




DNA methylation is regarded as a critical biochemical reaction which has been implicated in the regulation of a variety of biological processes across virtually every branch of the taxonomic tree [1, 2, 3]. DNA methylation is achieved by the catalysis of DNA methyltransferases (MTase),which specifically recognize short palindromic sequences and catalyze the transfer of a methyl group from several different donors to the target adenine or cytosine. DNA methylation is associated with the formation of heterochromatin and silencing of genes critical for the regulation of growth and proliferation [4, 5]. This covalent modification of DNA functions as an important mediator of gene regulation and, together with covalent modifications of histone proteins,also forms the cornerstone for the burgeoning field of epigenetics. Cancer cells are characterized by hypomethylation throughout their genomes,plus hypermethylation of specific regions,which has been associated with transcriptional silencing [6, 7]. Because of its key role in pathology and diagnostics,analysis of DNA methylation and MTase activities has increasingly received research interests. A few methods have been developed for the determination of DNA methylation and MTases activity,including polymerase chain reaction (PCR)-based techniques [8, 9],light scattering techniques [10],colorimetric approaches [11, 12, 13], capillary electrophoresis [14],high-performance liquid chromatography (HPLC) [15],fluorescence [16, 17, 18, 19],electrochemical [20, 21],and chemiluminescence methods [22],etc. However, most of those methods have shortcomings,like being timeintensive, laborious treatment,radiolabeling substrate requirement, or immobilization of a target analyte to surfaces.
In recent years,isothermal amplification techniques,such as rolling circle amplification (RCA) [23],strand displacement amplification (SDA) [24],and nicking enzyme signal amplification (NESA) [25],have been frequently used for an enzyme assay due to their wide variety of choices and significant signal amplification. Among these amplification methods,the SDA [26] and NESA [27] methods have been used for the DNA MTase assay. RCA technology has proven very useful for highly sensitive detection of target nucleic acids and proteins [28, 29]. In RCA-based assays,target quantification is achieved through the quantification of the RCA products. In this study,we adopted rolling circle amplification technique and molecular beacons to develop a sensitive and selective method to detect DNA methyltransferase activity. As shown as Scheme 1,we used DNA adenine methylation (Dam) MTase and restriction endonuclease Dpn I endonuclease,which both had the same 5'-G-A-T-C-3' recognition sequence. Dam MTase catalyzes the methylation of the sequence 5'-G-A-T-C-3', and restriction endonuclease Dpn I cleaves the sequence 5'-G-Am-T-C-3'. The cleaved short oligonucleotide can hybridize with the circular DNA template and trigger RCA. Each RCA product containing a large number of repeated sequences might hybridize with a large number of molecular beacons (detection probes), resulting in an enhanced fluorescence signal. In contrast,in the absence of Dam MTase,the unmethylated DNA cannot be cleaved by the restriction endonuclease Dpn I. At the same time,Bst polymerase (large fragment) only can copy complementary sequences from the 3' end of oligonucleotides,and the polymer reaction cannot be triggered. Therefore,no obvious change in the fluorescence intensity of the detection probe is observed. The unique ability of restriction endonuclease Dpn I to transduce DNA adenine methylation to sequence information was melded perfectly with the capabilities of RCA to amplify nucleic acid sequences.

|
Download:
|
| Scheme 1.Schematic diagram of DNA methylation detection based on the Dam MTase-restriction endonuclease Dpn I interaction system and RCA technology. | |
Oligonucleotides designed in this study were synthesized by Takara Biotechnology Co.,Ltd. (Dalian,China),and the sequences of all oligonucleotides are listed in Table 1. The underlined portion in the probe is the specific recognize sequence for Dam methyltransferase. The Dam MTase (Escherichia coli),restriction endonuclease Dpn I,Bst DNA polymerase (large fragment),Sadenosylmethionine (SAM),dNTP and the corresponding buffer solution were purchased from New England Biolabs Inc. (NEB, U.K.). The gentamycin sulfate was purchased from Bo Mei Biotechnology Co.,Ltd. Other chemicals were of analytical grade and were used without further purification. The solutions in the experiments were prepared with ultrapure water (Milli-Q 18.2 MΩ cm,Millipore System Inc.). Several buffers were used in the present work: The methylation buffer contained 10 mmol/L Tris-HCl (pH 7.5 @ 25°C),15 mmol/L MgCl2,50 mmol/L NaCl,and 1 mmol/L DTT. The RCA-reaction buffer was composed of 20 mmol/L Tris-HCl (pH 8.8 @ 25°C),10 mmol/L KCl,2 mmol/L MgSO4,10 mmol/L (NH4)2SO4,and 0.1% Triton X-100.
| Table 1 Oligonucleotides |

{kind=link}
The reaction mixtures (200 mL) consisted of a 200 nmol/L hairpin probe,1× methylation buffer,160 μmol/L SAM,20 units of restriction endonuclease Dpn I,and 40 units of Dam MTase. In the gel electrophoresis assay,5 μL samples were removed from the reaction solution after having been incubated at 37°C for 90 min and then heated at 65°C for 15 min to stop the reaction. Subsequently,the samples were put on a polyacrylamide gel (20% acrylamide,19:1 acrylamide/bisacrylamide) to separate the cleaved products from the substrate. The electrophoresis was carried in 1× TBE (pH 8.0) at 100 V constant voltage for 3 h,and then the gel was scanned using the Gel Image Analysis System (Tanon 2500R,Shang Hai,China). 2.3. Apparatus and fluorescence measurements
Fluorescence spectra were measured using a Hitachi F-7000 fluorescence spectrometer (Hitachi,Japan) with a personal computer data processing unit. Excitation and emission slits were all set for 5.0 nm band-pass. The fluorophore of FAM was excited at 488 nm,and the emission spectra from 500 nm to 600 nm were collected. The fluorescence intensity at 521 nm was used to evaluate the performance of the proposed assay strategy. All measurements were carried out at room temperature unless stated otherwise. 2.4. RCA and Dam MTase detection
A Dam MTase assay buffer solution was prepared in an Eppendorf tube containing 1× methylation buffer,1 μmol/L substrate oligonucleotide (namely Prime),160 μmol/L SAM,20 units of restriction endonuclease Dpn I,and a different activity of Dam MTase. The methylation was allowed to proceed at 37°C for 90 min and then heated at 65°C for 15 min to stop the reaction. Then,20 μL of the resulting solution was added to 20 mL of RCAreaction buffer containing 1× RCA-reaction buffer,500 mmol/L dNTPs and 0.2 U/μL Bst DNA polymerase (large fragment). The polymerization reaction was carried out at 60°C for 60 min. Subsequently,the resulting solution was incubated at 80°C for 20 min to inactivate the Bst DNA polymerase (large fragment). After the resulting solution was cooled to ambient temperature, the detection probes were added and allowed to hybridize with the RCA products for about 40 min prior to the fluorescence measurement. 3. Results and discussion
3.1. Design and feasibility of RCA based Dam MTase assay
As mentioned above,in the presence of Dam MTase,the enzyme will catalyze the methylation reaction on the recognition sequence to yield the methylation duplex DNA 50-G-Am-T-C-30. Then,the cleavage reaction by restriction endonuclease Dpn I was initiated, and the cleaved short oligonucleotide hybridized with the circular DNA template and triggered rolling circular amplification. The fluorescence signal was acquired after RCA product sequences opened the molecular beacons.
To confirm our assay method,gel electrophoresis experiments were carried out. As shown in Fig. 1A,new bands appear in lane 3 when both Dam MTase and restriction endonuclease Dpn I are added in the test solution,suggesting that a methylation reaction has taken place,and the methylated DNA had been cleaved into small pieces. As a comparison,it can be observed that there is only one band of the original probe in lanes 1 and 2 when the Dam MTases are absent,indicating that no methylation/ cleavage reaction occurs. Fig. 1B shows the fluorescence signal at λex/λem = 488/521 nmof three samples: curve a is for assay buffer, restriction endonuclease Dpn I with the addition of 200 U/mL Dam MTase; curve b is for assay buffer,restriction endonuclease Dpn I with the addition of 20 U/mL Dam MTase,and curve c is for assay bufferwith restriction endonuclease Dpn I,in absence of DamMTase. It can be observed that there is no signal enhancement in curve c, indicating that restriction endonuclease Dpn I will not cleave the DNA in the absence ofDam MTase. Nevertheless,as is shown in curve a and curve b,for the case that both restriction endonuclease Dpn I and Dam MTase are present,an obvious fluorescence at 521 nm can be detected. These experimental data also clearly confirm that hairpin molecular beacons fluorescence quenching as a result of the cleavage reaction cannot be initiated if the DNA methylation has not been achieved,which are in accordance with the results provided by the gel electrophoresis assay.

|
Download:
|
| Fig. 1.(A) The gel electrophoresis image of incubation samples for 90 min. Lanes: (1) hairpin probe; (2) hairpin probe with restriction endonuclease Dpn I; (3) hairpin probe, restriction endonuclease Dpn I,and Dam methyltransferase. (B) Fluorescence emission spectra of the detection probe in assay system in the different activity unit of Dam MTase. | |
{kind=link}
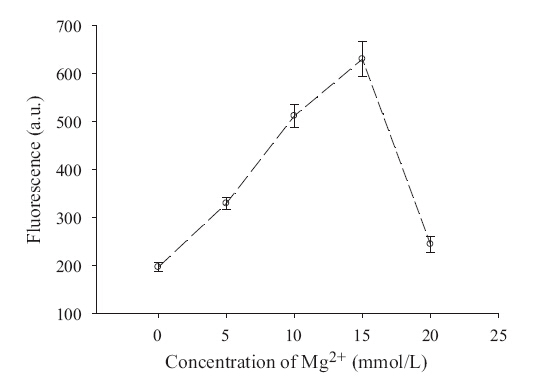
In the reaction system,magnesium ion was a critical factor. It is not only associated with the activity of methyltransferases,but also plays a key role in the hybridization and polymerization of RCA. Therefore,the effect of the concentration of magnesium ion was studied. A 200 mL Dam MTase assay buffer solution was prepared in a 200 mL Eppendorf tube containing 10 mmol/L Tris- HCl (pH 7.5 @ 25°C),50 mmol/L NaCl,1 mmol/L DTT,1 mmol/L hairpin probe,160 mmol/L SAM,200 U/mL restriction endonuclease Dpn I,20 U/mL Dam Mtase,and different concentrations of MgCl2. Fig. 2 indicates that the methylation/cleavage reaction also happened,but with a very low velocity if no Mg2+ existed in the reaction mixture. The activity of methylation/cleavage reached its maximum when the concentration of Mg2+ was 15 mmol/L,and the fluorescence decreased when the concentration of Mg2+ was above 15 mmol/L. It is well known that Mg2+ is an essential coenzyme for restriction endonuclease (including restriction endonuclease Dpn I) and will hinder the process of methylation if its concentration is very high. Experimental results reveal that 15 mmol/L Mg2+ is the optimum concentration that offers the highest activity of DNA methylation for our system; thus,we have chosen 15 mmol/L Mg2+ in the following experiments.

|
Download:
|
| Fig. 2.Optimization of magnesium ions in the reaction system. The fluorescence intensity of the detection probe contains various concentration of the Mg2+ range from 0 to 20 mmol/L for the reaction system. (Error bars were deduced from N = 3 experiments). λex/λem = 488/521 nm,slit width: 5 nm. | |
{kind=link}
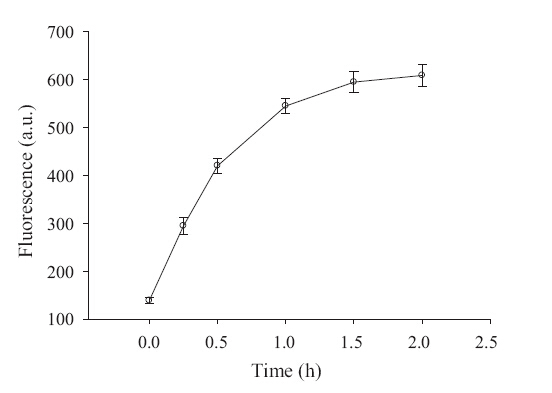
For this study,we developed an assay that used a simple hairpin probe to monitor the DNA methylation process by coupling it with the RCA reaction. A 200 mL Dam MTase assay buffer solution was prepared in a 200 μL Eppendorf tube containing 1× methylation buffer,1 μmol/L hairpin probe,160 mMSAM,200 U/mL restriction endonuclease Dpn I,and 20 U/mL Dam MTase. The methylation was allowed to proceed at 37°C for 120 min. Then,20 mL of the reaction solution was extracted respectively at t0 = 0 min, t1 = 15 min,t2 = 30 min,t3 = 60 min,t4 = 90 min,and t5 = 120 min and added in the Eppendorf tube. After incubation at 65°C for 20 min to stop methylation,the resulting solution was added to 20 μL of RCA-reaction buffer and trigger RCA. Fig. 3 shows the detection fluorescence of the reaction system at various points of time. At 90 min,the reaction system was relatively steady-going and close to the maximal response.

|
Download:
|
| Fig. 3.Total time course of DNA methylation. The concentration of Dam MTase is 20 U/mL. The error bars represent the standard deviation of three measurements. | |
{kind=link}
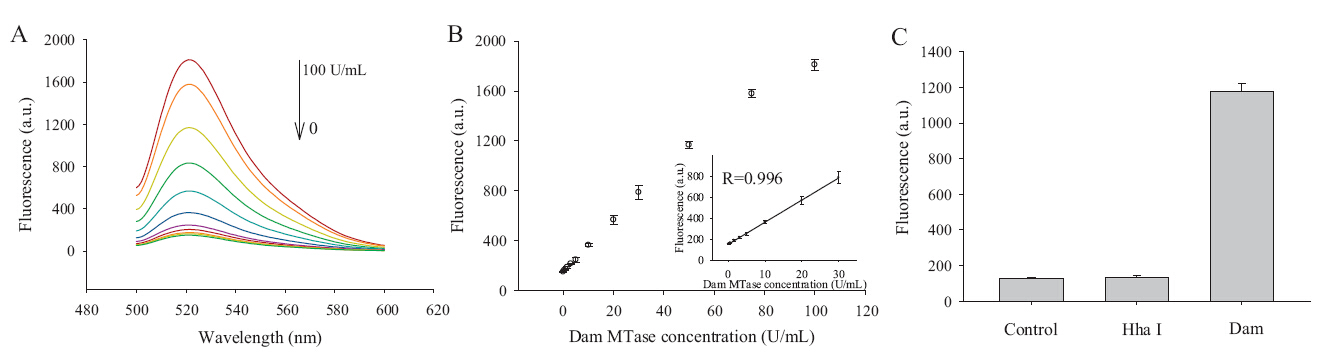
To investigate the analytical performance of the proposed method,we measured the Dam MTase at various concentrations under the optimal conditions. Fig. 4A shows the variance of fluorescence intensity with the concentration of Dam MTase. As the concentration of Dam MTase increases,large numbers of primers are produced for RCA,resulting in the increase of fluorescence intensity correspondingly. Under the reaction conditions mentioned above,the fluorescence showed a clear dependence on concentration of Dam MTase with a linear relationship (R = 0.996) in the range of 0.5-30 nmol/L. The detection limit of the Dam MTase assays was determined to be ~0.18 U/mL (3s/slope,s is the standard deviation in 3 independent measurements of slope in Fig. 4B inset). In addition,we chose Hha I MTase (recognize 5’-G-C-G-C-3’) as an interference factor. Due to the specific site recognition of Dam MTase toward its substrate,the proposed method can easily discriminate Dam MTase from Hha I MTase. As shown in Fig. 4C,significant fluorescence enhancement is observed in the presence of Dam MTase. In contrast,no distinct fluorescence signal is observed in the presence of Hha I MTase, suggesting the high selectivity of the proposed method toward Dam MTase.

|
Download:
|
| Fig. 4.(A) The fluorescence spectra of molecular beacons obtained from RCA triggered by different concentrations of Dam MTase. From upper to bottom are 0,0.5,1.5,3,5,10, 20,30,50,70 and 100 U/mL. (B) The fluorescence intensity for various concentration of Dam MTase varying from 0 to 100 U/mL. Inset is the calibration curve for concentration ranging from 0 to 30 U/mL. (C) Selectivity of the proposed method. The concentration of Dam MTase is 50 U/mL,and the concentration of Hha I MTase is 50 U/mL. Measurements were performed in 20 mmol/L Tris-HCl (pH 8.8 @ 25°C),10 mmol/L KCl,2 mmol/L MgSO4,10 mmol/L (NH4)2SO4,and 0.1% Triton X-100 (error bars were deduced from N = 3 experiments). λex/λem = 488/521 nm,slit width: 5 nm. | |
{kind=link}
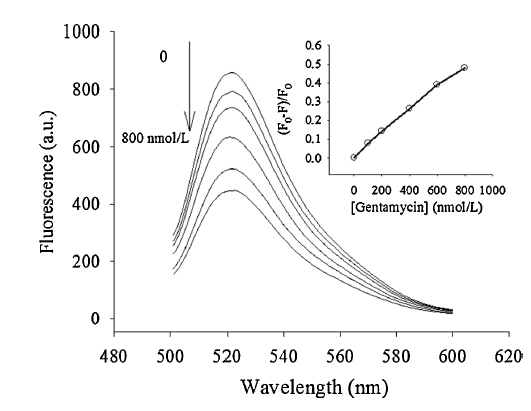
DNA MTase exists extensively in both prokaryotes and eukaryotes,which means the pharmacological inhibition of DNA MTase provides a broad spectrum of therapeutic applications. To further extend the potential application of this assay in the inhibition assay,the influence of drugs on Dam MTase activity has been investigated by using gentamycin as a model direct inhibitor of Dam MTase. The mechanism is that,if transferring of themethyl group to the DNA residue is blocked at the presence of inhibitor,the specific site of the DNA molecules will not be recognized and cleaved; thus,the RCA will not be triggered and the fluorescence probe will keep the state of quenching. Experimental results show that the inhibitory effect of the inhibitor can be indeed quantitatively analyzed using the fluorescence intensity ratio,(F0 - F)/F0 (see Fig. 5). Therefore, this method presented here has the potential to evaluate drugs for antimicrobial and anticancer activity through influencing the activity of Dam MTase.

|
Download:
|
| Fig. 5.Inhibition of the activity of Dam MTase by gentamycin. Fluorescence courses of samples with addition of different amounts of gentamycin: 0,100,200,400,600 and 800 nmol/L. Inset is the calibration curve for gentamycin concentrations ranging from 0 to 800 nmol/L. F0,F are the fluorescence intensity in the absence of gentamycin and in the presence of gentamycin,respectively. | |
{kind=link}
In summary,we have described a new method for the detection of the genomic DNA methylation level based on the Dam MTase- restriction endonuclease Dpn I interaction system and RCA technology. This new assay method takes advantage of specific recognition of the MTase and methyl-sensitive endonuclease as well as the unique properties of Bst polymerase (large fragment). Compared with the traditional methods for methylation detection, this proposed assay method can be very easily operated and achieved a lower detection limit of 0.18 U/mL. In addition,we also evaluated the inhibitors of methyltransferase,which may be of help in the discovery of anticancer drugs. Acknowledgments
This work was supported by the National Natural Science Foundation of China (Nos. 21190044 and 21175035),National Basic Research Program (No. 2011CB911002),and International Science & Technology operation Program of China (No. 2010DFB30300).
| [1] | M.G. Marinus, J. Casadesus, Roles of DNA adenine methylation in host-pathogen interactions: mismatch repair, transcriptional regulation, and more, Fems. Microbiol Rev. 33 (2009) 488-503. |
| [2] | S.J. Cokus, S.H. Feng, X.Y. Zhang, et al., Shotgun bisulphite sequencing of the arabidopsis genome reveals DNA methylation patterning, Nature 452 (2008) 215-219. |
| [3] | R. Lister, R.C. O'Malley, J. Tonti-Filippini, et al., Highly integrated single-base resolution maps of the epigenome in arabidopsis, Cell 133 (2008) 523-536. |
| [4] | C.B. Klein, M. Costa, DNA methylation and gene expression: introduction and overview, Mutat. Res. 386 (1997) 103-105. |
| [5] | A. Bird, DNA methylation patterns and epigenetic memory, Gene Dev. 16 (2002) 6-21. |
| [6] | A.P. Feinberg, B. Tycko, The history of cancer epigenetics, Nat. Rev. Cancer 4 (2004) 143-153. |
| [7] | P.A. Jones, S.B. Baylin, The epigenomics of cancer, Cell 128 (2007) 683-692. |
| [8] | F. Lyko, B.H. Ramsahoye, R. Jaenisch, DNA methylation in drosophila melanogaster, Nature 408 (2000) 538-540. |
| [9] | I. Hatada, Y. Hayashizaki, S. Hirotsune, et al., A genomic scanning method for higher organisms using restriction sites as landmarks, Proc. Natl. Acad. Sci. U.S.A. 88 (1991) 9523-9527. |
| [10] | C. Zhao, K.G. Qu, Y.J. Song, et al., A universal, label-free, and sensitive optical enzyme-sensing system for nuclease and methyltransferase activity based on light scattering of carbon nanotubes, Adv. Funct. Mater. 21 (2011) 583-590. |
| [11] | T. Liu, J. Zhao, D.M. Zhang, G.X. Li, Novel method to detect DNA methylation using gold nanoparticles coupled with enzyme-linkage reactions, Anal. Chem. 82 (2010) 229-233. |
| [12] | W. Li, Z.L. Liu, H. Lin, et al., Label-free colorimetric assay for methyltransferase activity based on a novel methylation-responsive DNAzyme strategy, Anal. Chem. 82 (2010) 1935-1941. |
| [13] | G.T. Song, C.E. Chen, J.S. Ren, et al., A simple, universal colorimetric assay for endonuclease/methyltransferase activity and inhibition based on an enzymeresponsive nanoparticle system, ACS Nano 3 (2009) 1183-1189. |
| [14] | M.F. Fraga, R. Rodríguez, M.J. Cañal, Rapid quantification of DNA methylation by high performance capillary electrophoresis, Electrophoresis 21 (2000) 2990-2994. |
| [15] | S. Friso, S.W. Choi, G.G. Dolnikowski, et al., A method to assess genomic DNA methylation using high-performance liquid chromatography/electrospray ionization mass spectrometry, Anal. Chem. 74 (2002) 4526-4531. |
| [16] | J. Li, H.F. Yan, K.M. Wang, W.H. Tan, X.W. Zhou, Hairpin fluorescence DNA probe for real-time monitoring of DNA methylation, Anal. Chem. 79 (2007) 1050-1056. |
| [17] | X.L. Wang, Y.L. Song, M.Y. Song, et al., Fluorescence polarization combined capillary electrophoresis immunoassay for the sensitive detection of genomic DNA methylation, Anal. Chem. 81 (2009) 7885-7891. |
| [18] | R.J. Wood, J.C. McKelvie, M.D. Maynard-Smith, P.L. Roach, A real-time assay for CpG-specific cytosine-C5 methyltransferase activity, Nucleic Acids Res. 38 (2010) e107. |
| [19] | F.D. Feng, H.Z. Wang, L.L. Han, S. Wang, Fluorescent conjugated polyelectrolyte as an indicator for convenient detection of DNA methylation, J. Am. Chem. Soc. 130 (2008) 11338-11343. |
| [20] | P. Wang, Z.B. Mai, Z. Dai, X.Y. Zou, Investigation of DNA methylation by direct electrocatalytic oxidation, Chem. Commun. 46 (2010) 7781-7783. |
| [21] | X.X. He, J. Su, Y.H. Wang, et al., A sensitive signal-on electrochemical assay for MTase activity using AuNPs amplification, Biosens. Bioelectron. 28 (2011) 298-303. |
| [22] | Y.P. Zeng, J. Hu, Y. Long, C.Y. Zhang, Sensitive detection of DNA methyltransferase using hairpin probe-based primer generation rolling circle amplification-induced chemiluminescence, Anal. Chem. 85 (2013) 6143-6150. |
| [23] | J. Nosek, A. Rycovska, A.M. Makhov, J.D. Griffith, L. Tomaska, Amplification of telomeric arrays via rolling-circle mechanism, J. Biol. Chem. 280 (2005) 10840-10845. |
| [24] | H.Q. Wang, W.Y. Liu, Z. Wu, et al., Homogeneous label-free genotyping of single nucleotide polymorphism using ligation-mediated strand displacement amplification with DNAzyme-based chemiluminescence detection, Anal. Chem. 83 (2011) 1883-1889. |
| [25] | X. Hun, H. Chen, W. Wang, Design of ultrasensitive chemiluminescence detection of lysozyme in cancer cells based on nicking endonuclease signal amplification technology, Biosens. Bioelectron. 26 (2010) 248-254. |
| [26] | C.F. Zhu, Y.Q. Wen, H.Z. Peng, et al., A methylation-stimulated DNA machine: an autonomous isothermal route to methyltransferase activity and inhibition analysis, Anal. Bioanal. Chem. 399 (2011) 3459-3464. |
| [27] | F. Chen, Y. Zhao, Methylation-blocked enzymatic recycling amplification for highly sensitive fluorescence sensing of DNA methyltransferase activity, Analyst 138 (2013) 284-289. |
| [28] | F. Andrew, S.Q. Xu, Rolling replication of short DNA circles, Proc. Natl. Acad. Sci. U.S.A. 92 (1995) 4641-4645. |
| [29] | M.L. Li, D.R. Zhou, H. Zhao, J.K. Wang, Z.H. Lu, Endonuclease-rolling circle amplification-based method for sensitive analysis of DNA-binding protein, Chin. Chem. Lett. 20 (2009) 1315-1318. |