 2016, Vol.27
2016, Vol.27 



Angiotensin Ⅱ (Ang Ⅱ) occupies a vital role in renin-angiotensin system (RAS), which acts as an endocrine system and plays a primary role in renal physiology and cardiovascular [1]. The overactivation of RAS would bring out the progression of atherosclerosis, cardiac disease, diabetes, hypertension and renovascular disorder [2, 3, 4, 5]. Furthermore, Ang Ⅱ, an essential mediator in regulation of blood pressure, could promote oxidative stress via increasing the reactive oxygen species (ROS) generation by interaction with Ang Ⅱ receptor-1 (AT1) to activate nicotinamide adenine dinucleotide phosphate-oxidase (NADPH oxidase) that is an essential source of ROS production in vascular cells [6]. At physiological low concentration, ROS is one kind of important second messenger in intracellular signaling and regulation [7, 8]. While, high level ROS can induce intracellular oxidative stress resulting in inhibition of protein function and DNA damage [9, 10]. Oxidative stress induced by Ang Ⅱ is related to a significant cause of inflammation and cardiovascular disease. Besides, the overactivation of Ang Ⅱ can lead to the expression of monocyte chemoattractant protein-1 (MCP-1), vascular cell adhesion molecule- 1 (VCAM-1) and lectin-like oxidized low-density lipoprotein receptor-1 (LOX-1) which were reported to critically be involved in inflammation and ROS generation in vascular cells as well [11, 12, 13].
Dihydropyridine derivatives (DHPs) can not only regulate the influx of calcium ions but also possess the vasoprotective effects response to cardiovascular events, as DHP can depress the production of ROS effectively induced by Ang Ⅱ through inhibition of the activity of NADPH oxidase [14, 15]. As a result, DHP holds the potential to regulate the intracellular oxidative stress and decrease the proliferation of inflammation, and may be employed as a powerful tool to prevent the progression of heart disease [16, 17, 18, 19].
Conjugated polymers (CPs), formed by a large number of chromic repeat units, exhibit significant optical properties and the ability of easy modification, and provide an excellent platform for imaging and detection [20, 21, 22, 23, 24, 25, 26]. Besides, CPs can harvest the energy of light efficiently and sensitize the surrounding oxygen molecules to generate ROS, thus, CPs are employed to inhibit the proliferation of cancer cells and used as antimicrobial [27, 28, 29]. Polythiophene modified with dihydropyridine (PTDHP) was reported owning ROS-scavenging ability via oxidizing dihydropyridine into pyridine [30]. In this work, we utilize PTDHP to realize the control of intracellular oxidative stress induced by Ang Ⅱ stimulation and the light regulation through ROS generated from polythiophene.
2. Experimental 2.1. Ang Ⅱ incubation time experimentRat aortic endothelial cells were obtained from Pricells (Wuhan, China), and cultured in medium special for primary cells. After reaching 80% confluence, cells were incubated with conditioned medium at 37 ℃ for 24 h before the experiment for inducing quiescence. Then new conditioned medium was exchanged for next steps. At the beginning of the experiment, rat aortic endothelial cells were incubated with Ang Ⅱ (1 × 10-7 mol/L) for 0, 3, 6, 8, 10 h, respectively, in order to induce the production of ROS. As the positive control, cells were incubated with H2O2 (1 × 10-4 mol/L) only. Dihydroethidium (DHE) (5 × 10-6 mol/L) was applied to determinate the concentration of ROS. The cells were washed by PBS for three times and incubated with DHE in PBS for 30 min in dark. Images were obtained by CLSM with the excited wavelength of 559 nm and analyzed by scanning densitometry.
2.2. PTDHP antioxidant and light regulation experimentsRat aortic cells were cultured as above. In the primary experiment, following incubated with PTDHP (2 × 10-5 mol/L) for 1 h and the endothelial cells Ang Ⅱ (1 × 10-7 mol/L) for 8 h. For control of experimental condition, DHP (5 × 10-6 mol/L) and PT (2 × 10-5 mol/L) were incubated with cells, respectively. For blank control group, cells were incubated with Ang Ⅱ only. For light group, after incubated with PTDHP, cells were exposed to white light with the dose of 0.4 mW/cm2 for 10 min. DHE (5 × 10-6 mol/ L) was applied to terrify the concentration of ROS. All cells were washed by PBS for three times and incubated with DHE in PBS for 30 min in dark. Images were obtained by CLSM with the excited wavelength of 559 nm and analyzed by scanning densitometry. Here, the white light source used in this work was equipped with a metal halogen lamp (MVL-210, Mejiro Genossen, Japan), and a radiometer (Photoelectric Instrument Factory of Beijing Normal University) was used to estimate the intensity of the incident beam.
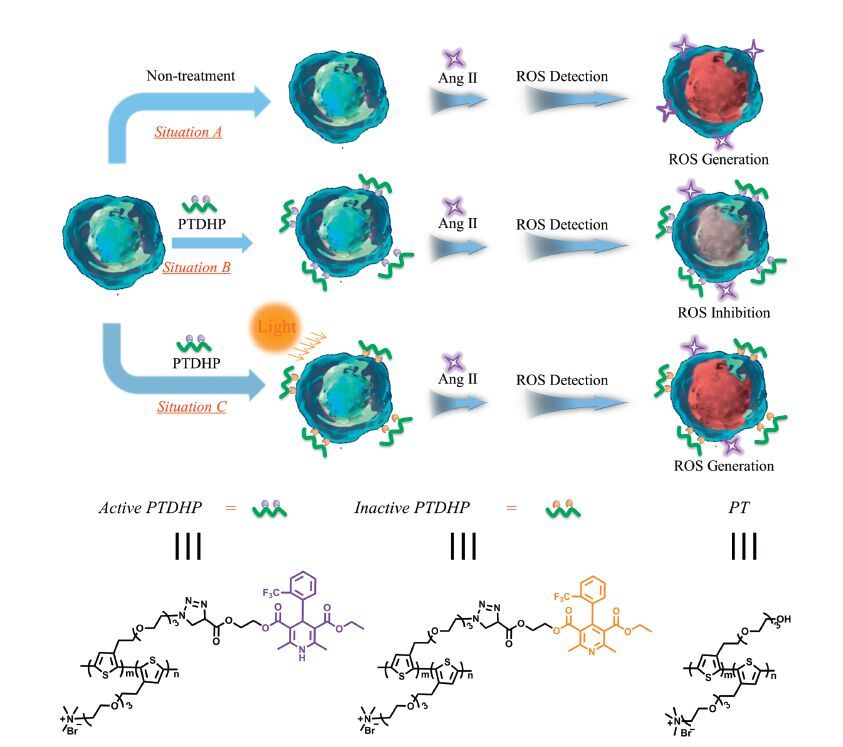
3. Results and discussionScheme 1 shows the mechanism of inhibition of oxidative stress induced by Ang Ⅱ via polythiophene and regulation of antioxidative ability by light. Stimulation of Ang Ⅱ will promote oxidative stress by increasing ROS generation in rat aortic endothelial cells, and hence cells can be stained by ROS probe obviously (situation A). However this process can be prevented by PTDHP, accounting to the antioxidative ability of DHP, which have been demonstrated that DHP derivatives could reduce the generation of ROS by the inhibition of the overexpression of nicotinamide adenine dinucleotide (NADPH) oxidase in response to Ang Ⅱ stimulation (situation B). When exposed to white light, the anti-oxidative potency was damaged by ROS generated through polythiophene sensitization, since dihydropyridine structure, which is the pharmacophore of DHP, can be destroyed by ROS (situation C). The chemical structures of active PTDHP, inactive PTDHP and unmodified polythiophene (PT) are shown in Scheme 1 as well.

|
Download:
|
| Scheme 1.The regulation of Ang Ⅱ-mediated ROS generation in living cells in the absence (situation A) and presence (situation B) of PTDHP, and the regulation of antioxidative effect under light (situation C). | |
{kind=link}
It was established that PTDHP displayed a good biocompatibility with the concentration ranging from 0 to 32 µmol/L, and preferred to interacting with cell membrane. Given that DHP can down regulate the expression of NADPH oxidase induced by Ang Ⅱ, which is amajor source of ROS generation in endothelial cells, to reduce the production of ROS, the antioxidate property of PTDHP was studied. Initially, Ang Ⅱ was verified to cause an enhancement in production of ROS. Cellswere incubated withAng Ⅱ for 0 h, 3 h, 6 h, 8 h and 10 h, respectively, in order to determine the production of ROS. Considering that DHE is oxidized into ethidium bromide (EB) which binds to DNA and affords red fluorescence in the presence of ROS, DHE was employed to observe the production of ROS in this work. The result demonstrated that the incubation with Ang Ⅱ caused an increase in the amount of ROS generation, which achieved its maximumafter8 h(Fig. 1).As a result, cellswere in cubated with Ang Ⅱ for 8 h in all subsequent experiments. Meanwhile, cells incubated with H2O2 were set as the positive group.

|
Download:
|
| Fig. 1.CLSM fluorescent images of rat aortic endothelial cells incubated with Ang Ⅱ for 0 h, 3 h, 6 h, 8 h and 10 h and stained with DHE. Cells treated with H2O2 was set as positive control. EB was highlighted in red. | |
{kind=link}
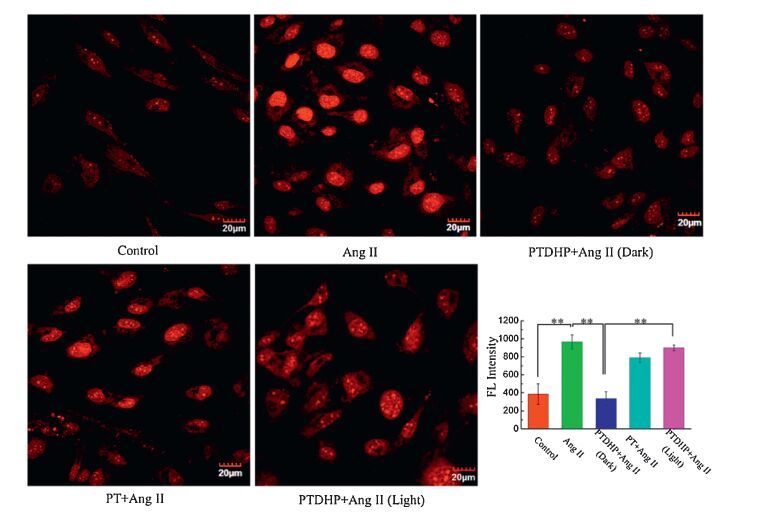
To testify the intrucellular antioxidate of PTDHP, cells were pretreated with PTDHP for 1 h before incubation with Ang Ⅱ. As shown in Fig. 2, the fluorescence intensity of DHE in rat aortic endothelial cells pretreated with PTDHP was dramatically weaker than that of cells only incubated with Ang Ⅱ, which indicated PTDHP could inhibit of production of ROS in rat aortic endothelial cells. It was interesting that cells incubated with PT achieved more infirm fluorescence than Ang Ⅱ group as well, revealing that PT could suppress the ROS generation induced by Ang Ⅱ in some degree. From the above experimental observations, PTDHP could depress ROS generation effectively and we speculated the reason for the inhibition activity of PT was that the polymer bounded to and localized on cell membrane leading to block the interaction of Ang Ⅱ to AT1 in some degree, which helpfully realized the suppression of ROS in cells. In addition, the majority distribution of PTDHP on cell membrane was beneficial for the interaction and the long retention to binding site for DHP to realize intracellular antioxidant, since DHP was internalized by rat aortic endothelial cells quickly.

|
Download:
|
| Fig. 2.Superoxide generation in rat endothelial aortic cells pretreated with PTDHP and PT, respectively, followed with Ang Ⅱ for 8 h in culture medium. The light group was exposed to white light with the dose of 0.4 mW/cm2 for 10 min after incubation with PTDHP. Untreated cells set as control group. [PTDHP] = 2 × 10-5 mol/L, [PT] = 2 × 10-5 mol/L, [Ang Ⅱ] = 1 × 10-7 mol/L, [DHE] = 5 × 10-6 mol/L. The graph shows the quantitative analysis of ROS production, analyzed by scanning densitometry. The error is the standard deviation from the mean (n = 3, * P < 0.05, ** P < 0.01). | |
{kind=link}
We further observed the light operation of the anti-oxidative effect of PTDHP based on the fact that the dihydropyridine structure can be oxidized to pyridine ring by ROS. Rat aortic endothelial cells were incubated with PTDHP followed by exposure to white light to destroy the structure of DHP, and the fluorescence intensity of DHE was counted. As shown in Fig. 2, the fluorescence intensity ofDHE in cells incubated with PTDHP upon exposing to light was much stronger than that of cells incubated with PTDHP under dark, which indicated that the anti-oxidative effect of PTDHP was disrupted by the ROSproduced by polymer sensitization. Besides, our experiment has verified that exposure to light could not lead to the enhancement of fluorescence intensity of DHE.
In the light of previous results, we further verified the outcomes from the genetic level and utilized the RNA isolation and reverse transcription-polymerase chain reaction (RT-PCR) to observe the expression of mRNA related to inflammation or ROS generation. The RT-PCR primers sequences and PCR protocols were operated according to literature [17]. LOX-1 can enhance the activity of NADPH oxidase in vascular endothelial cells and is suggested as an essential role in the further generation of ROS, in addition, the secretion of MCP-1 critically is involved in the ROS production via NADPH oxidase. VCAM-1 is a type of inflammatory molecules and related to cardiovascular disease caused by the over-activation of Ang Ⅱ. As shown in Fig. 3, the LOX-1 mRNA level in rat aortic endothelial cells incubated with Ang Ⅱ was increased significantly compared to the control group. The expressions of protein which are responsible for inflammation and even cardiovascular disease, can be enhanced by Ang Ⅱ inducement. LOX-1 was identified as a kind of scanvenger receptor, whose expression can be unregulated in atherosclerotic tissues related to the activation of Ang Ⅱ. The increased expression of LOX-1 mRNA level was prevented by the pretreatment of PTDHP. However, the expression of LOX-1 mRNA increased trough exposing to white light compared to cells treated by PTDHP in dark. Meanwhile, the increase of LOX-1 mRNA level induced by Ang Ⅱ stimulation was suppressed by pretreatment of PT in some degree. The outcome was perfectly consistent with the result of cell imaging assay. In addition, similar behaviors were observed with the mRNA levels for MCP-1 and VCAM-1. These results indicated that PTDHP can control the intracellular oxidative stress effectively, and polymers located on cell membrane and prevent the interaction of Ang Ⅱ to AT1, which can realize the suppression of ROS to some degree. Furthermore, the light regulation of anti-oxidative effect was achieved.

|
Download:
|
| Fig. 3.RT PCR assay of the expression of mRNA for LOX-1, MCP-1 and VCAM-1 in rat aortic endothelial cells with PTDHP, PT and DHP, respectively, for 1 h at 37 ℃, followed with stimulation with Ang Ⅱ for 8 h in culture medium. Untreated cells set as control. [PT] = 2 × 10-5 mol/L in RUs. [PTDHP] = 2 × 10-5 mol/L in RUs. [Ang Ⅱ] = 1 × 10-7 mol/L. Quantitative analysis of expression of mRNA levels were analyzed by E-gel imager. The error is the standard deviation from the mean (n = 3, * P < 0.05, ** P < 0.01). | |
{kind=link}
In conclusion, the inhibition of intracellular oxidative stress induced by Ang Ⅱ stimulation and light regulation by polythiophene (PT) derivatives was carried out in this work. By modification with DHP groups, PTDHP exhibited anti-oxidative properties in living cells through inhibiting the activity of NADPH oxidase in response to Ang Ⅱ stimulation, which is the original source of ROS in mammalian cells. Besides, the PTDHP could sensitize surrounding oxygen molecules to generate ROS under light irradiation inducing the oxidization of pendant DHP into pyridine group, which inactivated the control ability of DHP to oxidative stress in living cells and destroyed the pharmacophore of DHP. The antioxidative and anti-inflammatory properties of PTDHP might provide an insight into the protection against the progression of cardiovascular events.
AcknowledgmentThe authors are grateful to the National Natural Science Foundation of China (Nos. 21473220, 21373243, 21473221).
| [1] | R. Kumar, C.M. Thomas, Q.C. Yong, W. Chen, K.M. Baker, The intracrine renin-angiotensin system, Clin. Sci. 123(2012) 273-284. |
| [2] | A.R. Brasier, A. Recinos, M.S. Eledrisi, Vascular inflammation and the renin-angiotensin system, Arterioscler. Thromb. Vasc. Biol. 22(2002) 1257-1266. |
| [3] | C. Unterberg, H. Kreuzer, A.B. Buchwald, Renin-angiotensin system and cardiovascular diseases, Med. Klin. 93(1998) 416-425. |
| [4] | C. Tikellis, R.J. Pickering, D. Tsorotes, et al., Activation of the renin-angiotensin system mediates the effects of dietary salt intake on atherogenesis in the apolipoprotein E knockout mouse, Hypertension 60(2012) 98-105. |
| [5] | H. Lu, L.A. Cassis, A. Daugherty, Atherosclerosis and arterial blood pressure in mice, Curr. Drug Targets 8(2007) 1181-1189. |
| [6] | A.M. Garrido, K.K. Griendling, NADPH oxidases and angiotensin Ⅱ receptor signaling, Mol. Cell. Endocrinol. 302(2009) 148-158. |
| [7] | Q. Felty,W.C.Xiong,D.M. Sun,etal.,Estrogen-inducedmitochondrial reactiveoxygen species as signal-transducing messengers, Biochemistry 44(2005) 6900-6909. |
| [8] | H. Sauer, M. Wartenberg, J. Hescheler, Reactive oxygen species as intracellular messengers during cell growth and differentiation, Cell. Physiol. Biochem. 11(2001) 173-186. |
| [9] | B.C. Dickinson, C.J. Chang, A targetable fluorescent probe for imaging hydrogen peroxide in themitochondria of living cells, J.Am. Chem. Soc.130(2008) 9638-9639. |
| [10] | W. Dröge, Free radicals in the physiological control of cell function, Physiol. Rev. 82(2002) 47-95. |
| [11] | T. Volk, M. Hensel, H. Schuster, W.J. Kox, Secretion of MCP-1 and IL-6 by cytokine stimulated production of reactive oxygen species in endothelial cells, Mol. Cell. Biochem. 206(2000) 105-112. |
| [12] | C.P. Hu, A. Dandapat, J.L. Mehta, Angiotensin Ⅱ induces capillary formation from endothelial cells via the LOX-1-dependent redox-sensitive pathway, Hypertension 50(2007) 952-957. |
| [13] | M. Nahrendorf, E. Keliher, P. Panizzi, et al., 18F-4V for PET-CT imaging of VCAM-1 expression in atherosclerosis, JACC-Cardiovasc. Imag. 2(2009) 1213-1222. |
| [14] | M.F. Navedo, L.F. Santana, CaV 1.2 sparklets in heart and vascular smooth muscle, J. Mol. Cell. Cardiol. 58(2013) 67-76. |
| [15] | A. Neely, P. Hidalgo, Structure-function of proteins interacting with the α1 poreforming subunit of high-voltage-activated calcium channels, Front. Physiol. 5(2014) 209. |
| [16] | R. Berkels, T. Breitenbach, H. Bartels, et al., Different antioxidative potencies of dihydropyridine calcium channel modulators in various models, Vascul. Pharmacol. 42(2005) 145-152. |
| [17] | H. Toba, T. Shimizu, S. Miki, et al., Channel blockers reduce angiotensin Ⅱ-induced superoxide generation and inhibit lectin-like oxidized low-density lipoprotein receptor-1 expression in endothelial cells, Hypertens. Res. 29(2006) 105-116. |
| [18] | N. Ishii, T. Matsumura, S. Shimoda, E. Araki, Anti-atherosclerotic potential of dihydropyridine calcium channel blockers, J. Atheroscler. Thromb. 19(2012) 693-704. |
| [19] | J.L. Yeh, J.H. Hsu, J.C. Liang, I.-J. Chen, S.-F. Liou, Lercanidipine and labedipinedilol-A attenuate lipopolysaccharide/interferon-gamma-induced inflammation in rat vascular smooth muscle cells through inhibition of HMGB1 release and MMP-2, 9 activities, Atherosclerosis 226(2013) 364-372. |
| [20] | B. Wang, J.Z. Song, H.X. Yuan, et al., Multicellular assembly and light-regulation of cell-cell communication by conjugated polymer materials, Adv. Mater. 26(2014) 2371-2375. |
| [21] | C.L. Zhu, L.B. Liu, Q. Yang, F.T. Lv, S. Wang, Water-soluble conjugated polymers for imaging, diagnosis, and therapy, Chem. Rev. 112(2012) 4687-4735. |
| [22] | A.V. Ambade, B.S. Sandanaraj, A. Klaikherd, S. Thayumanavan, Fluorescent polyelectrolytes as protein sensors, Polym. Int. 56(2007) 474-481. |
| [23] | H.-A. Ho, A. Najari, M. Leclerc, Optical detection of DNA and proteins moth cationic polythiophenes, Acc. Chem. Res. 41(2008) 168-178. |
| [24] | S. Kim, C.-K. Lim, J. Na, et al., Conjugated polymer nanoparticles for biomedical in vivo imaging, Chem. Commun. 46(2010) 1617-1619. |
| [25] | F. Qiu, Q. Zhu, G.S. Tong, et al., Highly fluorescent core-shell hybrid nanoparticles templated by a unimolecular star conjugated polymer for a biological tool, Chem. Commun. 48(2012) 11954-11956. |
| [26] | B.Q. Bao, N.J. Tao, D.L. Yang, et al., A multi-core-shell structured composite cathode material with a conductive polymer network for Li-S batteries, Chem. Commun. 49(2013)10263-10265. |
| [27] | K. Liu, Y.L. Liu, Y.X. Yao, et al., Supramolecular photosensitizers with enhanced antibacterial efficiency, Angew. Chem. Int. Ed. 52(2013) 8285-8289. |
| [28] | C.F. Xing, Q.L. Xu, H.W. Tang, L.B. Liu, S. Wang, Conjugated polymer/porphyrin complexes for efficient energy transfer and improving light-activated antibacterial activity, J. Am. Chem. Soc. 131(2009) 13117-13124. |
| [29] | C.L. Zhu, Q. Yang, L.B. Liu, et al., Multifunctional cationic poly(p-phenylene vinylene) polyelectrolytes for selective recognition, imaging, and killing of bacteria over mammalian cells, Adv. Mater. 23(2011) 4805-4810. |
| [30] | R. Hu, F.Y. Wang, S.L. Li, et al., ROS self-scavenging polythiophene materials for cell imaging, Polym. Chem. 6(2015) 8244-8247. |