


|
收稿日期: 2018-03-29; 预印本: 2018-06-06
基金项目: 国家重点研发计划(编号:2017YFC0405804,2017YFC0405801);国家自然科学基金(编号:51309254);中国水利水电科学研究院基本科研业务费专项项目(编号:WR0145B272016,WR0145B012017);云南省重大水利项目(编号:YSZD-2014-001,YNWRM-2012-01)
第一作者简介: 曹引,1991年生,男,工程师,研究方向为流域水资源遥感及水循环模型数据同化。E-mail:yinc0416@163.com
通信作者简介: 冶运涛,1983年生,男,高级工程师,研究方向为水信息与智慧流域、流域水循环模拟及水资源遥感。E-mail:yeyuntao@iwhr.com
|
摘要
草型湖泊水质遥感监测中水生植物会造成“水体—水生植物”混合像元问题,针对因混合像元导致草型湖泊水生植物覆盖区域水质难以直接利用遥感监测的问题,本文以草型湖泊微山湖为研究对象,提出定性和定量相结合的总悬浮物浓度和浊度分区监测方法,实现微山湖水体总悬浮物浓度和浊度的时空变化监测。基于获取的2014年7月—2015年6月覆盖微山湖的多期高分一号(GF-1) WFV和HJ-1A/1B CCD影像,利用归一化水体指数将微山湖区分为水生植物覆盖区和水体区。针对水生植物覆盖区,利用时序MODIS NDVI数据获取微山湖主要水生植物的时谱曲线,识别不同水生植物的物候特征;基于不同物候期内的水生植物对总悬浮物浓度和浊度的指示作用,对微山湖水生植物覆盖区水体总悬浮物浓度和浊度进行定性监测。针对水体区,分别构建水体总悬浮物浓度和浊度的单波段/波段比值模型和偏最小二乘模型,定量反演微山湖水体区总悬浮物浓度和浊度。研究结果表明,微山湖中水生植物以光叶眼子菜、穗花狐尾藻和菹草等沉水植物为主,其中光叶眼子菜/穗花狐尾藻和菹草的空间分布和物候特征存在明显差异,不同水生植物在不同物候期内对水质具有不同的指示作用;微山湖水体总悬浮物浓度和浊度具有显著的空间变异性,基于定性和定量相结合的方法可以有效监测微山湖水体总悬浮物浓度和浊度的时空变化规律。本文提出的定性和定量相结合的监测方法为草型湖泊水质监测的业务化应用提供了新思路。
关键词
遥感, 草型湖泊, 水生植物, 时谱曲线, 物候, 遥感监测, 高分一号, HT-1
Abstract
Remote sensing has been recognized as an effective tool for monitoring water quality in inland waters, especially in algal lakes. However, water quality in macrophytic lakes where aquatic vegetation grows because of mixed pixels is difficult to retrieve. In this study, a classification retrieval method and image data acquired between July 2014 and June 2015 using GF-1 WFV and HJ-1A/1B CCD sensors were proposed to monitor Total Suspended Matter (TSM) and turbidity in a macrophytic lake, namely, the Weishan Lake, considering aquatic vegetation phenology. In the classification retrieval method, the Weishan Lake was divided into water overlying aquatic vegetation and water area using normalized difference water index. First, a qualitative method was proposed to retrieve TSM concentration and turbidity in water overlying aquatic vegetation considering aquatic vegetation phenology. In the qualitative method, time series MODIS NDVI data were used to obtain the time–spectrum curves of aquatic vegetation in the Weishan Lake for identifying different aquatic vegetation phenological periods. The characteristics of aquatic vegetation in different phenological periods were used to estimate the TSM concentration and turbidity. Second, single band, band ratio, and partial least squares models were applied to retrieve the TSM concentration and turbidity in the water area. Finally, the temporal and spatial variations in the TSM concentration and turbidity along with aquatic vegetation in the entire Weishan Lake were analyzed. Results showed that the three main aquatic vegetation areas in the Weishan Lake, namely, Potamogeton lucens, Myriophyllum spicatum, and Potamogeton crispus, have different phenological periods. P. lucens and M. spicatum started to grow in spring, reached their peak at the end of summer, and gradually died in autumn. P. crispus started to grow in April, reached its peak at the end of spring, and quickly died in the early summer. Different aquatic vegetation areas had various indicators of TSM concentration and turbidity in different phenological periods. The TSM concentration was less than 15 mg/L in the water overlying three aquatic vegetation areas in the growth stage. The turbidity in the water overlying P. lucens, M. spicatum, and P. crispus in the growth stage were less than 30 and 15 NTU. The death of P. lucens and M. spicatum did not result in the deterioration of water quality because of their extensive death time. However, the death of P. crispus within a short time resulted in serious deteriorations of water quality. The TSM concentration and turbidity were 15–145 mg/L and 30–140 NTU, correspondingly, in the water overlying P. crispus in the death stage. The TSM concentration and turbidity in the Weishan Lake had a significant temporal–spatial variability. The TSM concentration and turbidity in the Southwestern Weishan Lake where P. lucens and M. spicatum grew had low levels in the seasons. By contrast, the TSM concentration and turbidity in the Northeastern Weishan Lake with P. crispus growth had low levels in spring. However, they became large in summer because of the quick death of P. crispus and gradually decreased in autumn and winter. A classification retrieval method coupled with quantitative monitoring in the water overlying aquatic vegetation and qualitative monitoring in the water area was proposed to monitor the TSM concentration and turbidity in the Weishan Lake considering aquatic vegetation phenology. This method effectively monitored the temporal and spatial variations in water quality in the entire Weishan Lake. The findings indicated that the proposed method can be used to monitor the water quality in other macrophytic lakes.
Key words
remote sensing, macrophytic lake, aquatic vegetation, time-spectrum curve, phenology, remote sensing monitoring, GF-1, HT-1
1 引 言
湖泊水资源作为内陆水资源的重要组成部分,为人类提供大量水资源的同时具有防洪、养殖、航运以及生物多样性保护等重要功能(Hou 等,2017;Wang 等,2014)。然而随着社会经济的高速发展,日益增强的人类活动加剧了湖泊的污染趋势,导致湖泊呈现富营养化状态的形势愈加严峻,大面积蓝藻爆发时有发生(Matthews 等,2010;朱庆 等,2016;Duan 等,2009)。因此,加强湖泊水质的动态监测对实现其精细化和科学化管理具有重要意义。
遥感作为一种区域化监测手段,能够快速获取湖泊水质的时空分布,越来越多地应用于湖泊水环境的监测和管理(Dörnhöfer和Oppelt,2016;蒋云钟 等, 2010,2011,2014)。总悬浮物浓度和浊度是湖泊水质遥感监测的重用指标,其中总悬浮物是总氮、总磷和有机污染物的重要载体,同时影响着光在水中的衰减和垂直分布(Panigrahi 等,2009);浊度是和总悬浮物浓度密切相关的重要参数,能够影响光在水中的传播,对浮游植物生长和水体富营养化状况具有指示作用(Petus等,2010)。在叶绿素a浓度低的水体中,浊度主要由总悬浮物浓度决定(Güttler 等,2013)。总悬浮物浓度和浊度遥感监测主要基于遥感反射率与总悬浮物浓度和浊度之间的统计关系构建经验模型(线性或非线性回归模型) (Shi 等,2015;Zhang 等;2014;Ali和Ortiz,2016;Kim 等,2016;Lobo 等,2015;Hicks 等,2013)和神经网络模型(Giardino 等,2010;Matthews 等,2010) 或者基于辐射传输机理构建生物光学模型(如查找表法,矩阵反演算法)(Huang 等,2014;Giardino 等,2015;Dekker 等,2011;张兵 等,2009)来实现水质参数的动态监测。由于生物光学模型的复杂性,简单有效的经验模型成为总悬浮物和浊度遥感监测的常用模型。如Shi等(2015)基于2003年—2013年的MODIS数据在 545 nm处的遥感反射率和实测太湖水体总悬浮物浓度之间的稳定关系构建了太湖水体总悬浮物浓度反演的经验模型,分析了太湖水体总悬浮浓度的时空变异性;Lobo等(2015)利用1973年—2013年的Landsat MSS/TM/OLI影像获取的近红外波段反射率和实测矿坑水体总悬浮物浓度构建单波段指数模型,利用该模型对不同时间点矿坑水体总悬浮物浓度进行监测,在此基础上分析了矿坑水体总悬浮物浓度的时空变化规律及其驱动因素;Hicks等(2013)利用新西兰怀卡托区域部分湖泊实测的总悬浮物浓度、浊度和透明度数据和时序Landsat ETM+数据构建波段组合模型,对有观测的湖泊水体3种水质参数进行遥感监测,并将该模型应用于无观测的湖泊,弥补了无观测湖泊水质监测的空白;Ali和Ortiz(2016)利用400—900 nm范围内的全部波段反射率和总悬浮物浓度分别作为自变量和因变量,构建水体总悬浮物浓度偏最小二乘反演模型,取得较高的反演精度。由于内陆水体光学特性的复杂性,经验模型的时空移植性较差,因此,针对特定湖泊需要构建适用于该湖泊水体总悬浮物浓度和浊度监测的遥感反演模型。
由于很多草型或者混合型湖泊中生长有大量的水生植物,水生植物会造成“水体—水生植物”混合像元问题,导致卫星传感器获取的水生植物生长区域的光谱信息为水体和水生植物的混合光谱,直接利用经验和半经验模型反演水生植物生长区域水质参数存在不确定性(李俊生 等,2009;Giardino等,2015)。针对水生植物生长区域的水质反演,部分学者对水生植物生长区域不做特殊处理,直接利用水质遥感模型对水生植物生长区域水质进行反演,如Shi等(2015)和Zhang等(2014)利用遥感模型直接对东太湖水生植物生长区域水体总悬浮物浓度进行反演;还有部分学者利用目视解译(Birk和Ecke,2014)、光谱指数(如归一化植被指数和叶绿素a光谱指数)(Villa 等,2013;朱庆 等,2016)、监督或非监督分类方法(Hunter 等,2010;Oppelt 等,2012;Bolpagni 等,2014;Cao 等,2018)、生物光学模型和混合像元分解方法(Giardino 等,2015;Brooks 等,2015)将水生植物生长区域和水体区域分离,仅对水体区水质参数进行遥感监测(Giardino 等,2015;Bolpagni 等,2014;Cao 等,2018);此外,乔娜等(2016) 将南四湖分为水生植物覆盖区和非覆盖区,利用水生植物对水质的指示作用对水生植物覆盖区总悬浮物浓度定性反演,但未考虑水生植物类型和水生植物生长状况对水质的影响。不同物候期的水生植物对水体组分的迁移转化的影响机制不同,处于生长期的水生植物能够吸收湖泊沉积物间隙水中氮磷等营养盐,抑制浮游植物生长和藻类水华过程 (张云 等;2018),此外,处于生长期的沉水植物可以吸附、固着和沉降水体中的悬浮物,其根部可以有效抑制沉积物的再悬浮(Zhang 等,2014;Abo El-lil,2006),降低沉积物营养盐的释放,提高水体的透明度 (常素云 等,2016);但处于衰亡期的水生植物,由于降解过程的氮磷代谢会导致水体氮磷浓度升高,加上衰亡过程产生的植物残体和厌氧条件,导致水体透明度下降,水质不断恶化 (申秋实 等,2014;Cao 等,2018)。因此,如何实现不同物候期内水生植物生长水域的水质遥感监测对实现整个草型或混合型湖泊水质遥感监测至关重要。
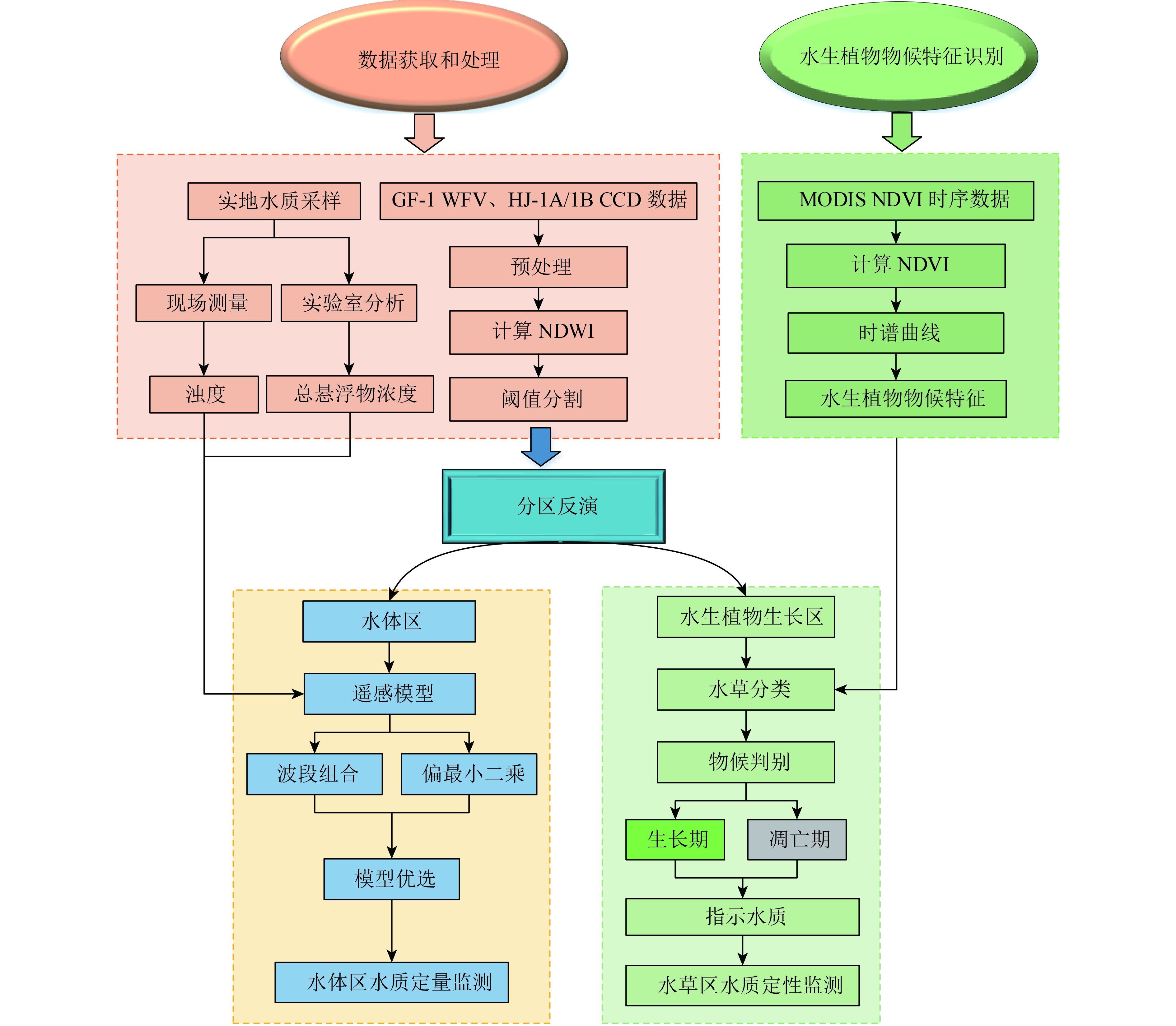
本文基于分区反演思路将草型湖泊微山湖区分为水生植物覆盖区和水体区,针对水生植物覆盖区,利用时序MODIS植被指数(NDVI)产品识别微山湖典型水生植物的物候特征,基于不同物候期内水生植物对微山湖水体总悬浮物浓度和浊度的指示作用,对微山湖水生植物覆盖区水体总悬浮物浓度和浊度进行定性遥感监测;针对于水体区,利用波段组合模型和偏最小二乘模型对微山湖水体总悬浮物浓度和浊度进行定量遥感监测。将定量和定性遥感监测方法相结合,对草型湖泊微山湖总悬浮物浓度和浊度进行遥感监测,分析了微山湖水体浮物浓度和浊度的时空变化规律。
2 材料和方法
2.1 研究区
微山湖地处116°58′E—117°21′E,34° 27′N—34°52′N,隶属于山东省济宁市微山县,与南阳湖、独山湖和昭阳湖一起称为南四湖,位于南四湖的最南端,是面积最大的一个湖泊,面积达660 km2。微山湖平均水深不足3 m,春夏湖泊中生长有各种水生植物,其中以光叶眼子菜、穗花狐尾藻和菹草为主,是典型的内陆浅水草型湖泊,此外,微山湖西南边界存在部分湿地和农田。
微山湖是中国北方最大的淡水湖,也是南水北调东线工程重要的湖泊水源地,同时具有防洪、灌溉、养殖和航运等生态系统服务功能,利用遥感手段动态监测微山湖水质对南水北调东线水资源监控管理具有重要意义。
2.2 数据获取和处理
2014年7月—2015年6月,在微山湖开展了6次野外试验,采用均匀布点原则在微山湖布置采样点。乘坐快艇到达预定位置,待水流平稳后利用采样瓶采集水样,对于水生植物生长茂密区域,采集表层水样(图1(a));对于水植物生长相对稀疏区域,在水生植物生长空隙处采集水面以下0.5m左右处的水样(图1(b));对于水体区域则就近采集水面以下0.5 m左右处的水样(图1(c)),水样用锡纸包裹,冷藏后送回实验室分析,利用称重法计算水体总悬浮物浓度。采集水样的同时,利用美国哈希(HACH)浊度仪1900C现场测定水体浊度。具体采样时间、采样点个数、总悬浮物浓度和浊度统计结果如表1所示。由表1可以看出,微山湖水体总悬浮物浓度和浊度具有显著的时空变异性,不同月份微山湖水体总悬浮物浓度和浊度最大值、最小值、均值和标准存在显著差异,其中2014年7月微山湖水体最为浑浊,总悬浮物浓度和浊度4项统计指标均显著大于其他月份;而2015年4月微山湖水体最为清澈,水体浊度4项统计指标均小于其他月份。从均值指标来看,微山湖水体总悬浮物浓度和浊度从4月—7月呈逐渐增加的趋势,随后逐渐降低。


表 1 微山湖水体总悬浮物浓度和浊度统计
Table 1 Statistics of total suspended matter and turbidity in Weishan Lake
| 试验时间 | 样点数N | TSM/(mg/L) | Turbidity/(NTU) | 卫星遥感数据源 | ||||||||||
| Max | Min | Mean | Std | Max | Min | Mean | Std | HJ-1A/1B | GF-1 | |||||
| 2014-07-21—23 | 17 | 352.0 | 12.0 | 80.5 | 95.1 | 343.0 | 13.7 | 77.2 | 98.8 | — | 2014-07-21 | |||
| 2014-08-29 | 11 | 76.0 | 3.0 | 42.7 | 23.2 | 78.7 | 3.0 | 40.1 | 24.4 | 2014-08-29 | — | |||
| 2014-11-17 | 13 | 42.0 | 5.0 | 18.1 | 9.1 | 39.9 | 3.8 | 17.2 | 8.5 | — | 2014-11-14 | |||
| 2015-04-06—09 | 31 | — | — | — | — | 30.4 | 1.5 | 9.0 | 6.1 | 2015-04-10 | ||||
| 2015-05-24 | 29 | 36.9 | 2.0 | 14.3 | 10.2 | 40.7 | 2.3 | 15.9 | 11.8 | — | 2015-05-25 | |||
| 2015-06-11—13 | 41 | 141.0 | 1.0 | 35.9 | 35.3 | 140.0 | 2.1 | 44.5 | 41.1 | — | 2015-06-06 | |||
| 注:—代表无有效数据。 | ||||||||||||||
从中国资源卫星应用中心网站(http://www.cresda.com/CN/[2018-03-26]) 上下载和采样时间同步或准同步的HJ-1A/1B CCD和GF-1 WFV数据,影像获取情况和拍摄时间见表1。利用ENVI 5.0软件对获取的HJ-1A/1B CCD和GF-1WFV影像进行辐射定标、几何精校正和大气校正处理,获取微山湖水体遥感反射率;辐射定标利用中国资源卫星应用中心提供的定标系数,将影像中的DN值转换为辐亮度;几何精校正以Landsat 8影像作为基准影像,对HJ-1A/1B CCD和GF-1WFV影像的几何形变进行纠正;大气校正采用的是ENVI 5.0软件中FLAASH模块,FLAASH采用MODTRAN 4辐射传输模型,该模型是目前世界上发展较为完善的大气校正方法之一,可以快速有效地获取水体遥感反射率(朱云芳 等,2017)。此外在MODIS WEB网站上(https://modis.gsfc.nasa.gov/[2018-03-26])下载时序MOD13A1植被指数(NDVI)产品。
2.3 草型湖泊水体总悬浮物浓度和浊度遥感监测方法
2.3.1 总体研究思路
基于分区思想,利用NDWI将微山湖区分为水生植物覆盖区和水体区;针对水生植物覆盖区,考虑水生植物的物候特征,利用水生植物对微山湖水体总悬浮物浓度和浊度的指示作用对微山湖水生植物覆盖区水体总悬浮物浓度和浊度进行定性监测;针对水体区,构建反演微山湖水体区总悬浮物浓度和浊度的经验模型,对微山湖水体区总悬浮物浓度和浊度进行定量反演。具体思路如图2所示。
2.3.2 水生植物候特征识别
不同水生植物生长和凋亡的时间节点不同,即水生植物存在不同的物候特征(Hestir 等,2015)。水生植物的生长周期可利用时序NDVI构建时谱曲线进行表征(李瑶 等,2016)。选择500m空间分辨率和16天时间分辨率的MODIS NDVI产品(MOD13A1)作为数据源,每年可以获取23个时相的MODIS NDVI数据,利用微山湖矢量数据对2014年—2015年覆盖微山湖区域的MODIS NDVI产品进行裁剪,将不同时相的NDVI数据按时间顺序进行叠加,建立微山湖区NDVI时谱曲线。每一景NDVI图像相当于时谱曲线的一个“波段”。不同地物的时谱曲线不同,从微山湖区时谱曲线中提取典型地物的时谱曲线。
2.3.3 水生植物遥感监测
归一化水体指数NDWI(Normalized Difference Water Index)是提取水体的重要指数(Hou 等,2017),可有效区分水体和水生植物。基于大气校正后的多光谱影像,计算微山湖区NDWI值。由于水生植物会增加对近红外波段的反射率,在假彩色影像(RGB:近红、红、绿) 中往往呈暗红色,因此结合目视解译,设定阈值将影像中呈暗红色区域提取出来。
2.3.4 水体区总悬浮物浓度和浊度遥感模型构建
基于多光谱数据的水体总悬浮物浓度和浊度遥感监测模型以波段组合模型(Bonansea 等,2015;Hicks 等,2013)为主,该模型以多光谱波段组合和对应的总悬浮物浓度或浊度之间的相关分析为基础,选择和总悬浮物浓度或浊度相关系数最高的波段组合和水质参数建立统计回归模型。此外,偏最小二乘模型作为一种多元回归模型,可以同时利用多个波段信息构建水质参数的多元回归反演模型,目前多应用于高光谱水质遥感(刘忠华 等,2011;Song 等,2013;Cao 等,2018)。本研究中,偏最小二乘模型以GF-1 WFV和HJ-1A/1B CCD的4个波段作为自变量,总悬浮物浓度和浊度作为因变量,建模详细步骤参考刘忠华等(2011)论文。
3 结果与讨论
3.1 水生植物物候特征识别
利用2014年7月—2015年6月的MODIS植被指数时序数据获取的微山湖区光叶眼子菜、菹草、水体、农田和湿地的时谱曲线如图3所示,其中穗花狐尾藻和光叶眼子菜的时谱曲线类似,将两者归为一类。由图3可以看出,不同地物时谱曲线存在显著差异(李瑶 等,2016),说明不同地物具有不同的物候特征,其中,农田的时谱曲线具有双峰特征,夏季农作物的收割导致农田的时谱曲线在夏季存在一个低谷;湿地的NDVI从春季开始上升,到夏季末达到峰值,然后开始下降;光叶眼子菜和穗花狐尾藻的NDVI变化趋势和湿地的一致,也是从春季开始上升,到夏季末达到峰值,然后开始下降,但是NDVI的数值较小;菹草的NDVI从初春开始上升,到春末的时候达到峰值,然后急速下降,此后一直保持较低的状态;水体的NDVI一直保持较低的状态,且变化不大。

图4(a)—(k)为不同物候期内光叶眼子菜、穗花狐尾藻和菹草生长照片。由图4(a)—(h)可以看出光叶眼子菜和穗花狐尾藻在4月开始生长,分布零散,在5月—7月期间内逐渐生长茂盛,成片的光叶眼子菜和穗花狐尾藻贴近甚至浮于水面,此后逐渐衰亡。由图4(i)—(k)可以看出菹草在4月开始迅速生长,成片的菹草逐渐贴近水面,到5月末开始,大片菹草贴近甚至浮于水面,此时水草进入凋亡期,到6月中旬,大片菹草迅速腐烂凋亡。利用NDVI获取的光叶眼子菜/穗花狐尾藻和菹草的物候特征和实地采样中观测到的光叶眼子菜/穗花狐尾藻和菹草的物候特征保持一致。

3.2 微山湖水生植物时空变化监测
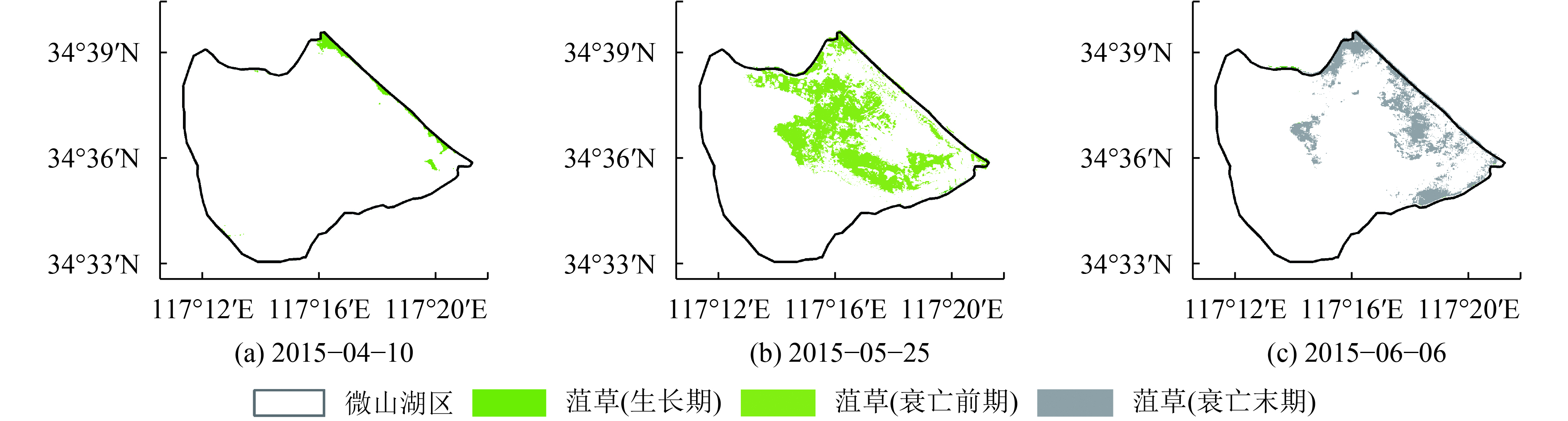
利用时序GF-1 WFV数据和HJ-1A/1B CCD数据计算微山湖区水体归一化水体指数,基于实地采样获取的水生植物分布,利用目视解译设定阈值,结合不同水生植物的物候特征得到2014年7月21日、2014年8月29日、2014年11月14日、2015年4月10日、2015年5月25日和2015年6月6日微山湖区光叶眼子菜/穗花狐尾藻(图5)和菹草的时空变化图(图6)。由图5可以看出,2014年7月21日微山湖水生植物主要集中于西南湖区,此时光叶眼子菜和穗花狐尾藻长势茂盛,成片的贴于水面(图4(d));到2014年8月底,光叶眼子菜和穗花狐尾藻逐渐开始进入衰亡期,但衰亡十分缓慢,直至2014年11月份才逐渐衰亡沉入水底(图5(b)—(d));到2015年5月底时,微山湖区西南角的光叶眼子菜和穗花狐尾藻开始进入生长期,部分光叶眼子菜和穗花狐尾藻长势茂盛,开始贴近水面(图5(e)),到6月份时,贴近水面的光叶眼子菜和穗花狐尾藻面积进一步扩大(图5(f)),和图3显示的光叶眼子菜和穗花狐尾藻的物候特征保持一致。由图6可以看出,2015年4月10日,微山湖东北湖边附近的菹草迅速生长,逐渐贴近水面(图4(i)),到2015年5月份时,菹草生长区域基本覆盖了整个微山湖东北湖区,由于菹草的生长周期较短(图3),到2015年5月底时,菹草进入衰亡期;菹草在6月份的时候开始迅速衰亡降解,湖山湖东北湖区的菹草覆盖面积迅速减少,部分菹草残体浮于水面(图4(k))。微山湖区水生植物具有明显的时空变化规律,菹草主要生长于微山湖东北湖区,在4月份开始迅速生长,然后至6月份迅速衰亡腐烂;光叶眼子菜和穗花狐尾藻主要生长在西南湖区,从5月份开始进入生长期,到7月份基本遍布整个西南湖区,随后逐渐进入衰亡期,直至11月份才逐渐衰亡降解完全。

3.3 水生植物对水质的指示作用
处于生长期的水生植物能够吸收沉积物和水体中的营养盐(黄亮 等,2010),和浮游植物形成竞争关系,同时会分泌抑制悬浮物生长的化感物质 (Wu 等,2009),可以有效抑制藻类生长;沉水植物枝叶对水体悬浮泥沙具有吸附、固着和沉降作用(Abo El-lil,2006),通过改变水体上下水动力条件,沉水植物能够减小水体挟沙能力,且植物表面的分泌物能促使悬浮泥沙颗粒由分散的悬移质向絮凝团转化,当浮力小于重力时下降,悬浮泥沙将沉积于植物根部周围(郭长城 等,2007),此外沉水植物根部可以固着底泥,防止底泥再悬浮(Zhang 等,2014);此外沉水植物光合作用增加了水中的溶解氧,可以促进微生物对污染物的分解作用(Srivastava 等,2017)。因此,处于生长期的水生植物区域水质一般较好,水体透明度较高(秦伯强,2002)。微山湖中生长的光叶眼子菜、穗花狐尾藻和菹草均属于沉水植物,其中光叶眼子菜具有椭圆形的大叶片,茎粗壮;穗花狐尾藻具有穗状花序,生长茂密的的穗花狐尾藻可以浮于水体表面;菹草叶片呈针形,先端钝圆,叶片边缘具有锯齿。沉水植物一般整个植株都处于水中,在生长高峰期可以接近或者浮于水体表面,表2和表3分别为微山湖区处于不同物候期的菹草、光叶眼子菜和穗花狐尾藻分布区域水体总悬浮物浓度和浊度的统计结果。可以看出处于生长期的3种沉水植物分布区域水体总悬浮物浓度和浊度均值均小于10 mg/L(NTU),标准差均较小,处于生长期的3种水生植物分布区域水质整体较好。
表 2 微山湖区菹草生长区域水体总悬浮物浓度和浊度的统计结果
Table 2 Statistics of total suspended matter concentration and turbidity in Potamogeton crispus growth district of Weishan Lake
| 2015-04-06—09 | 2015-05-24 | 2015-06-11—13 | ||||||||
| TSM/(mg/L) | 浊度/(NTU) | TSM/(mg/L) | 浊度/(NTU) | TSM/(mg/L) | 浊度/(NTU) | |||||
| 最大值 | 12.0 | 13.1 | 20.0 | 23.3 | 141.1 | 140.0 | ||||
| 最小值 | 4.0 | 4.6 | 3.2 | 2.7 | 19.0 | 34.5 | ||||
| 均值 | 7.5 | 9.1 | 8.9 | 8.9 | 65.0 | 84.3 | ||||
| 标准差 | 2.2 | 2.3 | 6.5 | 7.8 | 31.3 | 31.8 | ||||
| 采样点个数 | 8 | 9 | 16 | |||||||
表 3 微山湖区光叶眼子菜和穗花狐尾藻生长区域水体总悬浮物浓度和浊度的统计结果
Table 3 Statistics of total suspended matter concentration and turbidity in Potamogeton lucens and Myriophyllum spicatum growth district of Weishan Lake
| 光叶眼子菜/穗花狐尾藻 | 光叶眼子菜 | 穗花狐尾藻 | ||||||||
| TSM/(mg/L) | 浊度/(NTU) | TSM/(mg/L) | 浊度/(NTU) | TSM/(mg/L) | 浊度/(NTU) | |||||
| 最大值 | 19.0 | 23.6 | 6.9 | 12.8 | 13.1 | 26.30 | ||||
| 最小值 | 5.0 | 3.8 | 2.0 | 2.1 | 1.1 | 4.3 | ||||
| 均值 | 12.6 | 12.2 | 3.8 | 6.4 | 6.6 | 9.6 | ||||
| 标准差 | 5.0 | 6.6 | 1.6 | 3.4 | 3.7 | 7.1 | ||||
| 样点数 | 5 | 6 | 6 | |||||||
| 采样时间 | 2014-11-17 | 2015-06-11—13 | 2015-06-11—13 | |||||||
由图3可以看出,菹草的时谱曲线在6月初急剧下降,菹草短在时间内迅速衰亡腐烂(图4(k)),菹草在生长期内吸收的营养盐重新释放进入水体(王锦旗 等,2011),造成严重的内源污染,此外水生植物衰亡过程产生的植物残体(图4(k)),导致水体悬浮物浓度上升,水体十分浑浊,水质短时间内迅速恶化(申秋实 等,2014;Cao 等,2018)。由表2可以看出,2015年4月06—09日微山湖菹草生长区域水体总悬浮物浓度和浊度均小于15 mg/L(NTU),2015年5月24日菹草分布区域水体总悬浮物浓度和浊度均值同样处于较低水平,但标准差较2015年4月份有所增加。根据菹草的物候特征可知,2015年5月24日菹草已经开始衰亡,结合实地采样观察,此时菹草衰亡程度较低,菹草分布区域水质依旧较好,该区域总悬浮物浓度和浊度分别低于20 mg/L和25 NTU;2015年6月11—13日微山湖区菹草分布区域水质显著变差,总悬浮物浓度和浊度均值均高于60 mg/L(NTU),总悬浮物浓度和浊度最大值达到140 mg/L(NTU)(表2),此时微山湖区的菹草基本全部腐烂,部分残体浮于水面 (图4(k))。相比于菹草,光叶眼子菜和穗花狐尾藻的生长衰亡周期较长(图3),结合实地采样,发现2014年11月17日微山湖区光叶眼子菜和穗花狐尾藻已经腐烂沉降,此时微山湖西南湖区总悬浮物浓度和浊度均值均低于30 mg/L(NTU),水质无明显恶化(表3)。处于生长期的光叶眼子菜和穗花狐尾藻生长区域水体总悬浮物浓度和浊度范围分别为0—15 mg/L和0—30 NTU,水质整体较好。
微山湖区光叶眼子菜、穗花狐尾藻和菹草在不同生长周期内对水质具有不同的指示作用。因此,基于不同水生植物的物候特征,可利用沉水植物对水质的指示作用对水生植物生长区域水质进行定性监测。其中处于生长期的光叶眼子菜和穗花狐尾藻分布区域水体总悬浮物浓度和浊度为0—15 mg/L和0—30 NTU,处于生长期的菹草分布区域水体总悬浮物浓度和浊度分别为0—15 mg/L(NTU);处于衰亡前期的菹草分布区域水体总悬浮物浓度和浊度分别为0—20 mg/L和0—25 NTU;处于衰亡末期的菹草分布区域水体总悬浮物浓度和浊度分别为15—145 mg/L和30—140 NTU。由于缺乏处于衰亡期的光叶眼子菜和穗花狐尾藻分布区域水体总悬浮物浓度和浊度,处于衰亡期的光叶眼子菜和穗花狐尾藻对水质的指示作用需要后续研究。本研究中水生植物对水质的指示作用适用于水生植物分布区域,不包括水生植物分布边界以外水域,此外,由于大风或强降雨等恶劣天气条件会造成湖泊底泥的再悬浮,此时水体会变得浑浊,水生植物对悬浮颗粒的吸附和沉降需要一定的时间,所以恶劣天气过后短时间内不适合利用水生植物盖度的大小来指示总悬浮物浓度和浊度(乔娜 等,2016)。
水生植物和水质的作用是相互的,沉水植物对水质具有净化作用,恶劣的水质同样影响着沉水植物的生长和分布(Shields和Moore,2016)。富营养化浅水湖泊中水生植物会逐渐减少或消失(Scheffer 等,2003),导致大面积的蓝藻水华爆发。因此,水生植物可以作为湖泊水环境水质状况的指示物。
3.4 微山湖水质时空变化分析
针对水生植物覆盖区,利用不同生长期内不同水草对水质的指示作用对微山湖水生植物覆盖区水体总悬浮物浓度和浊度进行定性反演;针对水体区,基于2014年7月—2015年6月在微山湖区6次试验获取的微山湖区水体总悬浮物浓度、浊度和准同步的GF-1和HJ-1A/1B多光谱影像,分别构建水体区总悬浮物浓度和浊度单波段/波段组合模型和偏最小二乘反演模型,不同模型决定系数如表4所示。选择决定系数最高的单波段/波段组合模型或偏最小二乘模型对微山湖水体区总悬浮物浓度和浊度进行定量反演。2014年7月—2015年6月微山湖水生植物覆盖区总悬浮物浓度的定性监测结果和水体区总悬浮物浓度的定量监测结果如图7和图8所示,其中2015年4月总悬浮物浓度测量数据无效,共5组反演结果;2014年7月—2015年6月微山湖水草区浊度的定性监测结果和水体区浊度的定量监测结果如图9和图10所示。由于缺少处于衰亡期的光叶眼子菜和穗花狐尾藻对水体总悬浮物浓度和浊度的具体指示作用,未对2014年8月29日光叶眼子菜和穗花狐尾藻覆盖区水体总悬浮物浓度和浊度进行定性监测。
表 4 微山湖水体区总悬浮物浓度和浊度反演模型
Table 4 Retrieval models of total suspended matter concentration and turbidity in water district of Weishan Lake
| 2014-07-21 | 2014-08-29 | 2014-11-17 | 2015-04-10 | 2015-05-25 | 2015-06-06 | ||||||||||||
| B3/B4e | PLS | B2l | PLS | B3/B11 | PLS | — | B3/B11 | PLS | B3e | PLS | |||||||
| 总悬浮浓度 | 0.60 | 0.63 | 0.76 | 0.73 | 0.69 | 0.72 | — | 0.74 | 0.83 | 0.83 | 0.67 | ||||||
| 浊度 | 0.60 | 0.84 | 0.90 | 0.91 | 0.72 | 0.67 | 0.68 | 0.64 | 0.75 | 0.79 | 0.79 | 0.63 | |||||
| 注:e代表指数拟合,l代表线性拟合,粗体代表最终采用的模型。 | |||||||||||||||||




由图7和图8可以看出,微山湖水体总悬浮物浓度具有显著的时空变异性。如图7(a)所示,2014年7月21日,位于微山湖区东北方向的菹草全部腐烂,生长期内吸收的大量营养盐集中释放,同时生成用于繁殖的鳞枝(张敏 等,2015),导致微山湖水体总悬浮物浓度显著增高,由定量监测结果可以看出微山湖区东北方向水体总悬浮物浓度普遍高于100 mg/L(图8(a)),而西南湖区水体总悬浮物浓度整体较低,总悬浮物浓度大都低于20 mg/L (图7(a)),这是因为此时该区域长有光叶眼子菜和穗花狐尾藻等沉水植物,沉水植物对悬浮物具有吸附和沉降作用(Abo El-lil,2006),同时抑制因风浪等原因造成的底泥的再悬浮(Zhang 等,2014),降低了沉水植物生长区域水体总悬浮物浓度;如图8(b),2014年8月29日,微山湖东北湖区水体总悬浮物较2014年7月21日有明显的降低,菹草短期内迅速降解造成的悬浮物浓度上升的持续时间较短,随着时间的推移,菹草衰亡产生的鳞枝和悬浮颗粒最终沉降至湖底(张敏 等,2015),总悬浮物浓度逐渐降低;由图8(c)可以看出,2014年11月14日微山湖区基本已无水草,加上悬浮物的沉降作用,水体总悬浮物浓度处于较低水平;图7(b)可以看出,2015年4月10日微山湖东北边界处菹草开始快速生长,到2015年5月25日时,微山湖整个东北湖区均被大量的菹草覆盖,西南湖区中的光叶眼子菜和穗花狐尾藻也开始生长,整个湖区的悬浮物浓度均处于较低水平(图7(c)和图8(d);由于菹草的生长周期较短,到6月初,微山湖区菹草迅速衰亡降解,导致微山湖东北区域总悬浮物浓度急剧上升,水质短时间内迅速恶化,而此时微山湖西南湖区长有大量的光叶眼子菜和穗花狐尾藻,总悬浮物浓度均处于较低水平(图7(d)和图8(e))。
对比图7和图9以及图8和图10,可以看出微山湖水体浊度的时空变化规律和总悬浮物浓度变化规律具有一致性,这是因为微山湖水体总悬浮物浓度和浊度具有显著的相关性,相关系数达0.9以上,微山湖水体浊度主要由总悬浮浓度主导(Cao 等,2018)。微山湖水体总悬浮物浓度和浊度具有明显的时空变化规律,西南湖区生长着具有较长生长周期的光叶眼子菜和穗花狐尾藻,该区域水体总悬浮物浓度和浊度长期保持着较低水平,而东北湖区在6月之前长有大量的菹草,水质整体较好,但菹草在6月初会迅速衰亡降低,短期内产生大量的植物残体,导致该区域水质急剧恶化,随后菹草的降解完全,东北湖区的悬浮物浓度会逐渐降低。
4 结 论
本文针对草型湖泊水生植物覆盖区域水质难以直接利用遥感监测的问题,提出考虑水生植物物候特征的草型湖泊水质监测方法,识别了2014年7月—2015年6月微山湖水体水生植物和水质的时空变化规律,得到如下主要结论:
(1) 微山湖区长有大量水生植物,其中以光叶眼子菜、穗花狐尾藻和菹草等沉水植物为主,微山湖区光叶眼子菜、穗花狐尾藻和菹草具有显著时空分布规律。时间上,3种水生植物的物候特征存在差异,其中菹草的生长周期较短,从初春开始迅速生长,到春末的时候达到峰值,随后迅速衰亡降解,而光叶眼子菜和穗花狐尾藻的生长周期较长,从5月份开始生长,到7月中旬达到峰值,随后慢慢衰亡,直至11月份降解完全;空间上,菹草主要分布于微山湖东北湖区,而光叶眼子菜和穗花狐尾藻主要分布于微山湖西南湖区。
(2) 不同水生植物在不同生长周期内对水质具有不同的指示作用。处于生长期内光叶眼子菜、穗花狐尾藻和菹草分布区域水质整体较好,悬浮物浓度和浊度基于低于15 mg/L和30 NTU,但菹草从6月份开始迅速衰亡降解,导致水体迅速恶化,总悬浮物浓度和浊度分布范围分别为15—145 mg/L和30—140 NTU。
(3) 微山湖总悬浮物浓度和浊度具有明显的时空分布规律。2014年7月,随着菹草的凋亡降解,微山湖东北湖区水体总悬浮物浓度和浊度显著升高,随着时间的推移,东北湖区水体总悬浮物浓度和浊度逐渐降低;2015年4月—5月,由于东北湖区长有大量的菹草,整个东北湖区水质较好,但到6月份时,由于菹草的迅速凋亡,东北湖区的总悬浮物浓度和浊度逐渐上升。而西南湖区长有生长周期较长的光叶眼子菜和穗花狐尾藻,水体总悬浮物浓度和浊度一直保持较低水平。
(4) 基于不同水生植物的物候特征,利用水生植物对水质的指示作用可以实现草型湖泊水生植物覆盖区域水质的定性监测,结合水质遥感模型对水体区水质的定量监测,可以实现整个草型湖泊水质的遥感监测。但受采样次数的限制,对不同水生植物在不同物候期内对水质的指示作用研究的还不够深入,后续将继续积累数据进行进一步的研究。
志 谢 野外实验数据获取得到了济南大学桑国庆副教授和中国科学院遥感与数字地球研究所黄长平副研究员的大力支持,此外张立福研究员、田庆久教授、陈子丹教授对草型湖泊水生植物覆盖区水质遥感监测提出了宝贵建议,在此表示衷心的感谢!
参考文献(References)
-
Abo El-lil A H. 2006. Evaluation of the efficiency of some hydrophytes for trapping suspended matters from different aquatic ecosystems. Biotechnology, 5 (1): 90–97. [DOI: 10.3923/biotech.2006.90.97]
-
Ali K A and Ortiz J D. 2016. Multivariate approach for chlorophyll-a and suspended matter retrievals in Case II type waters using hyperspectral data. Hydrological Sciences Journal, 61 (1): 200–213. [DOI: 10.1080/02626667.2014.964242]
-
Birk S and Ecke F. 2014. The potential of remote sensing in ecological status assessment of coloured lakes using aquatic plants. Ecological Indicators, 46 : 398–406. [DOI: 10.1016/j.ecolind.2014.06.035]
-
Bolpagni R, Bresciani M, Laini A, Pinardi M, Matta E, Ampe E M, Giardino C, Viaroli P and Bartoli M. 2014. Remote sensing of phytoplankton-macrophyte coexistence in shallow hypereutrophic fluvial lakes. Hydrobiologia, 737 (1): 67–76. [DOI: 10.1007/s10750-013-1800-6]
-
Bonansea M, Rodriguez M C, Pinotti L and Ferrero S. 2015. Using multi-temporal Landsat imagery and linear mixed models for assessing water quality parameters in Río Tercero reservoir (Argentina). Remote Sensing of Environment, 158 : 28–41. [DOI: 10.1016/j.rse.2014.10.032]
-
Brooks C, Grimm A, Shuchman R, Sayers M and Jessee N. 2015. A satellite-based multi-temporal assessment of the extent of nuisance Cladophora and related submerged aquatic vegetation for the Laurentian Great Lakes . Remote Sensing of Environment, 157 : 58–71. [DOI: 10.1016/j.rse.2014.04.032]
-
Cao Y, Ye Y T, Zhao H L, Jiang Y Z, Wang H, Shang Y Z and Wang J F. 2018. Remote sensing of water quality based on HJ-1A HSI imagery with modified discrete binary particle swarm optimization-partial least squares (MDBPSO-PLS) in inland waters: A case in Weishan Lake. Ecological Informatics, 44 : 21–32. [DOI: 10.1016/j.ecoinf.2018.01.004]
-
Chang S Y, Wu T and Zhao J J. 2016. Effect of planting different submerged macrophytes groups on water purification of Beidagang Reservoir. Chinese Journal of Environmental Engineering, 10 (1): 439–444. [DOI: 10.12030/j.cjee.20160173] ( 常素云, 吴涛, 赵静静. 2016. 不同沉水植物组配对北大港水库水体净化效果的影响. 环境工程学报, 10 (1): 439–444. [DOI: 10.12030/j.cjee.20160173] )
-
Dekker A G, Phinn S R, Anstee J, Bissett P, Brando V E, Casey B, Fearns P, Hedley J, Klonowski W, Lee Z P, Lynch M, Lyons M, Mobley C and Roelfsema C. 2011. Intercomparison of shallow water bathymetry, hydro-optics, and benthos mapping techniques in Australian and Caribbean coastal environments. Limnology and Oceanography: Methods, 9 (9): 396–425. [DOI: 10.4319/lom.2011.9.396]
-
Dörnhöfer K and Oppelt N. 2016. Remote sensing for lake research and monitoring - Recent advances. Ecological Indicators, 64 : 105–122. [DOI: 10.1016/j.ecolind.2015.12.009]
-
Duan H T, Ma R H, Xu X F, Kong F X, Zhang S H, Kong W J, Hao J Y and Shang L L. 2009. Two-decade reconstruction of algal blooms in China's Lake Taihu. Environmental Science and Technology, 43 (10): 3522–3528. [DOI: 10.1021/es8031852]
-
Giardino C, Bresciani M, Valentini E, Gasperini L, Bolpagni R and Brando V E. 2015. Airborne hyperspectral data to assess suspended particulate matter and aquatic vegetation in a shallow and turbid lake. Remote Sensing of Environment, 157 : 48–57. [DOI: 10.1016/j.rse.2014.04.034]
-
Giardino C, Bresciani M, Villa P and Martinelli A. 2010. Application of remote sensing in water resource management: the case study of Lake Trasimeno, Italy. Water Resources Management, 24 (14): 3885–3899. [DOI: 10.1007/s11269-010-9639-3]
-
Guo C C, Yu G H and Wang G X. 2007. Removal of Potamogeton crispus to suspended sediment and N,P of water . 2007. Journal of Soil and Water Conservation, 21 (3): 108–111, 117. [DOI: 10.3321/j.issn:1009-2242.2007.03.025] ( 郭长城, 喻国华, 王国祥. 2007. 菹草对水体悬浮泥沙及氮、磷污染物的净化. 水土保持学报, 21 (3): 108–111, 117. [DOI: 10.3321/j.issn:1009-2242.2007.03.025] )
-
Güttler F N, Niculescua S and Gohinb F. 2013. Turbidity retrieval and monitoring of Danube Delta waters using multi-sensor optical remote sensing data: An integrated view from the delta plain lakes to the western–northwestern Black Sea coastal zone. Remote Sensing of Environment, 132 : 86–101. [DOI: 10.1016/j.rse.2013.01.009]
-
Hestir E L, Brando V E, Bresciani M, Giardino C, Matta E, Villa P and Dekker A G. 2015. Measuring freshwater aquatic ecosystems: The need for a hyperspectral global mapping satellite mission. Remote Sensing of Environment, 167 : 181–195. [DOI: 10.1016/j.rse.2015.05.023]
-
Hicks B J, Stichbury G A, Brabyn L K, Allan M G and Ashraf S. 2013. Hindcasting water clarity from Landsat satellite images of unmonitored shallow lakes in the Waikato region, New Zealand. Environmental Monitoring and Assessment, 185 (9): 7245–7261. [DOI: 10.1007/s10661-013-3098-2]
-
Hou X J, Feng L, Duan H T, Chen X L, Sun D Y and Shi K. 2017. Fifteen-year monitoring of the turbidity dynamics in large lakes and reservoirs in the middle and lower basin of the Yangtze River, China. Remote Sensing of Environment, 190 : 107–121. [DOI: 10.1016/j.rse.2016.12.006]
-
Huang C C, Li Y M, Yang H, Li J S, Chen X, Sun D Y, Le C F, Zou J and Xu L J. 2014. Assessment of water constituents in highly turbid productive water by optimization bio-optical retrieval model after optical classification. Journal of Hydrology, 519 : 1572–1583. [DOI: 10.1016/j.jhydrol.2014.09.007]
-
Huang L, Wu N C, Tang T, Li D F and Cai Q H. 2010. Enrichment and removal of nutrients in eutrophic water by aquatic macrophytes. China Environmental Science, 30 (S1): 1–6. ( 黄亮, 吴乃成, 唐涛, 黎道丰, 蔡庆华. 2010. 水生植物对富营养化水系统中氮、磷的富集与转移. 中国环境科学, 30 (S1): 1–6. )
-
Hunter P D, Gilvear D J, Tyler A N, Willby N J and Kelly A. 2010. Mapping macrophytic vegetation in shallow lakes using the Compact Airborne Spectrographic Imager (CASI). Aquatic Conservation: Marine and Freshwater Ecosystems, 20 (7): 717–727. [DOI: 10.1002/aqc.1144]
-
Jiang Y Z, Ye Y T and Wang H. 2010. Discussion on intelligent regulation technology architecture for river basin based on internet of things. Water Resources Informatization (4): 1–5, 10. [DOI: 10.3969/j.issn.1674-9405.2010.05.001] ( 蒋云钟, 冶运涛, 王浩. 2010. 基于物联网理念的流域智能调度技术体系刍议. 水利信息化 (4): 1–5, 10. [DOI: 10.3969/j.issn.1674-9405.2010.05.001] )
-
Jiang Y Z, Ye Y T and Wang H. 2011. Smart basin and its prospects for application. Systems Engineering—Theory and Practice, 31 (6): 1174–1181. [DOI: 10.12011/1000-6788(2011)6-1174] ( 蒋云钟, 冶运涛, 王浩. 2011. 智慧流域及其应用前景. 系统工程理论与实践, 31 (6): 1174–1181. [DOI: 10.12011/1000-6788(2011)6-1174] )
-
Jiang Y Z, Ye Y T and Wang H. 2014. Intelligent control and emergency treatment system of water quality and quantity for the interconnected river system network based on the internet of things. Systems Engineering—Theory and Practice, 34 (7): 1895–1903. [DOI: 10.12011/1000-6788(2014)7-1895] ( 蒋云钟, 冶运涛, 王浩. 2014. 基于物联网的河湖水系连通水质水量智能调控及应急处置系统研究. 系统工程理论与实践, 34 (7): 1895–1903. [DOI: 10.12011/1000-6788(2014)7-1895] )
-
Kim W, Moon J E, Park Y and Ishizaka J. 2016. Evaluation of chlorophyll retrievals from Geostationary Ocean Color Imager (GOCI) for the North-East Asian region. Remote Sensing of Environment, 184 : 482–495. [DOI: 10.1016/j.rse.2016.07.031]
-
Li J S, Wu D, Wu Y F, Liu H X, Shen Q and Zhang H. 2009. Identification of algae-bloom and aquatic macrophytes in Lake Taihu from in-situ measured spectra data. Journal of Lake Sciences, 21 (2): 215–222. [DOI: 10.18307/2009.0209] ( 李俊生, 吴迪, 吴远峰, 刘海霞, 申茜, 张浩. 2009. 基于实测光谱数据的太湖水华和水生高等植物识别. 湖泊科学, 21 (2): 215–222. [DOI: 10.18307/2009.0209] )
-
Li Y, Zhang L F, Huang C P, Wang J N and Cen Y. 2016. Monitor of cyanobacteria bloom in Lake Taihu from 2001 to 2013 based on MODIS temporal spectral data. Spectroscopy and Spectral Analysis, 36 (4): 1406–1411. [DOI: 10.3964/j.issn.1000-0593(2016)05-1406-06] ( 李瑶, 张立福, 黄长平, 王晋年, 岑奕. 2016. 基于MODIS植被指数时间谱的太湖2001年—2013年蓝藻爆发监测. 光谱学与光谱分析, 36 (4): 1406–1411. [DOI: 10.3964/j.issn.1000-0593(2016)05-1406-06] )
-
Liu Z H, Li Y M, Lü H, Xu Y F, Xu X, Huang J Z, Tan J and Guo Y L. 2011. Inversion of suspended matter concentration in Lake Chaohu based on partial least-squares regression. Journal of Lake Sciences, 23 (3): 357–365. [DOI: 10.18307/2011.0307] ( 刘忠华, 李云梅, 吕恒, 徐祎凡, 徐昕, 黄家柱, 檀静, 郭宇龙. 2011. 基于偏最小二乘法的巢湖悬浮物浓度反演. 湖泊科学, 23 (3): 357–365. [DOI: 10.18307/2011.0307] )
-
Lobo F L, Costa M P F and Novo E M L M. 2015. Time-series analysis of Landsat-MSS/TM/OLI images over Amazonian waters impacted by gold mining activities. Remote Sensing of Environment, 157 : 170–184. [DOI: 10.1016/j.rse.2014.04.030]
-
Matthews M W, Bernard S and Winter K. 2010. Remote sensing of cyanobacteria-dominant algal blooms and water quality parameters in Zeekoevlei, a small hypertrophic lake, using MERIS. Remote Sensing of Environment, 114 (9): 2070–2087. [DOI: 10.1016/j.rse.2010.04.013]
-
Oppelt N M, Schulze F, Doernhoefer K, Eisenhardt I and Bartsch I. 2012. Hyperspectral classification approaches for intertidal macroalgae habitat mapping: a case study in Heligoland. Optical Engineering, 51 (11): 111703 [DOI: 10.1117/1.OE.51.11.111703]
-
Panigrahi S, Wikner J, Panigrahy R C, Satapathy K K and Acharya B C. 2009. Variability of nutrients and phytoplankton biomass in a shallow brackish water ecosystem (Chilika Lagoon, India). Limnology, 10 (2): 73–85. [DOI: 10.1007/s10201-009-0262-z]
-
Qiao N, Huang C P, Zhang L F, Zhao H L, Ye Y T, Cen Y and Li Y. 2016. Remote sensing inversion of total suspended matter concentration in typical shallow macrophytic lake. Journal of Hubei University (Natural Science), 38 (6): 510–516. [DOI: 10.3969/j.issn.1000-2375.2016.06.007] ( 乔娜, 黄长平, 张立福, 赵红莉, 冶运涛, 岑奕, 李瑶. 2016. 典型浅水草型湖泊水体悬浮物浓度遥感反演. 湖北大学学报(自然科学版), 38 (6): 510–516. [DOI: 10.3969/j.issn.1000-2375.2016.06.007] )
-
Qin B Q. 2002. Approaches to mechanisms and control of eutrophication of shallow Lakes in the middle and lower reaches of the Yangze River. Journal of Lake Sciences, 14 (3): 193–202. [DOI: 10.18307/2002.0301] ( 秦伯强. 2002. 长江中下游浅水湖泊富营养化发生机制与控制途径初探. 湖泊科学, 14 (3): 193–202. [DOI: 10.18307/2002.0301] )
-
Scheffer M, Szabó S, Gragnani A, Van Nes E H, Rinaldi S, Kautsky N, Norberg J, Roijackers R M M and Franken R J M. 2003. Floating plant dominance as a stable state. Proceedings of the National Academy of Sciences of the United States of America, 100 (7): 4040–4045. [DOI: 10.1073/pnas.0737918100]
-
Shen Q S, Zhou Q L, Shao S G, Liu C, Zhang L and Fan C X. 2014. Estimation of in-situ sediment nutrients release at the submerged plant induced black bloom area in Lake Taihu . Journal of Lake Sciences, 26 (2): 177–184. [DOI: 10.18307/2014.0202] ( 申秋实, 周麒麟, 邵世光, 刘成, 张雷, 范成新. 2014. 太湖草源性" 湖泛”水域沉积物营养盐释放估算. 湖泊科学, 26 (2): 177–184. [DOI: 10.18307/2014.0202] )
-
Shi K, Zhang Y L, Zhu G W, Liu X H, Zhou Y Q, Xu X, Qin B Q, Liu G and Li Y M. 2015. Long-term remote monitoring of total suspended matter concentration in Lake Taihu using 250 m MODIS-Aqua data. Remote Sensing of Environment, 164 : 43–56. [DOI: 10.1016/j.rse.2015.02.029]
-
Shields E C and Moore K A. 2016. Effects of sediment and salinity on the growth and competitive abilities of three submersed macrophytes. Aquatic Botany, 132 : 24–29. [DOI: 10.1016/j.aquabot.2016.03.005]
-
Song K S, Li L, Tedesco L P, Li S, Duan H T, Liu D W, Hall B E, Du J, Li Z C, Shi K and Zhao Y. 2013. Remote estimation of chlorophyll-a in turbid inland waters: Three-band model versus GA-PLS model. Remote Sensing of Environment, 136 : 342–357. [DOI: 10.1016/j.rse.2013.05.017]
-
Srivastava J K, Chandra H, Kalra S J S, Mishra P, Khan H and Yadav P. 2017. Plant–microbe interaction in aquatic system and their role in the management of water quality: a review. Applied Water Science, 7 (3): 1079–1090. [DOI: 10.1007/s13201-016-0415-2]
-
Villa P, Laini A, Bresciani M and Bolpagni R. 2013. A remote sensing approach to monitor the conservation status of lacustrine Phragmites australis beds . Wetlands Ecology and Management, 21 (6): 399–416. [DOI: 10.1007/s11273-013-9311-9]
-
Wang J D, Sheng Y W and Tong T S D. 2014. Monitoring decadal lake dynamics across the Yangtze Basin downstream of Three Gorges Dam. Remote Sensing of Environment, 152 : 251–269. [DOI: 10.1016/j.rse.2014.06.004]
-
Wang J Q, Zheng Y F and Wang G X. 2011. Influences of Potamogeton crispus population on the lake water quality distribution . Environmental Science, 32 (2): 416–422. [DOI: 10.13227/j.hjkx.2011.02.032] ( 王锦旗, 郑有飞, 王国祥. 2011. 菹草种群对湖泊水质空间分布的影响. 环境科学, 32 (2): 416–422. [DOI: 10.13227/j.hjkx.2011.02.032] )
-
Wu Z B, Wang Y N, Liu B Y, Zhou Q H and Zhang Y Y. 2009. Allelopathic effects of phenolic compounds present in submerged macrophytes on Microcystis aeruginosa. Allelopathy Journal, 23 (2): 403–410.
-
Zhang B, Shen Q, Li J S, Zhang H and Wu D. 2009. Retrieval of three kinds of representative water quality parameters of Lake Taihu from hyperspectral remote sensing data. Journal of Lake Sciences, 21 (2): 182–192. [DOI: 10.18307/2009.0205] ( 张兵, 申茜, 李俊生, 张浩, 吴迪. 2009. 太湖水体3种典型水质参数的高光谱遥感反演. 湖泊科学, 21 (2): 182–192. [DOI: 10.18307/2009.0205] )
-
Zhang M, Yin C B, Zhang C Y, Wang L and Liu Z Q. 2015. Ecological function of submerged plant potamogeton crispus and its application status. Soil and Water Conservation in China (3): 50–53. [DOI: 10.3969/j.issn.1000-0941.2015.03.022] ( 张敏, 尹传宝, 张翠英, 王龙, 刘自强. 2015. 沉水植物菹草的生态功能及其应用现状. 中国水土保持 (3): 50–53. [DOI: 10.3969/j.issn.1000-0941.2015.03.022] )
-
Zhang Y, Wang S R, Duan C Q, Jiao L X, Wang Y R and Gao Q S. 2018. Spatial-temporal variations of nitrogen and phosphorus forms in sediment porewater as affected by submerged plant in Lake Dianchi. Journal of Lake Sciences, 30 (2): 314–325. [DOI: 10.18307/2018.0204] ( 张云, 王圣瑞, 段昌群, 焦立新, 王一茹, 高秋生. 2018. 滇池沉水植物生长过程对间隙水氮、磷时空变化的影响. 湖泊科学, 30 (2): 314–325. [DOI: 10.18307/2018.0204] )
-
Zhang Y L, Shi K, Liu X H, Zhou Y Q and Qin B Q. 2014. Lake topography and wind waves determining seasonal-spatial dynamics of total suspended matter in turbid Lake Taihu, China: Assessment using long-term high-resolution MERIS data. PLoS One, 9 (5): e98055 [DOI: 10.1371/journal.pone.0098055]
-
Zhu Q, Li J S, Zhang F F, Shen Q, Lin H, Wang L J and Zhu L. 2016. Distinguishing cyanobacteria bloom and aquatic plants in Lake Taihu based on Hyperspectral Imager for the Coastal Ocean images. Remote Sensing Technology and Application, 31 (5): 879–885. [DOI: 10.11873/j.issn.1004-0323.2016.5.0879] ( 朱庆, 李俊生, 张方方, 申茜, 林卉, 王李娟, 朱琳. 2016. 基于海岸带高光谱成像仪影像的太湖蓝藻水华和水草识别. 遥感技术与应用, 31 (5): 879–885. [DOI: 10.11873/j.issn.1004-0323.2016.5.0879] )
-
Zhu Y F, Zhu L, Li J G, Chen Y J, Zhang Y H, Hou H Q, Ju X and Zhang Y Z. 2017. The study of inversion of chlorophyll a in Taihu based on GF-1 WFV image and BP neural network. Acta Scientiae Circumstantiae, 37 (1): 130–137. [DOI: 10.13671/j.hjkxxb.2016.0275] ( 朱云芳, 朱利, 李家国, 陈宜金, 张永红, 侯海倩, 鞠星, 张雅洲. 2017. 基于GF-1WFV影像和BP神经网络的太湖叶绿素a反演. 环境科学学报, 37 (1): 130–137. [DOI: 10.13671/j.hjkxxb.2016.0275] )


