases, which is reported as the third most conserved protein in bacteria, falling behind EF-G and EF-Tu [4]. EF4 is also present in the mitochondria and chloroplasts of eukaryotes, but it is absent from archaea and the cytosol of eukaryotes. Despite its highly conservation, absence of EF4 has no obvious effect on growth of bacteria under optimal growing conditions [5]. However, EF4 is required for growth under stress conditions, such as high concentration of magnesium, low pH, and non-optimal temperature [6-8]. Deletion of the lepA gene in Escherichia coli has been shown to cause growth defect in the presence of potassium tellurite or penicillin G [9]. On the other hand, deletion of the lepA gene increases survival rate of E. coli against lethal antimicrobials [10].
The overall structure of EF4 is highly similar to EF-G [11]. However, EF4 has an unique C-terminal domain (CTD), which contains a large number of positively charged residues, and lacks the IV domain and G' subdomain which are present in EF-G. EF4 has been shown to bind with ribosomes in different forms and conformations [12-16]. EF4 bound pre-translocation (PRE) ribosomal complex adopts an unrotated conformation with an intermediate A/L or A/4 state tRNA, which is remodeled from the canonical A/A state through interactions between EF4's CTD and CCA end of the tRNA. In addition, EF4 bound post-translocation (POST) ribosomal complexes have been visualized in unrotated, clockwise rotated, and counterclockwise rotated conformations. EF4's CTD can interact with either the 23S rRNA or P-site tRNA to disrupt base-pairing between the P loop and the P-site tRNA and to remodel the peptidyl transferase center (PTC).
EF4 was firstly named as LepA, because it is the first gene in the bicistronic leader peptidase operon [17]. EF4 locates in the membrane and is released to the cytosol under certain stress conditions [6, 18]. Absent of EF4 has no detectable effect on protein secretion at cellular level [5], which initiated further investigations on cellular functions and molecular mechanisms of EF4. Nierhaus and colleagues first reported that EF4 induces back translocation of the ribosome [4], which was proposed as the molecular function of EF4. However, this model has been challenged by several later discoveries [19-21]. Back translocation in the presence of EF4 takes at least several minutes to complete [22], which is as slow as the spontaneous back translocation in the absence of any elongation factor [23, 24]. In addition, further evidences indicated that EF4 can directly interact with PRE complexes and compete with EF-G to modulate translocation [19]. Furthermore, EF4 has been proposed to function in ribosome biogenesis and the initiation phase of translation [20, 21].
On top of these controversial cellular, structural, and biochemical studies, cellular functions and molecular mechanisms of EF4 have remained elusive. By far, how EF4 interacts with ribosomal PRE and POST complexes and how EF4 modulates elongation rates of translating ribosomes are not fully revealed. In the last two decades, single-molecule techniques have been emerging and become widely used tools to provide new insights into dynamics of biological processes and their molecular mechanisms [25-31]. Here, we established and applied several single-molecule fluorescence resonance energy transfer (smFRET) assays [32, 33] to address these key questions and to shed light on functions and mechanisms of EF4 in elongation cycle.
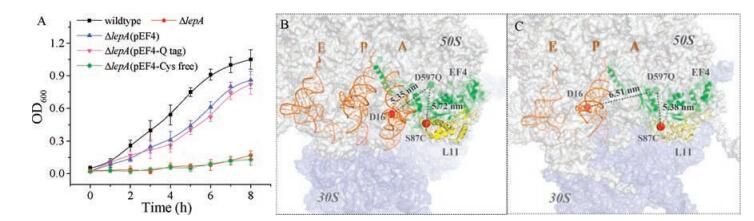
We designed two EF4 variants for site-specific labeling. A Cysfree EF4, whose endogenous cysteine residues were all mutated to other amino acids, and a Q-tag fused variant containing a short peptide (GQQQLG) at its C terminal (Table S1). Q-tag fused protein is one of alternative approaches to achieve site-specific labeling on non-cysteine residues [34-38]. Deletion of lepA gene has been shown to cause a growth defect in the presence of tellurite [9]. Introducing a plasmid encoding wt-EF4 or Q-tag fused EF4 into △lepA E. coli strain rescued its growth defect under tellurite stress, whereas introducing a plasmid carrying Cys-free EF4 failed to rescue growth defect of the △lepA strain (Fig. 1A). The fact that Cysfree EF4 cannot compensate cellular functions of wt-EF4 under tellurite stress suggests that one or several endogenous cysteine residues of EF4 are essential for its functions. Mutation on essential cysteine residues caused loss of functions. Therefore, we did not generate further variants on top of Cys-free EF4 for site-specific labeling. To label Q-tag fused EF4, tetramethyl rhodamine (TMR) was ligated on the third residues of Q-tag (GQQQLG, underlined Q) through site-specific ligation catalyzed by transglutaminase [34]. TMR labeled Q-tag fused EF4 (TMR-EF4) was used for further smFRET measurements (Figs. 1B and C).

|
Download:
|
| Fig. 1. EF4 variants for smFRET. (A) Growth curves of E. coli strains in the presence of 0.2 μg/mL potassium tellurite at 37 ℃. Strain BW25113 (wildtype) and strain JW2553 (△lepA) were obtained from the Keio collection. Plasmids encoding wt-EF4 (pEF4), Q-tag fused EF4 (pEF4-Q tag), and Cys-free EF4 (pEF4-Cys free) were transformed in △lepA strain, respectively. Structures of EF4 bound PRE (B, pdb: 3JCE) and POST (C, pdb: 4W2E) ribosomal complexes displaying fluorophore labeling sites and distances between FRET pairs. EF4, ribosomal protein L11, and tRNAs are shown in green, yellow, and brown, respectively. Ribosomal 50S and 30S subunits were shown in grey and light blue, respectively. | |
Firstly, we used EF4/L11 FRET pair to probe interactions between EF4 and the ribosome. The relative distances between labeling sites on Q-tag fused EF4 and ribosomal protein L11 are similar in EF4 bound PRE complex (5.72 nm, Fig. 1B) and EF4 bound POST complex (5.38 nm, Fig. 1C), which enable us to capture transient interactions between ribosomes and EF4 through smFRET between surface-attached ribosomes containing Cy5 labeled L11 (Cy5-L11) and freely-diffusing TMR-EF4 (Fig. 2A). Ribosomes programmed with mRNA-MF were prepared in three different forms, initiation complexes (IC) with initiator tRNAfMet in the P-site and an empty A-site, PRE complexes with initiator tRNAfMet in the P-site and tRNAPhe in the A-site, and POST complexes with tRNAPhe in the P-site and an empty A-site. Interactions of EF4 with different forms of ribosomes were characterized in the presence of GTP or GDP. From individual FRET trajectories (Fig. 2A), we extracted FRET values of each frame, waiting time before transient EF4 binding on the ribosome (τoff), and dwell time of transient EF4 binding (τon), from which rate of EF4 association on the ribosome kass = 1/([EF4]·τoff), rate of EF4 dissociation kdis = 1/τon, dissociation constant Kd = kdis/kass, and peak values of FRET distributions (E) were calculated (Table 1).

|
Download:
|
| Fig. 2. Interactions between EF4 and the ribosome probed by EF4/L11 FRET. (A) Single-molecule fluorescence and FRET traces captured transient interactions between EF4 and ribosomal PRE complex through FRET pair of TMR-EF4 and Cy5- L11. In addition, similar single-molecule traces were captured while using EF4/L11 FRET pair to probe interactions between EF4 and ribosomal POST or initiation complex. FRET efficiencies and dynamic parameters of transient EF4 binding events were extracted and listed in Table 1. (B) Changes of FRET efficiency during EF4 binding events probed by EF4/L11 FRET pairs in TAM15 buffer (red) or N buffer (grey). Distributions of △E, defined as Elast - Efirst, were plotted, in which Efirst and Elast were FRET efficiencies of the first and last frames of transient FRET events. △E = 0 was indicated by dash line. | |
|
|
Table 1 Parameters of transient EF4-ribosome interactions probed by EF4/L11 FRET. |

{kind=link}
{kind=link}
Dissociation constants between EF4 in its GTP and GDP forms and ribosomes in different forms were in the range of 1–60 μmol/L, which agree with reported dissociation constant between EF4 and 70S [39]. Therefore, fluorophore labeling has no significant effect on binding of EF4 on the ribosome. Association rates of EF4 onto ribosomes in all conditions were similar to or slightly slower than association rates of EF-G·GTP (0.3–3 μmol L-1 s-1) [40] and aatRNA·EF-Tu·GTP (10 μmol L-1 s-1) [33] onto ribosomes captured by smFRET measurements under similar experimental conditions. Dissociation rates of EF4 were also similar to dissociation rates of EF-G and EF-Tu (3–10 s-1) [40-43]. Together, our results indicated both EF4·GTP and EF4·GDP have similar and slightly weaker binding affinities on the ribosome than EF-G·GTP and aa-tRNA·EFTu·GTP, respectively. Furthermore, EF4·GTP has higher binding affinity towards PRE and POST complexes than EF4·GDP, whereas both EF4·GTP and EF4·GDP present similar binding affinity on IC. Among three different forms of the ribosome, EF4 prefers to bind PRE complex in the presence of either GTP or GDP. Similar behaviors were found in TAM15 buffer containing 15 mmol/L Mg2+ and N buffer containing 4.5 mmol/L Mg2+. Both EF4·GDPNP and EF4 H81A·GTP displayed transient binding towards both PRE and POST complexes as EF4·GTP, which implied that GTP hydrolysis has minor effects on dissociation of EF4 from ribosomes. However, GTP hydrolysis is needed for rapid release of EF-G from ribosomes [44].
FRET values between TMR-EF4 and Cy5-L11 were around 0.4- 0.5 under all conditions, which generally agree with the relative distances between two labeling sites. In addition, we captured a minor FRET decrease (△E = -0.1) when EF4·GTP interacted with PRE or POST complexes (Fig. 2B, Table 1), which corresponded to a ~0.3 nm increase of distance between labeling sites caused by relative movements between EF4 and the ribosome. In TAM15 buffer, such FRET decrease fully depended on GTP hydrolysis, whereas in N buffer, FRET decrease only partially depended on GTP hydrolysis. Therefore, buffer conditions, including concentrations of Mg2+ and polyamine, modulate conformational structures and dynamics of EF4 bound ribosomes.
Next, we captured smFRET between TMR-EF4 and Cy5 labeled tRNAPhe in the A-site of PRE complex or between TMR-EF4 and Cy5 labeled tRNAPhe in the P-site of POSTcomplex when EF4 transiently interacts with the ribosome (Fig. S1, Table S2 in Supporting information). FRET values in the presence of GDP were lower than the ones in the presence of GTP and highly depended on buffer conditions. In TAM15 buffer, FRET values between EF4·GDP and tRNA were significantly lower, which suggested that EF4 and labeled tRNA were further separated under such buffer environment. On the other hand, FRET values between EF4·GTP and tRNA were all around 0.35-0.42 for both PRE and POSTcomplexes in both buffers. However, in the existing structures, the relative distance between labeling sites on Q-tag fused EF4 and the A-site tRNA in PRE complex (5.35 nm, Fig. 1B) is shorter than the distance between labeling sites on EF4 and the P-site tRNA in POST complex (6.51 nm, Fig. 1C). Previous simulation has suggested that the CTD domain of EF4 is high flexible [13]. Therefore, EF4·GTP's CTD domain, including the labeling site, is likely to move closer to the Psite tRNA than what have been shown in the existing EF4 bound POST complexes. A minor FRET decrease (△E = ~0.05–0.09) was also captured via EF4/tRNA FRET pair with either PRE or POST complexes, which was mainly independent of GTP hydrolysis (Fig. S1B, Table S2).
Both ribosomal PRE and POST complexes have been shown to spontaneously fluctuate between two major conformational states, a non-ratcheted state with tRNAs in classical position and a ratcheted state with tRNAs in hybrid position [32, 45-47]. Based on our previous established smFRET assays [32, 33], we examined how EF4 and its H81A variant, which cannot hydrolyze GTP, affect conformational dynamics between these two states for ribosomal POST and PRE complexes through smFRET between L1 and the P-site tRNA and between L11 and the A-site tRNA, respectively (Figs. 3A and B). Transition rates between two conformational states and their FRET values were extracted from our results (Figs. 3C and D, Table S3 in Supporting information).

|
Download:
|
| Fig. 3. Conformational dynamics of the ribosome modulated by EF4. Cartoon demonstrating how to use L1/tRNA and L11/tRNA FRET pair to capture structural dynamics of ribosomal POST (A) and PRE (B) complexes, respectively. In both cases, ribosomes spontaneously fluctuate between a non-ratcheted state and a ratcheted state. (C) Transition rates between two conformational states under different conditions. (D) FRET distributions of non-ratcheted PRE complexes captured by L11/tRNA FRET in TAM15 buffer (red) or N buffer (grey). | |
{kind=link}
For POST complex, the presence of EF4·GTP shifted the equilibrium by stabilizing ratcheted state and destabilizing nonratcheted state. EF4·GDPNP, EF4·GDP, and EF4-H81A·GTP caused similar but smaller changes on equilibrium of ribosomal conformation than EF4·GTP. Similar results were found in both TAM15 buffer and N buffer. On the other hand, EF4 in GTP, GDPNP, and GDP forms and EF4-H81A in GTP form only caused minor changes on equilibrium of the PRE complex. However, FRET values of nonratcheted PRE state probed by L11/tRNA FRET pair increased from 0.68-0.69 in the absence of EF4 to 0.83-0.87 in the presence of EF4·GDPNP or EF4-H81A·GTP with TAM15 buffer and in the presence of all forms of EF4 with N buffer, which indicated that the A-site tRNA and L11 got closer under these conditions. Our results agree well with previous structures of EF4 bound PRE complexes, in which EF4 remodels the A-site tRNA to move towards L11 to form an A/L or A/4 state [12-14].
Lastly, we employed smFRET measurements between Cy5 labeled ribosomal protein L1 and Cy3 labeled Phe-tRNAPhe to examine how rates of aa-tRNA accommodation and translocation are modulated by EF4 (Fig. 4). Ribosomes programmed with mRNA-MF and mRNA-MVF were used to characterize influence of EF4 on elongation rates in the 1st and 2nd elongation cycles, respectively. Clearly, EF4·GTP severely hindered delivery of aatRNA and moderately increased translocation rates under all conditions we tested (Fig. 4). In addition, EF4-H81A·GTP hindered both aa-tRNA delivery and translocation, which indicated that GTP hydrolysis is essential for EF4 to facilitate translocation.

|
Download:
|
| Fig. 4. Elongation rates modulated by EF4. (A) Single-molecule fluorescence and FRET traces captured tRNA delivery and translocation through L1/tRNA FRET pair. Their dwell times were extracted from individual single-molecule traces and plotted as distributions in Figs. S2 and S3 in Supporting information, from which their average dwell times and apparent rates were fitted by single exponential decay. Normalized tRNA delivery rates (B) and elongation rates (C) under different conditions. | |
{kind=link}
Previous structural studies have captured EF4 bound ribosomes in PRE-translocation and POST-translocation states [12-16]. Here, using smFRET assays, we revealed that EF4·GTP directly interacts both PRE and POST complexes with similar affinities (Table 1). Similar behaviors have been found when examining interactions between EF-G and the ribosome. Several structural and singlemolecule studies have shown that EF-G can interact with both PRE and POST complexes, as well as several intermediate states along translation pathway [32, 41, 48-57]. Together, these studies emphasized that, EF4 and EF-G, two elongation factors sharing structural similarity, are flexible to bind with ribosomes in different forms and conformations.
All three published structures of EF4 bound ribosomal PRE complexes, which were prepared using non-hydrolyzable GTP analogue (GDPNP or GDPCP), contained a remodeled A/L or A/4 tRNA, which originally located at the canonical A-site. Through L11/tRNA FRET pair, we directly captured remodeling of the A-site tRNA in the presence of either EF4·GDPNP or EF4-H81A·GTP (Fig. 3D and Table S3 in Supporting information). However, in TAM15 buffer, FRET value of L11/tRNA pair was almost unaffected by the presence of EF4·GTP, which indicated that tRNA mainly remains in the canonical A-site under this condition. In addition, 4 mmol/L of EF4·GTP was used here, which is higher than binding affinity of EF4·GTP on PRE complexes, to ensure that majority of PRE complexes were bound by EF4 while capturing L11/tRNA FRET. Therefore, we suspected that, in TAM15 buffer, our smFRET measurements captured a new transient EF4·GTP bound ribosomal PRE complex containing the canonical A-site tRNA. Such complex might be short-lived and unstable, which is hard to be captured by structural methods.
So far, there are four structures of EF4 bound ribosomal POST complexes prepared in the presence of GDP, GDPNP, or GDPCP. Among them, ribosomal 30S subunit has been found in an unrotated, a clockwise-rotated, or a counterclockwise-rotated state relative to the 50S subunit. The unrotated and counterclockwise-rotated states are also known as the non-ratcheted and ratcheted states, respectively. Our results indicated that interactions between EF4 and POST complexes stabilize the ratcheted state. In addition, stabilization of the ratcheted state is mostly independent of GTP hydrolysis, becauseEF4 inGTP, GDPNP, and GDP formsandEF4-H81A in GTP form all shifted equilibrium towards the ratcheted state. Furthermore, FRETpairbetweenEF4·GTPandtheP-sitetRNAinPOST complex produced similar FRET values (E = 0.35–0.42) as FRET pair between EF4·GTP and the A-site tRNA in PRE complex (E = 0.36- 0.42). Our observations cannot be rationalized by current published structures (Figs. 1B and C). Because the distance between labeling sites on EF4 and tRNA in POSTcomplex was significantly longer than the one in PRE complex, which was expected to cause a significant difference between their FRET values (△E = ~0.1). Together, the EF4·GTP bound ribosomal POST complex captured by our smFRET measurements is likely to represent another new unknown structure, in which EF4 moves closer to the P-site tRNA than what was shown in current structures.
In the elongation phase of translation, ribosomes cycle between PRE and POST complexes. EF-Tu interacts with POST complex to delivery aa-tRNA to ribosomal A-site, which leads to the formation of PRE complex. EF-G interacts with PRE complex to promote translocation to form POST complex. EF4 may compete with EF-Tu and EF-G to interact with ribosomes, because they share the same ribosomal binding sites and have similar binding affinities towards ribosomes. Previous biochemical studies have shown that EF4 competes with EF-G to interact with PRE complex and to promote transition from PRE complex to a translocation intermediate state [19]. Here, we found that EF4 can moderately assist translocation under our experimental conditions. EF4-H81A, a variant cannot hydrolyze GTP, interacts with PRE complex and severely hinders translocation. In addition, both EF4·GTP and EF4-H81A·GTP stabilize POST complex in its ratcheted state, which is not the correct ribosomal conformational state for aa-tRNA·EF-Tu·GTP to bind and to delivery aa-tRNA [58]. As a result, the presence of EF4·GTP or EF4-H81A·GTP significantly slows delivery rate of aatRNA, which is in line with previous observations that EF4 inhibits delivery of tmRNA·EF-Tu·GTP to the ribosomal A-site [9]. In all, EF4 modulates elongation through its interactions with both PRE and POST complexes.
Several working mechanisms of EF4 have been proposed, including inducing back-translocation, sequestering translating ribosomes, unfolding and refolding a misfolded nascent peptide chain, and contributing to ribosome biogenesis [59]. Here, we mainly focus on elucidating molecular mechanisms of EF4 in elongation cycle. Our smFRET measurements showed that EF4 only transiently interacts with ribosomes for 0.5 s or less. However, EF4 induced back-translocation takes several minutes or longer to complete [22], whose rate is similar to the rate of spontaneous back-translocation in the absence of any elongation factor [23, 24]. Furthermore, we revealed that EF4 stabilizes the POST complex in the ratcheted state, in which the P-site tRNA moves to the hybrid P/E site, an opposite direction of back-translocation. Therefore, we have no evidence to support EF4 to function as a back-translocase.
GTP hydrolysis is essential for EF4's cellular functions. Results from ribosome profiling [20] indicated that, at cellular level, EF4- H81A cannot rescue phenotypes caused by deletion of EF4/LepA gene. The only major difference we found between EF4·GTP and EF4-H81A·GTP is that EF4·GTP moderately accelerates translocation whereas EF4-H81A·GTP slows translocation by 2 folds. On the other hand, both EF4·GTP and EF4-H81A·GTP severely hinder aatRNA delivery. Therefore, we suspected that molecular functions of EF4·GTP in elongation are to slow overall elongation rate and to stall majority of translating ribosomes in their POST states.
We proposed the following model to reconcile mechanisms and functions of EF4 during elongation (Fig. 5). Stress conditions, such as high concentration of magnesium, low pH, and non-optimal temperature might cause misfolding of nascent peptides and malfunction of translating ribosomes and elongation factors. Under these conditions, EF4 is released from membrane to the cytosol to interact with translating ribosomes. On one hand, EF4 interacts with ribosomal PRE complexes, probably including dysfunctional PRE complexes, to resume and promote translocation to form POST complexes. On the other hand, EF4 interacts with ribosomal POST complexes to slow aa-tRNA delivery, so that majority of ribosomes stay in POST states. As a result, overall elongation rate is slowed down and nascent peptides have more times to fold correctly. In addition, interactions between EF4 and tRNA in POST complexes could protect nascent peptide on the Psite tRNA from hydrolysis [16], so that normal elongation can be resumed once stress is released. If stress persists, stalled POST complexes might be recycled by RelE, tmRNA, or other factors mediated rescue pathways [60-62]. In conclusion, EF4 interacts with both PRE and POST ribosomal complexes and modulates substeps of elongation in different manners to fulfill its functions.

|
Download:
|
| Fig. 5. A proposed reaction scheme to illustrate working mechanisms and functions of EF4. EF4 interacts with both PRE and POST complexes. It slows down overall elongation rates and mainly stalls elongating ribosomes in their POST states by inhibiting delivery of aa-tRNA. Interactions between EF4 and the P-site tRNA in POST complexes could protect nascent peptide from hydrolysis, so that normal elongation can be resumed once stress is released. If stress persists, stalled POST complexes might be recycled by rescue pathways. | |
{kind=link}
Acknowledgments
This project was supported by funds from the National Natural Science Foundation of China (No. 31570754), Tsinghua-Peking Joint Center for Life Sciences and Beijing Advanced Innovation Center for Structural Biology to C. Chen and Lab Innovation Funding from Lab and Instrument Department, Tsinghua University to W. Wang.
Appendix A. Supplementary dataSupplementary data associated with this article can be found, in the online version, at https://doi.org/10.1016/j.cclet.2017.12.006.
| [1] |
T.M. Schmeing, V. Ramakrishnan, Nature 461 (2009) 1234-1242. DOI:10.1038/nature08403 |
| [2] |
T. Margus, M. Remm, T. Tenson, BMC Genom 8 (2007) 15. DOI:10.1186/1471-2164-8-15 |
| [3] |
R. Ero, V. Kumar, Y. Chen, Y.G. Gao, RNA Biol. 13 (2016) 1258-1273. DOI:10.1080/15476286.2016.1201627 |
| [4] |
Y. Qin, N. Polacek, O. Vesper, et al., Cell 127 (2006) 721-733. DOI:10.1016/j.cell.2006.09.037 |
| [5] |
N.J. Dibb, P.B. Wolfe, J. Bacteriol. 166 (1986) 83-87. DOI:10.1128/jb.166.1.83-87.1986 |
| [6] |
M. Pech, Z. Karim, H. Yamamoto, et al., Proc. Natl. Acad. Sci. U. S. A. 108 (2011) 3199-3203. DOI:10.1073/pnas.1012994108 |
| [7] |
F. Yang, Y. Gao, Z. Li, et al., Biochim. Biophys. Acta 1837 (2014) 1674-1683. DOI:10.1016/j.bbabio.2014.05.353 |
| [8] |
F. Yang, Z. Li, J. Hao, Y. Qin, Protein Cell 5 (2014) 563-567. DOI:10.1007/s13238-014-0050-3 |
| [9] |
S. Shoji, B.D. Janssen, C.S. Hayes, K. Fredrick, Biochimie 92 (2010) 157-163. DOI:10.1016/j.biochi.2009.11.002 |
| [10] |
L. Li, Y. Hong, G. Luan, et al., mBio 5 (2014) e01708. |
| [11] |
R.N. Evans, G. Blaha, S. Bailey, T.A. Steitz, Proc. Natl. Acad. Sci. U. S. A. 105 (2008) 4673-4678. DOI:10.1073/pnas.0801308105 |
| [12] |
S.R. Connell, M. Topf, Y. Qin, et al., Nat. Struct. Mol. Biol. 15 (2008) 910-915. DOI:10.1038/nsmb.1469 |
| [13] |
D. Zhang, K. Yan, G. Liu, et al., Nat. Struct. Mol. Biol. 23 (2016) 125-131. DOI:10.1038/nsmb.3160 |
| [14] |
M.G. Gagnon, J. Lin, T.A. Steitz, Proc. Natl. Acad. Sci. U. S. A. 113 (2016) 4994-4999. DOI:10.1073/pnas.1522932113 |
| [15] |
M.G. Gagnon, J. Lin, D. Bulkley, T.A. Steitz, Science 345 (2014) 684-687. DOI:10.1126/science.1253525 |
| [16] |
V. Kumar, R. Ero, T. Ahmed, et al., J. Biol. Chem. 291 (2016) 12943-12950. DOI:10.1074/jbc.M116.725945 |
| [17] |
P.E. March, M. Inouye, J. Biol. Chem. 260 (1985) 7206-7213. |
| [18] |
P.E. March, M. Inouye, Proc. Natl. Acad. Sci. U. S. A. 82 (1985) 7500-7504. DOI:10.1073/pnas.82.22.7500 |
| [19] |
H.Q. Liu, C.L. Chen, H.B. Zhang, et al., Proc. Natl. Acad. Sci. U. S. A. 108 (2011) 16223-16228. DOI:10.1073/pnas.1103820108 |
| [20] |
R. Balakrishnan, K. Oman, S. Shoji, R. Bundschuh, K. Fredrick, Nucleic Acids Res. 42 (2014) 13370-13383. DOI:10.1093/nar/gku1098 |
| [21] |
M.R. Gibbs, K.M. Moon, M. Chen, et al., Proc. Natl. Acad. Sci. U. S. A. 114 (2017) 980-985. DOI:10.1073/pnas.1613665114 |
| [22] |
H.Q. Liu, D.L. Pan, M. Pech, B.S. Cooperman, J. Mol. Biol. 396 (2010) 1043-1052. DOI:10.1016/j.jmb.2009.12.043 |
| [23] |
S. Shoji, S.E. Walker, K. Fredrick, Mol. Cell. 24 (2006) 931-942. DOI:10.1016/j.molcel.2006.11.025 |
| [24] |
A.L. Konevega, N. Fischer, Y.P. Semenkov, et al., Nat. Struct. Mol. Biol. 14 (2007) 318-324. DOI:10.1038/nsmb1221 |
| [25] |
S.K. Chakkarapani, G. Park, S.H. Kang, Chin. Chem. Lett. 26 (2015) 1490-1495. DOI:10.1016/j.cclet.2015.10.017 |
| [26] |
van Oijen A.M., N.E. Dixon, Nat. Struct. Mol. Biol. 22 (2015) 948-952. DOI:10.1038/nsmb.3119 |
| [27] |
C.J. Bustamante, C.M. Kaiser, R.A. Maillard, D.H. Goldman, C.A. Wilson, Annu. Rev. Biophys. 43 (2014) 119-140. DOI:10.1146/annurev-biophys-051013-022811 |
| [28] |
C.E. Aitken, A. Petrov, J.D. Puglisi, Annu. Rev. Biophys. 39 (2010) 491-513. DOI:10.1146/annurev.biophys.093008.131427 |
| [29] |
G. Park, S.K. Chakkarapani, S. Ju, S. Ahn, S.H. Kang, Chin. Chem. Lett. 29 (2018) 505-508. DOI:10.1016/j.cclet.2017.08.018 |
| [30] |
F.F. Li, J.N. Gu, X.C. Zhou, Chin. Chem. Lett. 26 (2015) 1514-1517. DOI:10.1016/j.cclet.2015.09.013 |
| [31] |
H. Cai, C. Zhou, Q. Yang, et al., Chin. Chem. Lett. 29 (2018) 531-534. DOI:10.1016/j.cclet.2017.09.010 |
| [32] |
C.L. Chen, B. Stevens, J. Kaur, et al., Mol. Cell. 42 (2011) 367-377. DOI:10.1016/j.molcel.2011.03.024 |
| [33] |
C.L. Chen, H.B. Zhang, S.L. Broitman, et al., Nat. Struct. Mol. Biol. 20 (2013) 582-588. DOI:10.1038/nsmb.2544 |
| [34] |
C.W. Lin, A.Y. Ting, J. Am. Chem. Soc. 128 (2006) 4542-4543. DOI:10.1021/ja0604111 |
| [35] |
J. Yin, A.J. Lin, D.E. Golan, C.T. Walsh, Nat. Protoc. 1 (2006) 280-285. DOI:10.1038/nprot.2006.43 |
| [36] |
S. Leng, Q.L. Qiao, Y. Gao, et al., Chin. Chem. Lett. 28 (2017) 1911-1915. DOI:10.1016/j.cclet.2017.03.034 |
| [37] |
L. Peng, Y. Xu, P. Zou, Chin. Chem. Lett. 28 (2017) 1925-1928. DOI:10.1016/j.cclet.2017.09.037 |
| [38] |
F. Sun, B. W.-Zhang, Chin. Chem. Lett. 28 (2017) 2078-2084. DOI:10.1016/j.cclet.2017.08.052 |
| [39] |
De Laurentiis E.I., H.J. Wieden, Sci. Rep. 5 (2015) 8573. DOI:10.1038/srep08573 |
| [40] |
J. Chen, A. Petrov, A. Tsai, O'Leary S.E., J.D. Puglisi, Nat. Struct. Mol. Biol. 20 (2013) 718-727. DOI:10.1038/nsmb.2567 |
| [41] |
C. Chen, X. Cui, J.F. Beausang, et al., Proc. Natl.Acad. Sci. U. S. A. 113 (2016) 7515-7520. DOI:10.1073/pnas.1602668113 |
| [42] |
T. Pape, W. Wintermeyer, M.V. Rodnina, Embo. J. 17 (1998) 7490-7497. DOI:10.1093/emboj/17.24.7490 |
| [43] |
W. Liu, C. Chen, D. Kavaliauskas, et al., Nucleic Acids Res. 43 (2015) 9519-9528. DOI:10.1093/nar/gkv856 |
| [44] |
M.V. Rodnina, A. Savelsbergh, V.I. Katunin, W. Wintermeyer, Nature 385 (1997) 37-41. DOI:10.1038/385037a0 |
| [45] |
A. Korostelev, D.N. Ermolenko, H.F. Noller, Curr. Opin. Chem. Biol. 12 (2008) 674-683. DOI:10.1016/j.cbpa.2008.08.037 |
| [46] |
J.B. Munro, K.Y. Sanbonmatsu, C.M.T. Spahn, S.C. Blanchard, Trends Biochem. Sci. 34 (2009) 390-400. DOI:10.1016/j.tibs.2009.04.004 |
| [47] |
J. Frank, R.L. Gonzalez, Annu. Rev. Biochem. 79 (2010) 381-412. DOI:10.1146/annurev-biochem-060408-173330 |
| [48] |
N. Fischer, A.L. Konevega, W. Wintermeyer, M.V. Rodnina, H. Stark, Nature 466 (2010) 329-333. DOI:10.1038/nature09206 |
| [49] |
X. Agirrezabala, H.Y. Liao, E. Schreiner, et al., Proc. Natl. Acad. Sci. U. S. A. 109 (2012) 6094-6099. DOI:10.1073/pnas.1201288109 |
| [50] |
D.S. Tourigny, I.S. Fernandez, A.C. Kelley, V. Ramakrishnan, Science 340 (2013) 1235490. DOI:10.1126/science.1235490 |
| [51] |
J. Zhou, L. Lancaster, J.P. Donohue, H.F. Noller, Science 340 (2013) 1236086. DOI:10.1126/science.1236086 |
| [52] |
A. Pulk, J.H.D. Cate, Science 340 (2013) 1235970. DOI:10.1126/science.1235970 |
| [53] |
A.F. Brilot, A.A. Korostelev, D.N. Ermolenko, N. Grigorieff, Proc. Natl. Acad. Sci. U. S. A. 110 (2013) 20994-20999. DOI:10.1073/pnas.1311423110 |
| [54] |
J. Zhou, L. Lancaster, J.P. Donohue, H.F. Noller, Science 345 (2014) 1188-1191. DOI:10.1126/science.1255030 |
| [55] |
E. Salsi, E. Farah, J. Dann, D.N. Ermolenko, Proc.Natl.Acad. Sci. U. S. A. 111 (2014) 15060-15065. DOI:10.1073/pnas.1410873111 |
| [56] |
E. Salsi, E. Farah, Z. Netter, J. Dann, D.N. Ermolenko, J. Mol. Biol. 427 (2015) 454-467. DOI:10.1016/j.jmb.2014.11.010 |
| [57] |
C.L. Chen, B. Stevens, J. Kaur, et al., Proc. Natl. Acad. Sci. U. S. A. 108 (2011) 16980-16985. DOI:10.1073/pnas.1106999108 |
| [58] |
T.M. Schmeing, R.M. Voorhees, A.C. Kelley, et al., Science 326 (2009) 688-694. DOI:10.1126/science.1179700 |
| [59] |
J.L.E. Heller, R. Kamalampeta, H.J. Wieden, Mol. Cell Biol. 37 (2017) e00653-16. |
| [60] |
A.L. Starosta, J. Lassak, K. Jung, D.N. Wilson, FEMS Microbiol. Rev. 38 (2014) 1172-1201. DOI:10.1111/1574-6976.12083 |
| [61] |
E. Giudice, R. Gillet, Trends Biochem. Sci. 38 (2013) 403-411. DOI:10.1016/j.tibs.2013.06.002 |
| [62] |
H. Himeno, N. Nameki, D. Kurita, A. Muto, T. Abo, Biochimie 114 (2015) 102-112. DOI:10.1016/j.biochi.2014.11.014 |