

|
收稿日期: 2017-12-31
基金项目: 国家重点基础研究发展计划(973计划)(编号:2013CB733403);国家重点研发专项课题“区域生物多样性和生态保护红线区成效评估应用示范”;国家自然科学基金(编号:81503183)
第一作者简介: 龙昶宇,1992年生,女,硕士研究生,研究方向为定量遥感在生态学方面的应用。E-mail:lauralcy@126.com
通信作者简介: 万华伟,1981年生,女,研究员,研究方向为生物多样性遥感监测与评价。E-mail:livelyhw@163.com
|
摘要
干旱区生态系统极易受到气候及土地利用变化的影响,其生物多样性格局及其形成机制是重要的生态学问题。基于新疆地区鸟类及哺乳动物物种多样性数据,结合气候、地形和长时间序列的植被遥感参数产品FAPAR数据等,主要在不同的土地利用类型及海拔带上采用单因子相关分析方法探讨了物种丰富度格局的形成机制。总体来说,不同生境类型中,植被遥感参数因子(DHI、NDVI等)与两种类群物种丰富度分布的相关性强于与气候因子(温度、降水)的相关性。具体而言,植被遥感参数因子中,基于FAPAR的生境指数因子与丰富度的相关性大于基于植被指数的因子(DHI_cum>NDVI_cum>EVI_cum);气候因子中,在草地生境或者较低的海拔上,年均降水因子对于丰富度分布的解释力强于年均温度因子。这表明在新疆地区,影响鸟类与哺乳类动物物种丰富度分布的主导理论是生境异质性假说与环境稳定性假说,其解释力在多种生境内均强于生产力与环境热量。
关键词
新疆地区, 物种丰富度格局, 环境因子, 指标参数, 相关关系
Abstract
The distinction of local biodiversity in arid areas is complicated and controversial. Hence, the investigation of the spatial pattern of species richness and its causes in Xinjiang can be used as the basis for biodiversity conservation in the region, and it is also important for regional biodiversity research. On the basis of the data on the distribution of birds and mammals in Xinjiang and long-time series environmental data, such as climate, topography, and remote sensing (FAPAR), this study investigated the species richness and environmental spatial pattern of birds and mammals. Furthermore, the forming mechanism of the disparity pattern of species richness was evaluated through a single-factor correlation analysis used in various land-use types and elevation zones. This research was based on the analysis of the richness of bird and mammal species and the temporal and spatial distribution of environmental factors in Xinjiang. A range of relevant environmental elements were utilized. Then, correlation and linear regression analyses were used to investigate the independent influence of environmental factors on the overall abundance pattern of birds and mammals in a specific living environment in Xinjiang. Finally, the main environmental factors that determine the richness pattern of birds and mammals in specific habitats were selected according to the correlation coefficient. Statistical analysis was realized via the MATLAB software. In general, among the various habitat types, the remote sensing parameter factors (e.g., DHI, NDVI, etc.) are more closely related to the species richness distribution of the two groups than to the climatic factors (e.g., temperature and precipitation). Specifically, among remote sensing parameter factors, the correlation between the habitat index factor based on FAPAR and richness was greater than that of the environmental factor based on vegetation index (DHI_cum>NDVI-cum>EVI-cum). Among the climatic factors, in grassland habitats or at low altitudes, the average annual precipitation factor was better than the average annual temperature factor in verifying the richness distribution. In Xinjiang, the dominant factors affecting the distribution of the richness of bird and mammal species were habitat heterogeneity and environmental stability. Their explanatory power was stronger than the productivity and the environment heat in many types of generation territories. Habitat heterogeneity was the main factor affecting the spatial distribution pattern of richness of birds and mammalian species in Xinjiang, where 70% of the arid areas are not covered by vegetation. Hence, habitat heterogeneity was the main factor affecting the spatial distribution pattern of the richness of bird and mammal species in Xinjiang. Species resources in Xinjiang were abundant, and the overall pattern was complex. The covariation trend of various species and environmental factors was not completely consistent. However, a considerable number of species.
Key words
Xinjiang, patterns of species richness, environmental variables, parameter, correlativity
1 引 言
干旱区或半干旱半湿润区的生态系统多变且不稳定,区内的生物多样性格局的形成机制复杂且充满争议。了解其空间格局及成因,可以为干旱区或半干旱半湿润区的生物多样性保护提供参考和依据。关于物种丰富度空间格局的成因有多种假说(Simpson,1964;Rahbek和Graves,2001),这些假说基于不同的影响因子及其生态、非生态过程,探讨物种丰富度空间格局的形成机制(林鑫 等,2009)。所有假说中研究最深入的分别为能量假说、环境稳定性假说和生境异质性假说(Rosenzweig,1995)。
能量假说认为,太阳辐射转化的各种能量高低与物种丰富度大小成正比(Wright,1983;Hawkins 等,2003b)。由不同能量形式,能量假说进一步划分为环境热量假说(ambient-energy hypothesis)和生产力假说(productivity hypothesis):环境热量假说认为温度是能量的表现形式,温度高的地区,物种丰富度越高(Turner 等,2003;Currie,1991),具体多以环境温度(temperature)或潜在蒸散量PET (potential Evapotranspiration)表示能量的多少;而生产力假说认为食物资源是能量的表现形式,初级生产力较高的地区能维持较高的物种丰富度水平(Hawkins 等,2003b;Clarke和Gaston,2006),通常以基于遥感数据获取的归一化植被指数NDVI (Normalized Difference Vegetation Index)或实际蒸散量AET (Actual Evapotranspiration)表达能量的多少。
环境稳定性假说认为,稳定环境趋使物种生态位狭窄,因此越稳定的地区能容纳更多种类的物种(Klopfer和MacArthur,1960)。生境是生物个体、种群或群落生活地域的环境,包含必需的生存条件和其他对生物起作用的生态要素(林鑫 等,2009)。生境异质性假说认为,区域内的生境越复杂,就能提供更多的小生境和小气候条件,在多变的环境中,物种的生理适应能力更强,从而实现多物种共存,物种丰富度增加(Shmida和Wilson,1985)。
遥感技术的快速发展为物种丰富度形成机制研究提供了更多的数据源,NDVI等植被专题产品已被大量用于物种丰富度的形成机制研究。为更充分利用遥感在描述年内植被生长变化的信息,Mackey等人(2004)首次针对澳大利亚的环境条件提出动态生境指数DHI(Dynamic Habitat Index)的指标,该指标基于FAPAR遥感参数产品计算获取,许多研究表明,冠层顶部光吸收与物种丰富度有较强的线性相关性(Livingston 等,2003),且与初级生产力线性相关(Goward 等,1985)。相比之前用到的单一NDVI参数,DHI有以下几个方面的优势:(1)可以表征森林覆盖,草原牧场,耕地作物等不同类型植被生产季或者全年的生产力状况。(2)也可以表征食物获取与生存最低条件的景观支撑能力,表示全年内的食物和栖息地资源可持续供应的水平,以及其变化可以影响到大多数植食性物种的生存行为、食肉性物种的捕食行为(Schwartz 等,2006)。已有研究表明一年中不同时间提供的植被覆盖对雌性和雄性的种族生存范围有不同的影响(Coops 等,2008)。(3)这类指标还可以表征植被发展的季节性模式,而季节性对诸如食物、水和营养物质等基本资源的影响,将对物种生存的历史轨迹施加选择性压力(McLoughlin 等,2000b)。农耕的土地覆盖类型与季节性草本植被覆盖景观具备较高的变异系数值,而常绿的森林区域的变异系数值则较低。Hobi等人(2017)对比了基于NDVI、EVI、LAI、FAPAR以及GPP计算得到的动态生境指数在物种丰富度分布方面的效用,发现所得到的参量都与物种丰富度分布联系紧密,且反映了鸟类物种丰富度的不同方面。所以,动态生境指数可以全面表征与物种丰富度相关的植被指标,本文在引入NDVI的同时,重点考虑了基于FAPAR的动态生境指数在物种丰富度格局研究中的应用。
新疆地区占地160多万km2,其特殊的山地—盆地生态系统,形成了独特的高山—干旱环境,巨大的环境差异非常适合开展区域尺度的物种丰富度格局研究。根据《新疆鸟类名录》(马鸣,2001)及《新疆哺乳类(兽纲)名录》(阿布力米提·阿布都卡迪尔,2002)等文献资料记载,该区域分布有154种哺乳动物及425种鸟类(李利平 等,2011),本研究基于李利平等人(2011)的物种多样性数据集结合高分辨率的气候、地形、植被遥感参数数据,分析新疆地区鸟类、哺乳类动物物种丰富度以及环境参量的空间格局,并结合几种假说探讨这一典型干旱区内不同生境类型下物种丰富度与环境因子的关系,探讨物种丰富度格局的形成机制。
2 数 据
2.1 物种分布数据
新疆地区鸟类以及哺乳类动物物种分布数据主要基于《新疆鸟类名录》及《新疆哺乳类(兽纲)名录》中记录的物种实际分布范围,在考虑海拔因子基础上,将所有物种的分布数字化,通过ArcGIS10.0做投影转换到WGS-84大地坐标系统。在以往物种丰富度格局研究中,面积是重要影响因素(He和Legendre,1996)。为了消除面积对物种丰富度的影响,本研究最终采用的是处理到10 km×10 km的等面积网格数据。在计算物种丰富度时,首先,将所有物种分布区与网格系统叠加;然后,统计每个网格内出现的物种数目,作为各个网格的物种丰富度值。这一格网系统将新疆地区划分为17454个网格,由于边界地区网格不完整(面积不等),且有的网格区域没有获取到有效的环境数据,可能会影响后续的对比分析,因此研究只选取了面积大于75 km2(即完整网格面积的3/4)以及有有效环境数据的网格(共计12400个)进行分析。
2.2 遥感数据
为了验证各成因假说对于新疆地区物种分布的影响程度以及筛选出可以代表新疆地区环境要素的特征量,本文采用了来自北京师范大学遥感科学国家重点实验室的全球陆表特征参量遥感数据(http://glass-product.bnu.edu.cn/,2016-01,2001-01—2010-12的月数据,GLASS光合有效辐射吸收比1 km产品,时间分辨率为8 d),对使用到的FAPAR遥感参数产品进行了数据预处理与时序特征分析,最终选定了由长时间序列FAPAR数据计算的10年平均夏季累积值(DHI_cum)、10年平均年最小值(DHI_min)、10年平均年际变化率(DHI_sea)以及来自全球植被指数产品(MOD13A3,空间分辨率为1 km,时间分辨率为16 d)计算得到的10年平均的夏季累积平均值(NDVI_cum)和10年平均的夏季累积平均值(EVI_cum)作为遥感参数数据的环境变量参与相关分析。在3.1节以FAPAR数据处理为例介绍本研究确定遥感参数特征变量过程。
2.3 其他数据
除了构建基于遥感数据的参数变量,本研究还利用了温度和降水(MAT、MAP)2种气候因子数据数据来自世界气候数据网站(http://www.worldclim.org/version2/,2015-10,2001-01-01—2010-12-31逐日数据),时间分辨率为24 h,空间分辨率为1 km。
除气候因子外,本研究还使用了地形数据与地表分类数据。地形数据来自美国地质调查局(USGS)1 km数字高程模型(DEM)数据。地表分类数据基于陈军(http://www.globallandcover.com/,2015-10,2010年全球分类数据)的全球30 m分辨率的土地覆盖分类数据,经过拼接剪裁重采样得到新疆地区1 km的土地覆盖分类数据。主要提取有植被分布的土地覆盖类型。
3 方 法
3.1 遥感参数数据特征提取与分析(以FAPAR为例)
3.1.1 FAPAR数据预处理
利用2001年1月—2010年12月的GLASS光合有效辐射吸收比产品(GLASS09A01)数据,对研究区覆盖的6景数据进行图幅拼接及剪裁,按照物种丰富度分布数据,将其重采样到10 km空间分辨率上。
3.1.2 FAPAR时序特征分析
依据2001年—2010年10年平均月最大均值的FAPAR数据集(图1),可知天山山脉与阿尔泰山山脉的FAPAR数值随季节变化比较显著,这可能是区域植被夏季覆盖度较高,植物生长发育、枯落死亡的季节更替造成的。而准噶尔盆地及塔里木盆地则全年几乎无植被覆盖的变化。
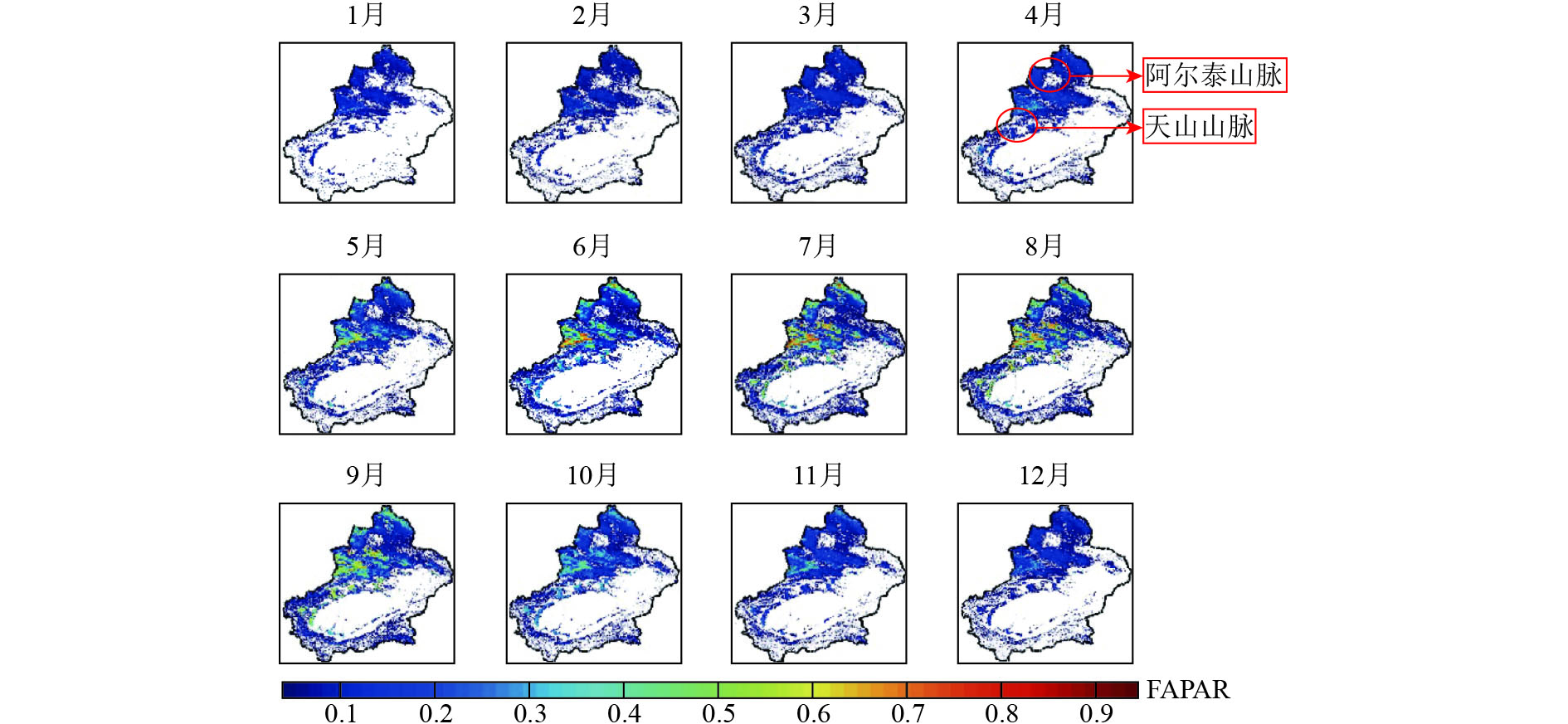

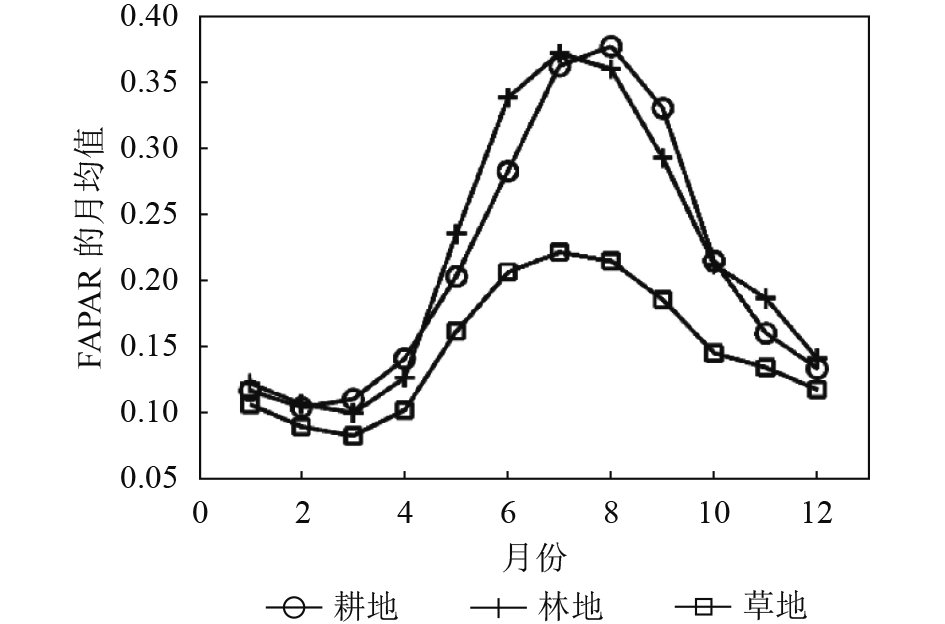
为了得到对丰富度空间分布变化敏感的生境指数,对每年的FAPAR最大值数据集做时序特征分析。在遥感多时相参数数据基础上加上同尺度(10 km)的新疆地区土地利用分类数据,分别提取了新疆地区主要的3种植被类型(耕地、草地、林地)2001年—2010年10年间的月均值。由图2可知新疆地区耕地、林地、草地生境中的FAPAR变化全年呈单峰季节变化趋势,峰值都出现在夏季。其中,林地与草地峰值出现在7月份,耕地类型生境的FAPAR最大值出现在8月份。3种生境类型中,冬季FAPAR数值均接近0.1。
在此基础上,将每年各季相的FAPAR最大值数据集按照春夏秋冬4个季节进行重组取季节平均,再分别与全年平均值做相关对比分析。发现各季相的FAPAR年际变化规律不一致。对比全年平均FAPAR,发现只有夏季平均的FAPAR与年平均的FAPAR趋势基本一致,二者达到极显著相关,其次秋季平均FAPAR与年平均FAPAR年际变化趋势也基本一致,达到显著相关(图3、图4、图5、表1)。从10年整体变化趋势看,除了耕地生境类型的夏秋冬3季与全年均FAPAR在10年间呈增加趋势(图3、图4、图5),林地与草地生境类型的4个季节与年平均FAPAR在2001年—2010年均呈减少趋势。
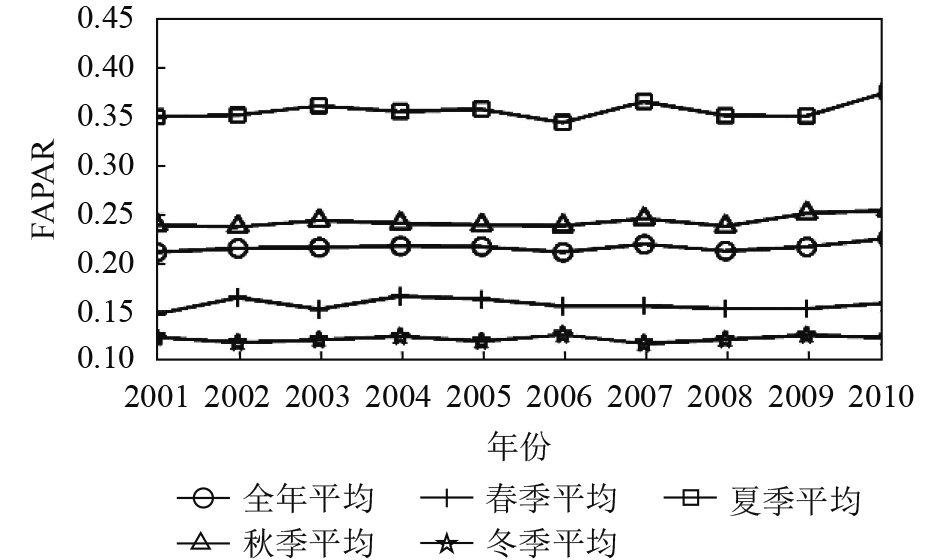
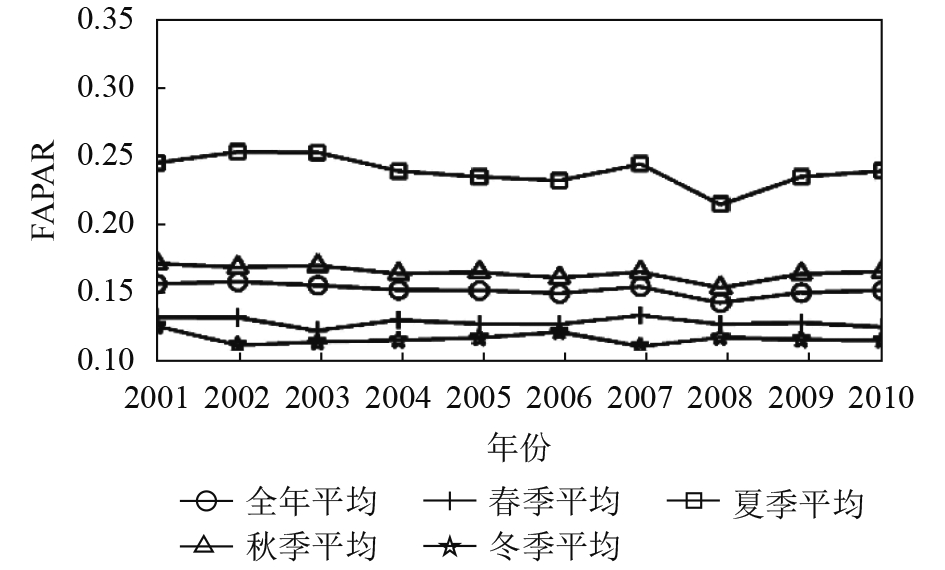
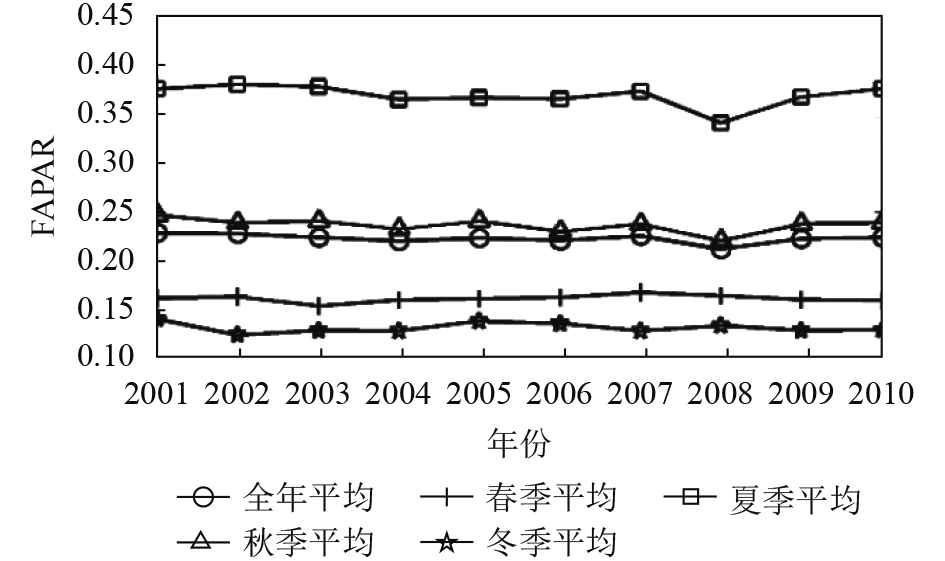
表 1 3种主要生境类型的季节平均年际变化与全年平均年际变化的相关性R2(P<0.05)
Table 1 Correlations R2(P<0.05) between seasonal average change and inter-annual average change of three habitat types
| 季节 | 耕地 | 林地 | 草地 |
| 春季 | 0.1099 | 0.4725 | 0.5613 |
| 夏季 | 0.9630 | 0.9120 | 0.9260 |
| 秋季 | 0.8751 | 0.8290 | 0.9042 |
| 冬季 | 0.0440 | 0.0393 | 0.0107 |
通过时序特征分析以及结合文献资料,得出仅采用夏季平均FAPAR可以很好地代表新疆地区植被的空间分布格局,并以此来构建生境指数。
3.1.3 动态生境指数的计算
本文所用的动态生境指数基于试验区年内的FAPAR数据产品而计算得到,包含3个分量,生产力累计值DHI_cum、生产力最小值DHI_min、生产力年内变异值DHI_sea,各分量的计算公式如式(1)、(2)、(3)所示。
| ${\rm{DHI}}\_{\rm{cum}} = \mathop \sum \limits_{i = 7}^9 {\rm{MAX}}\left( {{{{FAPAR}}_{\left(i \right)}}} \right)$ | (1) |
| $ \begin{split} \!\! & {\rm{DHI}}\_{\rm{min}} = \\ \!\! & MIN\left( {\left({{\rm{MAX}}\left( {{{FAPA}}{{{R}}_{\left(1 \right)}}} \right)} \right), \cdots \left({{\rm{MAX}}\left( {{{FAPA}}{{{R}}_{\left({{i}} \right)}}} \right)} \right)} \right) \end{split} $ | (2) |
| $ \begin{split} & {\rm{DHI}}\_{\rm{sea}} = \\ & \frac{{STD\left( {\left({{\rm{MAX}}\left( {{{FAPA}}{{{R}}_{\left(1 \right)}}} \right)} \right), \cdots \left({{\rm{MAX}}\left( {{{FAPA}}{{{R}}_{\left({{i}} \right)}}} \right)} \right)} \right)}}{{MEAN\left( {\left({{\rm{MAX}}\left( {{{FAPA}}{{{R}}_{\left(1 \right)}}} \right)} \right), \cdots \left({{\rm{MAX}}\left( {{{FAPA}}{{{R}}_{\left({{i}} \right)}}} \right)} \right)} \right)}} \end{split} $ | (3) |
式中,
3.2 各假说指标选取
环境热量假说:采用年均温(MAT)表征一个区域内的环境热量。
生产力假说:采用归一化植被指数年平均的夏季累积平均值(NDVI_cum)、增强型植被指数、年平均的夏季累积平均值(EVI_cum)以及年均降水量(MAP)反应一个地区的生产力水平。在新疆地区,年均降水量与两种植被指数呈强相关性(NDVI:r=0.743,P<0.01;EVI:r=0.875,P<0.01),也可将其作为生产力假说的替代指标。
环境稳定性假说:采用FAPAR年平均年际变化率(DHI_sea)与FAPAR年平均年最小值(DHI_min)作为环境稳定性的衡量指标。
生境异质性假说:采用FAPAR年平均夏季累积值(DHI_cum)作为生境异质性的指示量。
3.3 统计分析
本文主要采用单因素相关分析考察各环境因子在特定的生存环境中对新疆地区鸟类以及哺乳类动物总体丰富度格局的独立影响程度。依照相关系数,选出特定生境中决定鸟类、哺乳类动物丰富度格局的主要环境要素。已有研究表明,陆地尺度的物种分布数据与环境数据普遍存在显著空间自相关,这通常会降低样本独立性,而得到更显著的结果(Diniz-Filho 等,2003)。为了降低区域尺度上物种丰富度与环境影响因子的空间自相关程度,在检验相关系数的显著性水平时,使用了改进的t检验方法(Dutilleul 等,1993),首先估算样本的有效自由度(effective degree of freedom),并以此计算t统计量,再进行显著性检验。统计分析均在MATLAB中实现。
4 结 果
4.1 新疆地区鸟类和哺乳类动物丰富度空间格局
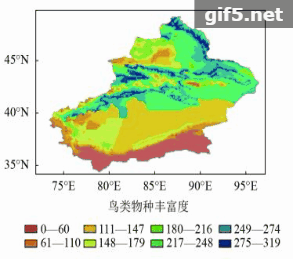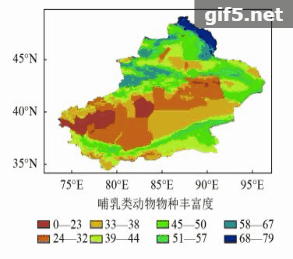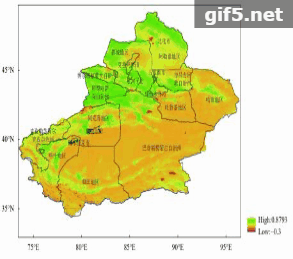
新疆鸟类共有425种,主要分布在阿尔泰山和天山南北坡中低海拔地区(图6)。新疆的哺乳动物共有154种,主要集中分布在山地区,包括阿尔泰山、准噶尔西部山地、天山中西部和昆仑山前山带(图7)。
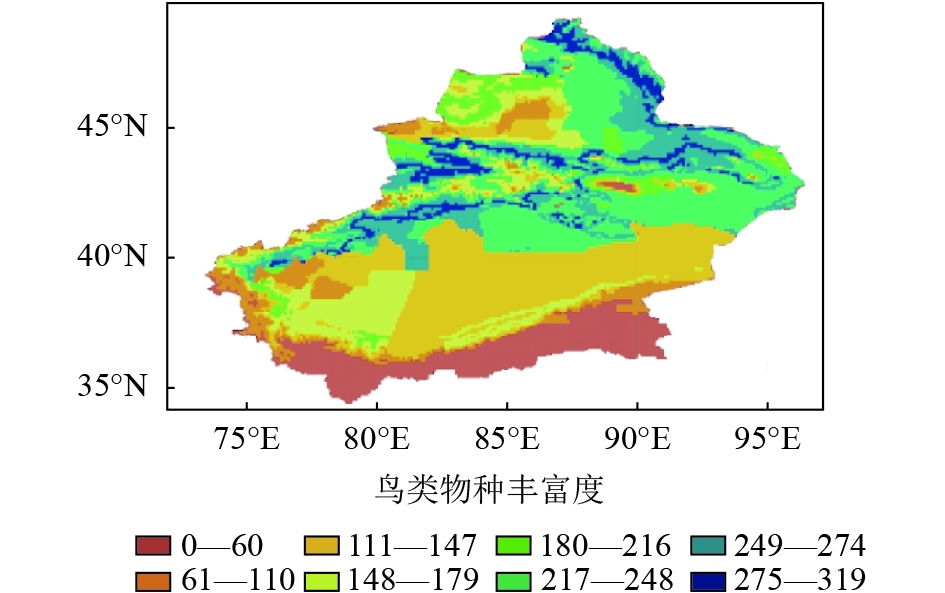

鸟类物种丰富度高值区主要分布在阿尔泰山西部、天山西部和准噶尔西部山地(图6),丰富度最高的850个网格包含了大约19目54科385种鸟类动物,均占鸟类总目、科和种的90%以上,分布范围较为分散,在天山的分布可扩展到天山东部,这可能与鸟类活动范围大有关。哺乳动物丰富度高值区在阿尔泰山和天山西部,整体上相比鸟类来说较为集中,丰富度最高的850个网格包含了7目22科111种哺乳动物,占哺乳动物总物种数的70%(图7)。两个类群物种丰富度高值区并不完全重叠,高值区包括阿尔泰山、准噶尔西部山地、天山中西部地区并延伸到天山东部,共有约1818个网格,约占新疆总面积的10%。
4.2 新疆地区遥感参数空间格局
利用2001年—2010年夏季FAPAR平均值的空间分布来说明新疆植被覆盖空间分布特征(图8)。新疆夏季FAPAR空间分布特征表现为西部与西北部高,东部和东南部低。西部与西北部的山地森林、草地分布区以及天山北坡绿洲的农作物、草地等属于FAPAR高值区;东南部和东部主要为塔克拉玛干沙漠、库木塔格沙漠和戈壁,植被分布稀少,FAPAR值很低,只有沙漠边缘的绿洲有农作物、草地和林地分布,有一定的FAPAR值。其中FAPAR值小于0.1的区域是沙漠、戈壁、水体等无植被地区。由图8可知,新疆的高植被覆盖区主要分布在伊犁哈萨克自治州、昌吉回族自治州的中部、塔城地区与阿勒泰地区的北部。
统计结果表明,植被覆盖区域占新疆总面积的40.57%,无植被区域占59.43%。其中,低值区域(0.1—0.4)占总面积的31.61%,值在0.4—0.5的区域占4.16%,在0.5—0.6的区域占3.14%,大于0.6的区域占1.66%。
4.3 野生物种丰富度与环境参数关系
已有不少研究表明,植被的类型以及其疏密程度可以作为生境的表征(Coops 等,2008)。尤其是目前针对某些特定的区域,还没有较高分辨率的生境图情况下,利用可以表征植被时空分布的遥感参数数据,建立生境指数,是较理想的生境指标,可以研究其对鸟类、哺乳类丰富度的影响。
4.3.1 数据集尺度不一致的影响
有研究表明,植被分布与物种数目分布的相关性会随着采样尺度的减小而降低,但是在监测的任何尺度上都还是会保持一定程度的强相关性(Kerr 等,1997)。本文所使用的遥感参数数据分辨率为1 km,而物种丰富度分布数据分辨率为10 km,二者空间分辨率相差较大,由于在进行后续的单因子分析时,直接用丰富度数据的格网在区域内对环境数据做了平均处理,为了消除统一尺度过程带来的误差,因此首先需要探讨在100 km2尺度范围内的空间异质性对于相关分析的响应情况。
在100 km2的尺度上,计算FAPAR夏季累积指数的方差。发现,随着FAPAR夏季累积值方差逐渐增大,其对于鸟类与哺乳类物种丰富度的分布状况解释能力逐渐降低(表2)。在FAPAR夏季累积值的方差位于0—0.5范围内,其对于鸟类及哺乳类动物物种数目分布的解释力高达90%。总体而言,鸟类与哺乳类动物物种数目随着FAPAR夏季累积值的方差的增加而增加,呈线性递增的状况。即100 km2范围内的FAPAR夏季累积指数的变化越大,鸟类或者哺乳类动物的物种数目就越多。在100 km2的空间范围内,鸟类物种数目在两百种以上、哺乳动物物种数目在60种以上的,其FAPAR夏季累积值的方差均大于1.5。说明,在这一尺度范围内,环境异质性越强,物种种类越丰富。通过对100 km2尺度范围内的FAPAR夏季累积指数方差与鸟类与哺乳类物种丰富度数据做线性相关分析,发现,在任何方差范围内,空间异质性都与丰富度分布呈线性正相关,因此可知,在100 km2尺度上空间异质性对于线性因子分析的影响不大,不会改变影响因素对于结果的正负线性关系。
表 2 FAPAR夏季累积值方差与鸟类、哺乳类物种丰富度线性正相关关系(R2)
Table 2 Positive linear relationship (R2) between summer accumulative Variance of FAPAR and species richness of birds & mammals
| 方差 | 鸟类 | 哺乳类 |
| 0—0.5 | 0.99 | 0.95 |
| 0.5—1 | 0.87 | 0.86 |
| 1—1.5 | 0.78 | 0.79 |
| >1.5 | 0.68 | 0.69 |
4.3.2 单因子相关分析
鸟类、哺乳类动物与各环境因子的单因素分析结果如表3、表4所示。总体来说,新疆地区的鸟类、哺乳类动物物种丰富度的分布与各环境因子的相关性强度是遥感参数因子大于气候因子;对于遥感参数因子来说,各个特定生境中,基于FAPAR的生境指数因子与丰富度的相关性大于基于植被指数的因子(DHI_cum>NDVI_cum>EVI_cum),对于气候因子来说,在草地生境中或者较低的海拔区间内,年均降水因子对于丰富度分布的解释力强于年均温度因子。由此可以得出,对于新疆这样大面积干旱的区域,影响鸟类与哺乳类动物物种丰富度分布的主导理论是生境异质性与环境稳定性,其解释力在多种生境内均强于生产力与环境热量。
表 3 鸟类物种丰富度分布在特定生境中与各因子的线性相关系数(R2)
Table 3 The linear correlation coefficient (R2) of bird species richness in specific habitats and the environment factors
| 气候数据 | 遥感参数数据 | ||||||||
| MAT/℃ | MAP/mm | DHI_cum | DHI_min | DHI_sea | NDVI-cum | EVI-cum | |||
| 地表类型 | 耕地 | 0.19 | 0.01 | 0.52 | 0.57 | 0.22 | 0.43 | 0.41 | |
| 林地 | — | — | — | — | — | — | — | ||
| 草地 | 0.14 | 0.61 | 0.91 | 0.78 | 0.84 | 0.87 | 0.79 | ||
| 海拔区间 | 低海拔 | 0.12 | 0.29 | 0.69 | 0.65 | — | 0.58 | 0.43 | |
| 中海拔 | 0.59 | 0.61 | 0.85 | 0.84 | — | 0.75 | 0.69 | ||
| 高海拔 | 0.78 | 0.05 | 0.74 | 0.75 | — | 0.69 | 0.51 | ||
表 4 哺乳类物种丰富度分布在特定生境中与各因子的线性相关系数(R2)
Table 4 The linear correlation coefficient (R2) of mammal species richness in specific habitats and the environment factors
| 气候数据 | 遥感参数数据 | ||||||||
| MAT/℃ | MAP/mm | DHI_cum | DHI_min | DHI_sea | NDVI-cum | EVI-cum | |||
| 地表类型 | 耕地 | 0.86 | 0.71 | 0.53 | 0.69 | 0.83 | 0.45 | 0.39 | |
| 林地 | — | — | — | — | — | — | — | ||
| 草地 | 0.71 | 0.80 | 0.74 | 0.75 | 0.26 | 0.65 | 0.54 | ||
| 海拔区间 | 低海拔 | 0.86 | 0.73 | 0.76 | 0.76 | — | 0.66 | 0.43 | |
| 中海拔 | 0.58 | 0.42 | 0.75 | 0.75 | — | 0.70 | 0.68 | ||
| 高海拔 | 0.03 | 0.06 | 0.07 | 0.07 | — | 0.05 | 0.02 | ||
具体来说,新疆地区的耕地以及草地生境中的DHI_cum指数与鸟类以及哺乳类动物的物种丰富度的分布呈现较好的线性正相关(R2=0.52,0.91,0.53,0.74,P<0.07);两种生境中的DHI_min指数与鸟类以及哺乳类动物的物种丰富度的分布也呈现较好的线性正相关(R2=0.57,0.78,0.69,0.75,P<0.05);耕地与草地生境中的DHI_sea指数与鸟类以及哺乳类动物的物种丰富度的分布呈线性负相关(R2=0.22,0.84,0.83,0.26,P<0.07)。即新疆地区,分布在耕地或者草地上的鸟类或哺乳类动物物种数目,会随着DHI_cum,min的值的增加而增加,随着DHI_sea值的增加而减少。
在海拔高度1200—4800 m(中海拔),DHI_cum指数可以解释80%的鸟类物种丰富度分布。从低海拔到中海拔,随着海拔升高,DHI_cum指数与鸟类物种丰富度线性越相关,到高海拔地区,相关性逐渐降低。除了在4800—6000 m呈线性负相关,其他海拔带内均呈线性正相关。在新疆地区的中低海拔带,哺乳动物物种丰富度与DHI_cum指数呈线性正相关,而高海拔带区域,其物种丰富度与生境指数呈线性负相关。并且中低海拔带内的鸟类、哺乳动物物种丰富度与DHI_cum指数的相关性远强于高海拔带。说明在海拔较低区域,物种数目的分布更依赖于生境本身的变化和稳定程度。
对于生产力要素而言,NDVI_cum与EVI_cum指数在不同的生境类型和海拔带上均与物种丰富度呈线性正相关。其中,草地生境类型的相关性强于耕地生境类型;中海拔带内的相关性强于低海拔带与高海拔带区域。年均降水量(MAP)与哺乳类动物物种丰富度分布相关性强于与鸟类的物种分布。并且除了在高海拔区域,年均降水量(MAP)与哺乳类动物物种丰富度分布呈线性负相关,其他类型中,年均降水量(MAP)与物种丰富度均呈线性正相关。
对于环境热量要素,年均温(MAT)在不同生境类型与海拔带上与鸟类物种丰富度分布呈线性正相关,与哺乳类动物物种丰富度呈线性负相关。其中,MAT与哺乳类动物的物种丰富度分布的相关性强于鸟类物种。说明在新疆地区,环境热量对哺乳类动物的物种聚集的影响比鸟类物种大。并且区域内热量累积越高,越有利于鸟类物种种类的聚集,而不利于哺乳类物种种类的聚集。
4.3.3 不同丰富度区域的FAPAR变化大小分布
在4.3.2部分得出结论,新疆地区的鸟类与哺乳动物物种丰富与基于FAPAR建立的动态生境指数线性相关最显著,在此基础上对几个参量做空间上的变化验证。
由表5、表6可知,新疆地区的鸟类与哺乳类动物物种60%都分布在FAPAR逐渐减少的区域(2001年—2010年10年间变化率位于–0.1—0)。而30%的物种分布在FAPAR逐渐增加的区域(2001年—2010 年10年间变化率位于0—0.1)。少量物种分布于变化率较大的地区。进一步说明了新疆地区,鸟类与哺乳类动物物种聚集对于生境变化与稳定性的依赖。
表 5 鸟类物种丰富度不同区域的FAPAR变化程度分布
Table 5 The distribution of FAPAR variation of bird species richness in different regions
| 鸟类物种丰富度 | 2001年—2010年FAPAR变化率 | 占比/% |
| 高(≥200) | [0.01,0.02) | 4 |
| [0,0.01) | 38 | |
| [–0.01,0) | 55 | |
| [–0.02,–0.01) | 3 | |
| 中([100,200)) | [0.02,0.03) | 2 |
| [0.01,0.02) | 6 | |
| [0,0.01) | 32 | |
| [–0.01,0) | 58 | |
| [–0.02,–0.01) | 2 | |
| 低(<100) | [0,0.01) | 35 |
| [–0.01,0) | 62 | |
| [–0.02,–0.01) | 3 |
表 6 哺乳类物种丰富度不同区域的FAPAR变化程度分布
Table 6 The distribution of FAPAR variation of mammal species richness in different regions
| 哺乳类物种丰富度 | 2001年—2010年FAPAR变化率 | 占比/% |
| 高(≥50) | [0.01,0.02) | 2 |
| [0,0.01) | 25 | |
| [–0.01,0) | 67 | |
| [–0.02,–0.01) | 6 | |
| 中([25,50)) | [0.02,0.03) | 1 |
| [0.01,0.02) | 5 | |
| [0,0.01) | 46 | |
| [–0.01,0) | 47 | |
| [–0.02,–0.01) | 1 | |
| 低(<25) | [0.01,0.02) | 1 |
| [0,0.01) | 34 | |
| [–0.01,0) | 62 | |
| [–0.02,–0.01) | 3 |
5 结 论
5.1 主要结论
(1)新疆地区的鸟类、哺乳类动物物种丰富度的分布与各环境因子相关性表明,植被遥感参数与物种丰富度显示强相关,高于气候因子。在不同的植被参数之间,基于FAPAR的动态生境指数的DHI_cum指标解释力强于NDVI、EVI等植被指数。因此,植被遥感参数,尤其是动态生境指数,在新疆地区鸟类和哺乳类丰富度格局解释中具有重要的作用。
(2)在文中所用的10 km网格尺度上,环境异质性越强,物种种类越丰富。且90%以上的物种生活在生境变化较小的区域。因此,在新疆地区,影响鸟类与哺乳类动物物种丰富度分布的主导理论是生境异质性与环境稳定性,其解释力在多种生境内强于生产力与环境热量。
(3)不同海拔区间的研究结果表明,在海拔较低区域,物种数目的分布更依赖于生境本身的变化和稳定程度。而低海拔区是最易受到人类活动影响的区域,因此更要维持该区域的生境稳定性。
(4)气候因子是影响新疆地区物种丰富度格局的重要因素,对于气候因子来说,在草地生境中或者较低的海拔区间内,年均降水因子对于丰富度分布的解释力强于年均温度因子。环境热量对哺乳类动物的物种聚集的影响比鸟类物种大,区域内热量累积越高,越有利于鸟类物种种类的聚集,而不利于哺乳类物种种类的聚集。
5.2 讨论
5.2.1 新疆地区鸟类与哺乳类动物物种丰富度格局的主要影响因素
本研究结果显示,新疆地区的鸟类与哺乳类动物的物种丰富度与各种环境因子有不同程度的相关性,而代表生境异质性与环境稳定性的环境因子对于物种丰富度格局的分布解释率最高,达76%。其中,表征生境异质性的环境因子(DHI_cum指数)与物种丰富度的相关性最强,表明对于新疆的70%都无植被覆盖的干旱区域,生境异质性是影响新疆地区鸟类与哺乳类动物物种丰富度空间分布格局的主要因素。该结果不同于以往对于陆地尺度上物种丰富度与不同环境因子的关系程度以及生产力资源较丰富的地区物种丰富度与不同环境因子的关系程度,主要支持了生境异质性假说。
5.2.2 新疆地区鸟类与哺乳类动物物种丰富度格局及其成因的复杂性
本研究涉及鸟类与哺乳类动物共计28目579种,新疆地区物种资源丰富,同时整体格局较为复杂,不同物种与环境因子的共变趋势并不完全一致,例如,哺乳类动物物种丰富度与环境热量因子均呈负相关,这与哺乳动物的生理机制以及对极端条件的适应有关。鸟类物种丰富度与DHI_cum指数关系最为密切,这与鸟类物种多分布于有植被覆盖的地表有关系,其物种丰富度主要受到植被类型以及疏密程度的影响。
此外,虽然研究中分析的因子已经解释了80%的物种丰富度空间分布,但仍有相当一部分物种的地理分异成因未被表达出来。说明仍有其他可能影响区域尺度的物种丰富度分布的因素未被考虑,比如局地尺度的环境因子(物种之间的干扰、捕食竞争关系等),但这些因子目前尚难以空间化,随着技术的发展,可进一步研究局地尺度环境因子的影响。
参考文献(References)
-
Abramite AbuQadir. 2002. A list of mammals (Oryzae) in Xinjiang. Beijing: science Press (阿布力米提·阿布都卡迪尔. 2002. 新疆哺乳类(兽纲)名录. 北京: 科学出版社)
-
Clarke A, Gaston KJ. 2006. Climate, energy and diversity. Proceedings of the Royal Society B: Biological Sciences, 273 : 2257–2266.
-
Coops N C, Wulder M A, Duro D C, Han T and Berry S. 2008. The development of a Canadian dynamic habitat index using multi-temporal satellite estimates of canopy light absorbance. Ecological Indicators, 8 (5): 754–766. [DOI: 10.1016/j.ecolind.2008.01.007]
-
Currie DJ. 1991. Energy and large-scale patterns of animal and plant-species richness. The American Naturalist, 137 : 27–49. [DOI: 10.1086/285144]
-
Diniz-Filho JAF, Bini LM, Hawkins BA. 2003. Spatial autocorrelation and red herrings in geographical ecology. Global Ecology and Biogeography, 12 : 53–64. [DOI: 10.1046/j.1466-822X.2003.00322.x]
-
Goward, S.N., Tucker, C.J., Dye, D.G. 1985. North American vegetation patterns observed with the NOAA-7 advanced very high resolution radiometer. Vegetation, 64 : 3–14. [DOI: 10.1007/BF00033449]
-
Hawkins B A, Field R, Cornell H V, Currie D J, Guégan J F, Kaufman D M, Kerr J T, Mittelbach G G, Oberdorff T, O’Brien E M, Porter E E and Turner J R G. 2003b. Energy, water, and broad-scale geographic patterns of species richness. Ecology, 84 (12): 3105–3117. [DOI: 10.1890/03-8006]
-
He F L and Legendre P. 1996. On species–area relations. The American Naturalist, 148 (4): 719–737. [DOI: 10.1086/285950]
-
Hobi M L, Dubinin M, Graham C H, Coops N C, Clayton M K, Pidgeon A M and Radeloff V C. 2017. A comparison of Dynamic Habitat Indices derived from different MODIS products as predictors of avian species richness. Remote Sensing of Environment, 195 : 142–152. [DOI: 10.1016/j.rse.2017.04.018]
-
Kerr J T and Packer L. 1997. Habitat heterogeneity as a determinant of mammal species richness in high-energy regions. Nature, 385 (6613): 252–254. [DOI: 10.1038/385252a0]
-
Klopfer P H and MacArthur R. 1960. Niche size and faunal diversity. The American Naturalist, 94 (877): 293–300. [DOI: 10.1086/282130]
-
Li L P, Yin L K and Tang Z Y. 2011. Distribution patterns of the species richness of plants and animals in Xinjiang, China. Arid Zone Research, 28 (1): 1–9. [DOI: 10.3724/SP.J.1148.2011.00001] ( 李利平, 尹林克, 唐志尧. 2011. 新疆野生动植物物种丰富度的分布格局. 干旱区研究, 28 (1): 1–9. [DOI: 10.3724/SP.J.1148.2011.00001] )
-
Lin X, Wang Z H, Tang Z Y, Zhao S Q and Fang J Y. 2009. Geographic patterns and environmental correlates of terrestrial mammal species richness in China. Biodiversity Science, 17 (6): 652–663. [DOI: 10.3724/SP.J.1003.2009.09065] ( 林鑫, 王志恒, 唐志尧, 赵淑清, 方精云. 2009. 中国陆栖哺乳动物物种丰富度的地理格局及其与环境因子的关系. 生物多样性, 17 (6): 652–663. [DOI: 10.3724/SP.J.1003.2009.09065] )
-
Livingston M, Shaw W W and Harris L K. 2003. A model for assessing wildlife habitats in urban landscapes of eastern Pima county, Arizona (USA). Landscape and Urban Planning, 64 (3): 131–144. [DOI: 10.1016/S0169-2046(02)00217-7]
-
Ma Ming. 2001. List of Birds in Xinjiang. Beijing: science Press (马鸣. 2001. 新疆鸟类名录. 北京: 科学出版社)
-
Mackey B, Bryan J and Randall L. 2004. Australia’s dynamic habitat template 2003 // Proceedings of the MODIS Vegetation Workshop II. Missoula, MT, American: [s.l.]
-
Rosenzweig M L. 1995. Species Diversity in Space and Time. Cambridge: Cambridge University Press
-
Schwartz, C. C., Haroldson, M. A., White, G. C., Harris, R. B., Cherry, S ., Keating, K. A., Moody, D ., Servheen, C .. 2006. Temporal spatial and environmental influences on the demographics of grizzly bears in the Greater Yellowstone Ecosystem. Wildlife Monogr, 161 : 1–68. [DOI: 10.2193/0084-0173(2006)161[1:TSAEIO]2.0.CO;2]
-
Shmida A and Wilson M V. 1985. Biological determinants of species diversity. Journal of Biogeography, 12 (1): 1–20. [DOI: 10.2307/2845026]
-
Simpson G G. 1964. Species density of North American recent mammals. Systematic Zoology, 13 (2): 57–73. [DOI: 10.2307/2411825]
-
Turner W, Spector S, Gardiner N, Fladeland M, Sterling E and Steininger M. 2003. Remote sensing for biodiversity science and conservation. Trends in Ecology and Evolution, 18 (6): 306–314. [DOI: 10.1016/S0169-5347(03)00070-3]
-
Wright D H. 1983. Species–energy theory: an extension of species–area theory. Oikos, 41 (3): 496–506. [DOI: 10.2307/3544109]


