 2016, Vol.27
2016, Vol.27 



b Department of Cell and Systems Biology, University of Toronto, Toronto, ON, Canada M5S 3G5
Juvenile hormone (JH) production in the corpora allata (CA) of Diploptera punctata is tightly controlled and highly predictable during the reproductive cycle. This characteristic provides an ideal model system to study the effect of compounds with anti-JH actions. FGLa-allatostatins (ASTs), a family of pleiotropic neuropeptides, were originally identified for their ability to inhibit JH biosynthesis by CA rapidly and reversibly [1]. Because of their high-efficiency, selectivity and safety to non-target organisms, FGLa-ASTs have been employed in the design of new potential IGRs [2, 3, 4]. Although ASTs inhibit JH biosynthesis effectively in vitro, they have some shortcomings as potential pesticides. First, the more amino acid residues, the higher the production costs. For ASTs, even the shortest AST peptide has six amino acids [5]. Second, ASTs are susceptible to metabolic inactivation by peptidases in hemolymph and midgut [3]. Third, difficulties in ASTs absorption through cuticle could prevent their effects in vivo. Therefore, much effort has been expended to overcome these problems and create better AST peptidomimetics.
Earlier structure-activity relationship (SAR) studies demonstrated that the C-terminal pentapeptide Y/FXFGLa (X = A, N, G, and S) represents the minimum sequence capable of eliciting inhibition of JH production in vitro [6, 7, 8]. Subsequently, based on the peptidomimetic approach and identification of catabolic cleavage sites of ASTs, several AST analogswere discovered to showactivity in vivo as well as reduced susceptibility to metabolic inactivation [2, 3, 4]. As neuropeptides, ASTs exert their effects by binding to a G proteincoupled receptor (GPCR). In 2008, a single FGLa-AstR was isolated and functionally characterized from D. punctata [9]. Recently, the single receptor of D. punctata was activated with thirteen natural FGLa-ASTs [10]. However, whether AST analogs can activate the D. punctata receptor (Dippu-AstR) remains uncertain.
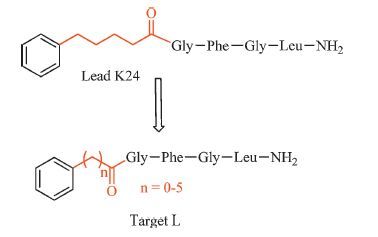
In our previous studies [11, 12, 13, 14, 15], four series of the pentapeptide analogs were designed and synthesized employing the peptidomimetic approach to probe the SAR of the core pentapeptide region of AST. The bioassay and hologram quantitative structure-activity relationships (HQSAR) calculation results suggested that the potent AST analogs should contain an aromatic group, an appropriate length of linker, and an FGLa moiety. Subsequently, a potent AST mimic H17 was found to have a significant inhibitory effect on JH biosynthesis by cockroach CA both in vitro and in vivo [11]. Additionally, owing to its high activity in vitro and its molecular flexibility, K24 was considered as another good lead. In the present work, a series of AST analogs (Fig. 1) were synthesized by modifying the linker of K24 and their bioactivities were evaluated for the purpose of discovering new potential mimics and exploring the effect of linker length on inhibition of JH biosynthesis.

|
Download:
|
| Fig. 1.Design strategy of target analog L. | |
All the solvents and reagents were purchased from commercial suppliers and used without further purification. The analogs structures were confirmed with high-resolution mass spectrometry (HRMS) and 1H NMR. HRMS data was obtained using Agilent Accurate-Mass-Q-TOF MS 6520 system equipped with an Electro Spray Ionization (ESI) source. All the MS experiments were detected in the positive ionization mode. For Q-TOF/MS conditions, fragment and capillary voltages were kept at 130 and 3500 V, respectively. Nitrogen was supplied as the nebulizing and drying gas. The temperature of the drying gas was set at 300 ℃. The flow rate of the drying gas and the pressure of the nebulizer were 10 L/ min and 25 psi, respectively. Full-scan spectra were acquired over a scan range of m/z 80-1200 at 1.03 spectra s-1. 1H NMR spectra was recorded on a Bruker AM-300 (300 MHz) spectrometer with DMSO-d6 as the solvent and TMS as the internal standard. Chemical shifts (δ) are reported in parts per million (ppm).
General synthetic procedure: The synthesis of analogs was illustrated in Scheme 1. GFGL with resin was synthesized from Rink Amide-AM resin (440 mg, 0.3 mmol) using the standard Fmoc/tBu chemistry and HBTU/HOBt protocol [15]. Incoming amino acids were activated with HBTU (456 mg, 1.2 mmol), HOBt (163 mg, 0.3 mmol) and DIEA (210 μL, 0.6 mmol) in DMF (5 mL) for 5 min, couplings were run for 2 h at room temperature. Removal of the Nterminal Fmoc group from the residues was accomplished with 20% piperidine in DMF (5 mL) for 20 min. Relative phenyl acids were coupled to the GFGL with resin with HBTU, HOBt and DIEA in DMF for 3 h at room temperature. Then, analogs were cleaved from the resin with TFA containing 5% phenol, 2.5% thioanisole and 5% water for 2 h at room temperature. The crude peptides were then washed with anhydrous ethyl ether, and lyophilized.

|
Download:
|
| Scheme 1.Synthetic route of lead compound L5 (K24) and target compounds L1–L4, L6. Reagents: (a) 20% piperidine/DMF, 20 min. (b) Fmoc-L-Leu-OH, HBTU, HOBT, DIEA, r.t., 2 h. (c) Fmoc-Gly-OH, HBTU, HOBT, DIEA, r.t., 2 h. (d) Fmoc-L-Phe-OH, HBTU, HOBT, DIEA, r.t., 2 h; (e) Fmoc-Gly-OH, HBTU, HOBT, DIEA, r.t., 2 h; (f) Relative phenyl acids, HBTU, HOBT, DIEA, r.t., 3 h; (g) 20% piperidine/DMF, 20 min; then 90% TFA, 5% phenol, 2.5% H2O, 2.5% TIS, r.t., 2 h. | |
The crude compounds were purified on a C18 reversed-phase preparative column (250 mm × 4.6 mm, 10 μm) with a flow rate of 10 mL/min using the ratio of acetonitrile/water (v/v) 50:50 containing 0.1% TFA. UV detection was at 215 nm. Purification by reversed phase high performance liquid chromatography (HPLC) yielded peptides with over 95% purity. The physical and identification data of target compounds L1-L6 are given in the Supporting information.
2.2. BioassaysThe detailed in vitro and in vivo biological evaluations and functional assays of Dippu-AstR in response to AST analogs are also shown in the Supporting information.
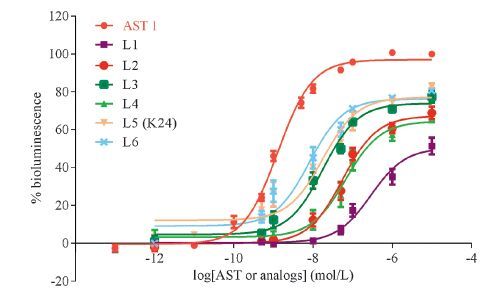
3. Results and discussion 3.1. In vitro effect of AST analogs on the inhibition of JH biosynthesisThe ability of target analogs to inhibit JH biosynthesis was evaluated in vitro using the CA of the cockroach D. punctata. As shown in Table 1, all target analogs exhibit differing potencies as a consequence of varying linker length. L1 (n = 0) shows the lowest activity with an IC50 of 957 nmol/L, compared to other analogs. L2 to L6 are more active in inhibiting JH biosynthesis with IC50 values in the low nanomolar range. L2 (n = 1) with IC50 of 93.5 nmol/L, is about 10-fold activity greater than that of L1. L2, L3 (n = 2) and L4 (n = 3) exhibit slightly improved activity by a small margin when their linker lengths increase from 1-carbon to 3-carbon. L6 (n = 5) with an IC50 of 27.2 nmol/L displays better activity than the lead compound K24/L5 (n = 4, IC50: 37.8 nmol/L). This result suggests that the potency of the analogs may be related to their lipophilicity. Generally, the lipophilicity can be characterized by the retention time or be calculated by the prediction system. Thus, the retention time and log P value of six AST analogs were further determined (Table 1). The result showed that the longer linker length, the longer retention time or the bigger log P value. This demonstrates that longer linkers provide a positive contribution to the inhibitory activity.
|
|
Table 1 Inhibitory effect on JH release (IC50) in vitro, potency in activation of Dippu-AstR (EC50), retention time and log P calculation of AST analogs. |

{kind=link}
{kind=link}
To study the mechanism of action of AST analogs in inhibiting JH biosynthesis, the activity of AST analogs in activation of Dippu-AstR was determined. AST 1, the shortest peptide of the ASTs, was chosen as a control [5]. As shown in Fig. 2, all tested AST analogs induced clear dose-dependent bioluminescence response in Dippu-AstR expressing cells, albeit with different potencies as a consequence of linker length. None of the AST analogs functioned as a full agonist in this cell line. CHO-WTA11 cells transfected with pcDNA3.1D (empty vector) did not show any response to the AST analogs. The EC50 values of AST analogs in activating Dippu-AstR ranged from 62 nmol/L to 2952 nmol/L (Table 1). In general, the ability of AST analogs to activate Dippu- AstR corresponded to their potencies as inhibitors of JH production by CA. This Dippu-AstR activity-linker length relationship was consistent with above relationship between JH inhibitory activity and linker length. However, the AST analogs K24/L5 and L6 which showed similar activity in inhibiting JH biosynthesis as AST 1, exhibited much lower EC50 values in activating the Dippu-AstR (Table 1). This suggests that L5 and L6 exhibit more resistant to metabolic inactivation.

|
Download:
|
| Fig. 2.Dose-response curves for ASTs in CHO-WTA11 cells expressing Dippu-AstR. Data points represent the average ± SEM of three independent measurements performed in duplicate and are expressed as percentage of the maximal response. The zero response level corresponds to treatment with BSA buffer only. | |
{kind=link}
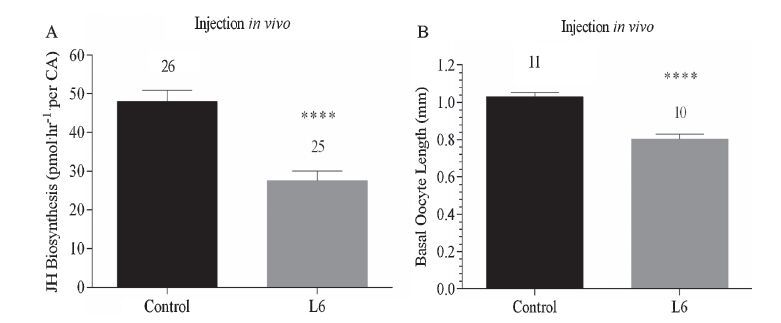
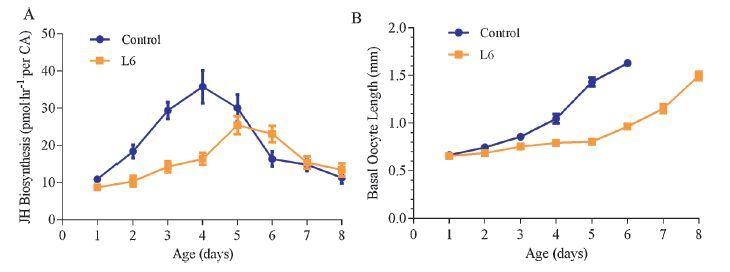
L6, which displays the highest activity in inhibition of JH biosynthesis in vitro, was used to determine its in vivo effect following injection and topical treatment in D. punctata. As shown in Fig. 3A, injection of L6 resulted in an apparent decrease in JH biosynthesis of 42.6% in comparison to control values. Moreover, fluctuation in rates of JH biosynthesis after emergence from day 1 to day 8 is shown in Fig. 4A. L6 exhibited a highly significant and sustained effect on JH biosynthesis following topical application, although the inhibitory effect was reduced on day 6. This indicates that appropriate optimizations in AST structure can increase their resistance to degradation in vivo. Simultaneously, the inhibitory effect of L6 on basal oocyte growth in adult female D. punctata was also assessed. For injected animals, the average oocyte length was about 0.25 mm less than that of untreated animals on day 3 (Fig. 3B). For topical assays, oocytes of treated animals developed more slowly than in control animals (Fig. 4B). Usually untreated females oviposit on day 7 of adult life. However, no treated females had oviposited by day 8.

|
Download:
|
| Fig. 3.Effect of L6 on JH biosynthesis by CA (A) and oocyte growth (B) from day 3 mated female D. punctata in vivo following injection. Five microliter of L6 (final concentration was 10 μmol/L) was injected into cockroaches on day 1 and assayed on day 3. Animals injected with 5 μL of H2O were used as control group. Each bar represents the mean ± SEM for the number of individual measurements indicated at the top of error bars. Asterisks indicate significant differences between peptide- and water-injected groups of animals as determined by t-test: **** P < 0.0001. | |
{kind=link}

|
Download:
|
| Fig. 4.Effect f L6 on JH biosynthesis by CA (A) and basal oocyte growth (B) from day 1 to day 8 in mated female D. punctata following topical application in vivo. Animals were topically treated with 5 μL of L6 in 20% DMSO and 80% acetone on day 0. Animals were topically treated with 5 μL solvent of 20% DMSO and 80% acetone without AST analogs were used as control group. (A) Each point represents mean ± SEM, N = 10–15. (B) Each point represents mean ± SEM, N = 6–8. | |
{kind=link}
Natural ASTs exert little or no effect in vivo because of their rapid degradation by peptidases and poor absorption through the insect cuticle in the case of topical application [16, 17]. To date, only H17 was found to have a significant effect on JH biosynthesis by cockroach CA, both in vitro and in vivo. In the present study, L6, has been shown to represent another potential AST analog with potential to act both in vitro and in vivo.
4. ConclusionIn summary, a series of AST analogs were synthesized by modifying the linker of K24 and their bioactivities were evaluated both in vitro and in vivo. These results show that all target analogs exhibited differing potencies in inhibiting JH biosynthesis and activating Dippu-AstR as a consequence of varying linker length. In particular, analog L6, with the longest linker (n = 5), had very good effect on inhibition of JH biosynthesis both in vitro and in vivo. Furthermore, it displayed good activity in inhibiting basal oocyte growth. These results suggest that L6 is a new potential IGR for cockroach control. Structure-activity relationship suggested that longer linkers increased their ability to inhibit JH biosynthesis.
AcknowledgmentsWeare very grateful to Prof. Jozef Vanden Broeck for his kind help on functional assay of AstR in response of AST analogs. This study was financially supported by the National Natural Science Foundation of China (No. 21372257), the National Basic Research Program of China (973 Program, No. 2010CB126104) and theNatural Sciences and Engineering Research Council of Canada. The author Xiao-qing Wuwas financially supported by China Scholarship Council (CSC) to study in the laboratory of SST at University of Toronto, Canada.
Appendix A. Supplementary dataSupplementary data associated with this article can be found, in the online version, at http://dx.doi.org/10.1016/j.cclet.2016.02.010.
| [1] | W.G. Bendena, B.C. Donly, S.S. Tobe, Allatostatins:a growing family of neuropeptideswith structural and functional diversity, Ann. N.Y. Acad. Sci. 897(1999) 311-329. |
| [2] | M.D. Piulachs, L. Vilaplana, J.M. Bartolome, et al., Ketomethylene and methyleneamino pseudopeptide analogues of insect allatostatins inhibit juvenile hormone and vitellogenin production in the cockroach Blattella germanica, Insect Biochem. Mol. Biol. 27(1997) 851-858. |
| [3] | R.J. Nachman, C.S. Garside, S.S. Tobe, Hemolymph and tissue-bound peptidaseresistant analogs of the insect allatostatins, Peptides (N.Y.) 20(1999) 23-29. |
| [4] | R.J. Nachman, G. Moyna, H.J. Williams, et al., Synthesis, biological activity, and conformational studies of insect allatostatin neuropeptide analogues incorporating turn-promoting moieties, Bioorg. Med. Chem. 6(1998) 1379-1388. |
| [5] | S.S. Tobe, J.R. Zhang, P.R.F. Bowser, et al., Biological activities of the allatostatin family of peptides in the cockroach, Diploptera punctata, and potential interactions with receptors, J. Insect Physiol. 46(2000) 231-242. |
| [6] | T.K. Hayes, X.-C. Guan, V. Johnson, et al., Structure-activity studies of allatostatin 4 on the inhibition of juvenile hormone biosynthesis by corpora allata:the importance of individual side chains and stereochemistry, Peptides (Tarrytown, N.Y.) 15(1994) 1165-1171. |
| [7] | G.E. Pratt, D.E. Farnsworth, K.F. Fok, et al., Identity of a second type of allatostatin from cockroach brains:an octadecapeptide amide with a tyrosine-rich address sequence, Proc. Natl. Acad. Sci. U.S.A. 88(1991) 2412-2416. |
| [8] | G.E. Pratt, D.E. Farnsworth, N.R. Siegel, et al., Identification of an allatostatin from adult Diploptera punctata, Biochem. Biophys. Res. Commun. 163(1989) 1243-1247. |
| [9] | P. Lungchukiet, B.C. Donly, J. Zhang, et al., Molecular cloning and characterization of an allatostatin-like receptor in the cockroach Diploptera punctata, Peptides (Amsterdam, Neth.) 29(2008) 276-285. |
| [10] | J. Huang, E. Marchal, E.F. Hult, et al., Mode of action of allatostatins in the regulation of juvenile hormone biosynthesis in the cockroach, Diploptera punctata, Insect Biochem. Mol. Biol. 54(2014) 61-68. |
| [11] | Z.P. Kai, J. Huang, S.S. Tobe, et al., A potential insect growth regulator:synthesis and bioactivity of an allatostatin mimic, Peptides 30(2009) 1249-1253. |
| [12] | Z.P. Kai, J. Huang, Y. Xie, et al., Synthesis, biological activity, and hologram quantitative structure-activity relationships of novel allatostatin analogues, J. Agric. Food Chem. 58(2010) 2652-2658. |
| [13] | Z.P. Kai, Y. Xie, J. Huang, et al., Peptidomimetics in the discovery of new insect growth regulators:studies on the structure-activity relationships of the core pentapeptide region of allatostatins, J. Agric. Food Chem. 59(2011) 2478-2485. |
| [14] | Y. Xie, Z.P. Kai, S.S. Tobe, et al., Design, synthesis and biological activity of peptidomimetic analogs of insect allatostatins, Peptides 32(2011) 581-586. |
| [15] | Y. Xie, L. Zhang, X.Q. Wu, et al., Probing the active conformation of FGLamide allatostatin analogs with N-terminal modifications using NMR spectroscopy and molecular modeling, Peptides 68(2015) 214-218. |
| [16] | G. Gaede, G.J. Goldsworthy, Insect peptide hormones:a selective review of their physiology and potential application for pest control, Pest Manage. Sci. 59(2003) 1063-1075. |
| [17] | C.S. Garside, R.J. Nachman, S.S. Tobe, Injection of Dip-allatostatin or Dip-allatostatin pseudopeptides into mated female Diploptera punctata inhibits endogenous rates of JH biosynthesis and basal oocyte growth, Insect Biochem. Mol. Biol. 30(2000) 703-710. |