 2022,
Vol. 31
2022,
Vol. 31



海参属底栖食性生物[1],摄入的食物复杂,其中包括一些病原微生物。微生物作为衡量养殖环境优劣的指标之一,其数量变化与海参细菌性疾病的发生密切相关。在养殖池塘中,异养菌是池塘环境中的主要分解者,能降解大量有机物,并且可有效地提高池塘自净能力。但部分异养菌本身也是条件致病菌, 在环境条件恶化等情况下会导致海参死亡[2]。弧菌是一种条件性致病菌,其数量会影响参池环境。马悦欣等[3]报道弧菌属的细菌是大连地区海参大面积流行病的病原之一。黄华伟[4]在研究海参腐皮综合征的发生与异养菌区系间的关系时发现, 低温时期发病池塘表层底泥异养菌、弧菌总数平均为3.7×106、1.3×105cfu/g,渡夏时期发病池塘表层底泥异养菌、弧菌总数平均为5.9×106、1.4×106cfu/g,并且发病参池底泥弧菌与异养菌数比值较正常参池高。陈文博等[5]研究表明,在高温养殖期,易引发海参病害的老池塘弧菌总数(3.365×105~3.525×105cfu/g)显著高于新池塘(2.545×105~2.735×105cfu/g)。
自然纳潮换水是一种传统水质调控方式,即池塘水体全部引用外海海水,尽管该方式操作简单,但常受自然天气及外海海水水质不稳定等制约,并不能真正提高池塘水质。微孔曝气是海参池塘常见的水质调控方式,该技术可有效地对池塘底部增氧,抑制有害微生物生长。林海等[6]对不同増氧方式及沉水植物修复河蟹池塘养殖环境的研究时指出,在养殖高温季节曝气增氧可以有效改善养殖水体水质。何义进等[7]研究发现,夏季高温时期,使用微孔增氧技术的河蟹养殖池塘弧菌数[(3.28±0.11) cfu/g]明显低于不充氧池塘[(5.33±0.27) cfu/g],对由弧菌引起的细菌性疾病有良好的预防效果。翟海佳[8]发现微孔曝气池塘中真菌相对细菌的含量长期处于较高水平。新型养水机是笔者团队自主研制养殖池塘水质调控设备,其工作原理是将表层优良水体经过生物加工注入池塘底部,利用机械设备搅动局部水体,在水循环动力作用下,有效地改善养殖池塘水质。相关研究[9]表明,参池使用养水机后,海参成活率及单产均有所提高。目前养殖池塘水质调控方式层出不穷,但在微生物方面对各种新型调控方式的比较研究相对较少,寻求新型的合理有效的养殖调控方式是当前急需解决的问题。
本研究在自然纳潮、微孔曝气和新型养水机3种水质调控下,对参池底泥中异养菌和弧菌的数量变化进行周年检测,探究不同水质调控方式对参池底质微生物数量的影响,旨在3种水质调控方式下为参池底质及生物的变化提供理论依据[10]。
1 材料与方法 1.1 试验参池本研究在大连庄河市宝发海珍品有限公司进行,试验选取3种参池,分别为2号(39.787 0°N,123.323 0°E)、4号(39.786 6°N,123.320 8°E)、11号(39.787 3°N,123.325 5°E)参池。参池为长605 m、宽85 m的标准矩形,南北走向。参池底质均为泥沙质,并均匀铺设网礁。参池采用水泥护坡,南北各有一进、排水闸门,参池南深北浅,深度相差0.5 m。每月(初一、十五)大潮期间换水3~5 d,年水深变化为1.2~2.0 m。各参池内海参规格、密度相同,所有参池不投饵、不投药,试验期间统一管理。
2号为养水机池塘,在参池最深处(南端)放置1台功率为750 W的养水机,每日21:00至次日9:00工作12 h,冬季正常工作。养水机(CN200620006141.5)主要由4部分组成,分别为进水组件(CN201621364576.7)、生物包、水动力装置和养殖池塘专用喷头(CN201410415402.8),见图 1,设备介绍及工作机理参考文献[9, 11-13];4号为自然池塘无其他养殖装置。11号为微孔曝气池塘在参池底部铺设底充氧装置,装置由微孔曝气盘、总供气管和空压机(1.5 kW/hm2)组成,当参池缺氧时开始工作,微孔曝气设备介绍及工作机理参考文献[14-16], 各参池基本情况和周年水化学特征参考文献[17-19],如表 1所示。

|
图 1 养水机模拟图 Fig. 1 Simulation diagram of water quality regulator |

|
表 1 养殖池塘基本状况和周年水化学特征 Tab.1 Basic situation and anniversary physical chemistry features in the ponds |
2015年10月—2016年9月,每逢满月大潮前3~5 d进行采样。选择各参池最深点(南端)为试验点,设置3个采样重复。参照《国家海洋调查规范》海洋生物调查中规定的方法进行采集。使用圆柱形采泥器采集泥样,采集样本为截面直径5 cm,取0~5 cm泥样,装入无菌自封袋中带回,在4 ℃下保存待测。
1.3 微生物数量的检测 1.3.1 异养菌总数对泥样进行梯度稀释,取适当的3个浓度,采用平板涂布法吸取0.1 mL样品涂布在2216E培养基上,每组3个重复,25 ℃下培养7 d后,选取菌落平均数为30~300的平板进行计数。
2216E培养基:选择市售粉末状成品培养基(青岛高科园海博生物技术有限公司),用蒸馏水加热溶解后在121 ℃下灭菌15 min。
1.3.2 弧菌总数对泥样进行梯度稀释,取适当的3个浓度,采用平板涂布法吸取0.1 mL样品涂布在TCBS培养基上,每组3个重复,25 ℃下培养3 d后,选取菌落平均数为30~300的平板进行计数。
TCBS培养基:选择市售粉末状成品培养基(青岛高科园海博生物技术有限公司),用蒸馏水加热溶解。
1.4 微生物菌落计算过程将所得样品混匀,一部分用于测量含水量,一部分泥样经过稀释、接种、倒平板无菌操作后,置于25 ℃恒温培养箱中进行培养,待菌落长出后进行计数:
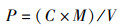 (1)
(1)
式中:P为每克干泥菌含量,cfu/g; C为菌落平均数,cfu;M为稀释倍数;V为泥样经105 ℃烘烤8 h后其含水量测定值,g。
1.5 数据处理运用Excel 2007及SPSS 17.0软件对试验数据进行分析处理。
2 结果 2.1 3种水质调控方式下参池底泥中异养菌数量的周年变化3种水质调控方式下各参池底泥中异养菌数量变化周年呈不规则波动,波动范围自潮池塘为94 400~377 625 cfu/g、微孔曝气池塘为62 633~247 309 cfu/g、养水机池塘为11 5037~273 071 cfu/g,周年极差比较养水机池塘最小(R养:158 034>R微:184 676>R自:283 225),见图 2。自然池塘最高值出现在1月,最低值出现在3月。微孔曝气池塘最高值出现在5月,最低值出现在6月。养水机池塘最高值出现在4月,最低值出现在8月。

|
图 2 3种水质调控方式下参池底泥中异养菌数量的周年变化 Fig. 2 Annual change of the number of heterotrophic bacteria in bottom mud of sea cucumber farming pond under three water quality control methods |
分别按照四季变化测定3组参池底泥中异养菌数量,如图 3所示。结果显示,自然池塘、微孔曝气池塘、养水机池塘异养菌数量四季变化范围分别为141 153~295 997、87 159~169 272、139 627~192 938 cfu/g。统计分析表明,在相同季节、不同组别的参池异养菌数量具有不同差异水平,春(3—5月)、秋(9—11月)两季各参池异养菌数量差异均不显著(P>0.05),夏季(6—8月)微孔曝气池塘异养菌数量显著低于自然池塘和养水机池塘(P<0.05)且另2种参池之间差异不显著(P>0.05),冬季(12—2月)自然池塘异养菌数量显著高于微孔曝气池塘和养水机池塘(P<0.05)且另2种参池之间差异不显著(P>0.05)。

|
不同的小写字母表示不同季节间数据有显著的统计学差异(P>0.05)。 Different lowercase letters indicate significant differences between the seasons. 图 3 3种水质调控方式下参池底泥中异养菌数量的季节变化 Fig. 3 Seasonal changes of the number of heterotrophic bacteria in bottom mud of sea cucumber farming pond under three water quality control methods |
3种水质调控方式下各参池底泥中弧菌数量变化周年变化趋势相近,自然池塘为0~3 291 cfu/g、微孔曝气池塘为35~5 412 cfu/g、养水机池塘为0~3 037 cfu/g,周年极差比较养水机池塘最小R养:3 037<R自:3 291<R微:5 377(图 4)。微孔曝气池塘最高值出现在4月,自然池塘和养水机池塘最高值均出现在5月,3参池最低值出现在2月。

|
图 4 3种水质调控方式下参池底泥中弧菌数量的周年变化 Fig. 4 Annual change of vibrio number in bottom mud of sea cucumber farming pond under three water quality control methods |
分别按照四季变化测定3组参池底泥中弧菌数量,如图 5所示。结果显示,自然池塘、微孔曝气池塘、养水机池塘弧菌数量四季变化范围分别为325~1 470、301~2 972、853~1 253 cfu/g。参池底泥弧菌呈“春季高,冬季低”的变化特征。统计分析表明,在相同季节、不同组别的参池异养菌数量具有不同差异水平,夏(6—8月)、秋(9—11月)、冬(12—2月)三季各参池弧菌数量差异均不显著(P>0.05),春季(3—5月)微孔曝气池塘异养菌数量显著高于自然池塘和养水机池塘(P<0.05)且另2种参池之间差异不显著(P>0.05)。

|
不同的小写字母表示不同季节间数据有显著的统计学差异(P>0.05)。 Different Lowercase letters indicate significant differences between the seasons. 图 5 3种水质调控方式下参池底泥中弧菌数量的季节变化 Fig. 5 Seasonal changes of vibrio number in bottom mud of sea cucumber farming pond under three water quality control methods |
由图 6可以发现:3种方式各参池底泥中弧菌与异养菌总数比值在4、5月相对较高,在2月较低;自然池塘周年变化范围为0~0.034;微孔曝气池塘周年变化范围为0~0.031;养水机池塘周年变化范围为0~0.017;参池底泥中弧菌与异养菌数比值呈“春季高,冬季低”的变化特征;在春季(3—5月)各参池3个月平均比值由低到高分别为养水机池塘(0.007)<自然池塘(0.014)<微孔曝气池塘(0.016),3参池差异不显著(P>0.05);在冬季(12—2月)各参池3个月平均比值由低到高分别为自然池塘(0.001)<微孔曝气池塘(0.003)<养水机池塘(0.004),3参池差异不显著(P>0.05)。

|
图 6 3种水质调控方式下参池底泥中弧菌与异养菌数比值的周年变化 Fig. 6 Annual change of the ratio of vibrio to heterotrophic bacteria in bottom mud of sea cucumber farming pond under three water quality control methods |
异养菌作为养殖环境中细菌的主要组成部分[20]具有提高池塘自净能力的作用,也是大多数水生生物营养成分的来源。相关研究指出底泥中异养菌数量较多,对海参的粪便等杂质利用率较高。众多学者[4, 21-22]认为异养菌的大量繁殖可以加快养殖环境中有机物的分解速度,在海参养殖期间细菌能够及时快速分解掉养殖生物未利用的残饵和产生的粪便等有机物,有效避免了有机物大量积累而导致的环境的恶化。本实验中自然、微孔曝气、养水机各池塘底泥异养菌数量周年平均值分别为186 569.60、120 225.27、173 152.19 cfu/g。养水机池塘底泥异养菌数量虽非最高,但有机质含量周年平均值(0.83 %)和底泥弧菌数量周年平均值(998 cfu/g)均为最低,说明养水机管理方式对有机质利用更充分,避免有机质过度积累,并防止致病菌造成参池底质恶化。相关研究[23-24]报道,养殖池中的各种细菌应维持在相对稳定的动态平衡状态。本实验中自然池塘底泥异养菌数量周年波动幅度(6.96×109)高于养水机池塘(2.90×109)和微孔曝气池塘(2.51×109)。分析原因可能是自然池塘底部没有增氧,而微孔曝气与养水机有助于提高养殖系统底部溶氧浓度,带动细菌的生长繁殖,并且养水机运行时水体会出现长时间的对流作用,不仅改善了水体环境,并逐渐深入调控了底泥的环境,改善底泥长期厌氧环境。
季节变化的比较上,养水机池塘季节变化幅度(5.44×108)明显低于另2种参池。首先异养菌与浮游植物生长繁殖有着密切关系[25],异养菌可以吸收浮游植物所产生的有机物质从而促进自身生长繁育,同时为浮游植物生长提供必要的营养成分,并为其微生长环境起到调节作用。3种参池周年平均生物量大小为养水机池塘(20.12 mg/L)、微孔曝气池塘(17.44 mg/L)、自然池塘(16.66 mg/L),养水机池塘浮游植物生物量明显高于另2种参池,且各季节生物量变化均较自然池塘和微孔曝气池塘更稳定[26]。同时pH的合宜不仅可以促进参池中浮游植物光合作用,还可以促进异养菌分解作用,增加参池水体溶氧和有机质浓度。3种参池周年pH变化范围均处于海参生长安全的范围内,而养水机池塘pH的周年变化更稳定,减小各个水层之间的pH差值[17]。其次养殖池塘异养细菌数量受营养盐积累影响,由于异养细菌是典型的基质限制性生物,其个体微小、繁殖速度快、生命周期短,营养成分均衡有利于促进异养菌的大量繁殖,降低有害菌数量[27]。3种池塘DIN和活性磷酸盐季节变化幅度大小比较为养水机池塘<底充氧池塘<自然池塘,且养水机池塘DIN周年均值小于另2种参池,而活性磷酸盐周年均值大于另2种参池。夏斌等[28]、乔方利等[29]研究发现,高氮低磷的水体环境会造成底质环境恶劣,严重影响海参健康。同时参池底质中微生物活性与底泥耗氧率大小有关,在化冰时期以及夏季高温季节,参池底部的水温较高,底泥耗氧率将会增大,底质中微生物的活性也会升高。本课题另有证明[18],各参池春季(3、4月)、夏季(7、8月)的底泥耗氧率明显高于其他月份。夏季初始降雨频繁,养水机池塘底泥异养菌数量高于2种参池塘,此时3种参池水温最大值提早出现在7月份,而溶解氧达到最小值。牛宇峰[30]研究发现参池底部缺氧不利于异养菌生长,危害海参健康。鲁晓倩[17]报道本试验夏季高温期养水机池塘溶解氧要高于其他2种参池,说明养水机给参池底部提供了充分氧气,促进了异养菌的生长,改善底泥长期厌氧环境。
3.2 3种水质调控方式对参池底泥中弧菌数量的影响在养殖环境条件恶化等情况下,弧菌是近海河口环境中常见的兼性厌氧、化能异养细菌,是引起海参细菌性疾病发生的主要原因之一,其数量分布与水产动物发病概率有着密切的关系[31]。弧菌病发病时间短、死亡快、传染性极强,严重制约着水产经济发展[32]。本研究发现参池底泥弧菌数量与异养细菌数量变化不同步,且其数量在一年中处于不断波动状态,这与其他报道相符。在高密度养殖环境中,若条件致病的弧菌总数处于高位水平,则对于海参的正常生长将构成威胁。3种调控方式下,养水机池塘底泥弧菌数量周年平均值(998 cfu/g)低于自然池塘(1 024 cfu/g)和微孔曝气池塘(1 242 cfu/g)。同时本试验发现养水机能够很好地维持参池弧菌数量的稳定性,底泥弧菌数量周年波动幅度(1.02×106 cfu/g)低于自然池塘(1.29×106 cfu/g)和微孔曝气池塘(2.57×106 cfu/g)。这是由于养水机配备独有的生物包,工作时生物包内接种的异养菌通过养水机射流方式随水流释放到池塘底部环境,参与底泥有机质降解,在弧菌大量繁殖的季节具有调节作用,故养水机对底泥弧菌有明显的抑制效果。
季节变化的比较上,各参池底泥弧菌变化规律一致,均呈“春季高,冬季低”的变化特征,与陈文博等[5]研究结论相符。春季(3—5月)养水机池塘底泥弧菌数量低于另2种参池的分析:春季是海参生长的适宜季节, 此时参体排泄物增多,有利于基质限制性细菌的生长繁殖,3种参池出现弧菌数量季节高峰。相关研究[33-35]指出春季随着气温回升的加快,受光照和气温的影响,参池表层水体水温较高,而底层水温较低,整个池水容易形成水温的分层,各种生物的代谢耗氧率迅速上升,底泥中厌氧或兼性厌氧细菌开始繁殖。海参属底栖杂食性生物,底质环境直接影响海参生长、摄食等活动。春季参池出现水温分层时在养水机作用下可有效调控上、下水体,抑制弧菌的繁殖,改善底质状况。同时笔者在现场观测到养水机池塘底泥不出现臭底、烂底现象也证明了上述观点。冬季(12—2月)养水机池塘底泥弧菌数量高于另2种参池的分析:冬季各参池水温均达到全年最低值,低温条件抑制了细菌的生长和有机物质的分解,3种参池均达到最低值,并且差异不显著。郑晓叶等[27]研究认为底泥中弧菌数量则受营养盐累积的影响较大。海参活动和底泥有机质分解会消耗底层水溶解氧,在底层水溶解氧浓度过低时,厌氧细菌会大量繁殖造成有机物厌氧分解,产生对参池有害物质,如氨氮、亚硝酸盐等。冬季养水机池塘底泥弧菌数量虽高于2种参池,但养水机池塘无机氮浓度总体维持在较低水平,并且表层和底层之间的浓度差较另2种参池更小。由于养水机内的生物包发挥了积极作用,增加了水中的益生菌浓度,所以弧菌数量虽高于2种参池但未对参池环境造成影响。此外冬季参池中浮游藻类繁殖会产生大量的氧气,海参、浮游动物和微生物等耗氧率降低,3种参池溶氧的含量均出现峰值,养水机池塘冬季溶解氧(12.0 mg/L)高于另2种参池,养水机对提高水体溶解氧浓度和促进海参健康生长起到了积极作用。高溶氧环境保障了有机质氧化反应的进行。据党子乔[18]对池塘沉积物研究,冬季3种参池底泥有机质含量比较自然池塘>微孔曝气池塘>养水机池塘证实了上述观点。
3.3 3种水质调控方式对参池底泥中弧菌与异养菌数比值的影响弧菌病的发生主要取决于致病菌数和异养菌总数的比值。相关研究[36]指出,参池底泥弧菌与异养菌数比值在春季较大。本试验3种参池底泥弧菌与异养菌数比值呈“春季高,冬季低”的周年变化特征,与现有研究结果一致。迟爽等[22]和李彬等[24]研究参池发现底泥弧菌与异养菌数比值越低,海参患病概率越低。从各参池的变化数据中可发现,养水机池塘弧菌与异养菌数比值明显低于另2种参池,对海参病害的预防起到了积极的作用。本实验中,春季(3—5月)各参池底泥弧菌与异养菌数比值逐渐升高,这与参池在春季尤其是“惊蛰”前后易暴发病害问题相符。此时养水机池塘底泥中弧菌与异养菌数之比最低,自然池塘次之,微孔曝气最高。在养殖高温时期,自然池塘最高,微孔曝气与养水机池塘比值相近,但微孔曝气稍高。从7月开始参池弧菌与异养菌数比值逐渐回升,分析原因可能是夏季7月参池水温升高,水体初级生产力会随之升高,浮游植物数量增加,有机物的分解速率加快,为细菌的生长繁殖创造了有利环境条件。已有相关研究[37]得出,随着水温升高,养殖环境中的细菌数量和种类也会随之增加。因此,在春季和夏季应密切关注参池底泥异养菌、弧菌数量的变化,及时发现问题并采取措施,避免海参生病死亡情况的发生。冬季结冰期(12—2月)由于水体中的有机物质沉降至底泥中,海参营底栖生活,冬季结冰后海参摄食少,无法充分利用沉积物。随着参池底部底泥层的富集和增厚,容易造成病原菌的不断滋生。同时,当养殖池塘水体富营养化比较严重,水体营养过剩会抑制某些对环境敏感的细菌生长[38],进而导致池塘底部溶解氧消耗和厌氧微生物大量滋生,产生有害代谢产物,造成养殖生物的死亡。前期研究发现[17],3种参池水体营养级别属于磷中等限制潜在性富营养水质,而养水机对参池水体中的N∶ P即水体营养结构的控制作用更优于另2种调控方式。综合分析比较各参池间弧菌与异养菌数比值的差异可以发现,养水机设备优于微孔曝气和自然纳潮调控方式,可使参池弧菌与异养菌比值在周年内维持在较低水平,在养殖发病率较高的季节有效抑制了底质有害细菌的滋生,在一定程度上降低了海参发病的可能。
3.4 总结综上所述,养水机对参池底泥异养菌和弧菌的调控效果优于另2种调控方式,同时期养水机池塘溶解氧、底泥有机质、营养盐含量、浮游植物生物量和初级生产力等指标方面,表现出可促进池塘上、下层水体交换,帮助水体与底泥之间物质循环,有效解决底层水体缺氧问题,避免弧菌的厌氧发酵,改善池塘底质的效果,与实践上淤泥少、无杂草、少病害的现象一致。因此,笔者认为海参养殖池塘搭配水质辅助调控设备可降低致病菌数量和海参患病的威胁。
| [1] |
田传远, 李琪, 梁英. 刺参健康养殖技术[M]. 青岛: 中国海洋大学出版社, 2008: 20-21. TIAN C Y, LI Q, LIANG Y. Sea cucumber healthy breeding technology[M]. Qingdao: Ocean University of China Press, 2008: 20-21. |
| [2] |
杨莺莺, 李卓佳, 梁晓华, 等. 芽胞杆菌对鱼池微生物群落代谢功能的影响[J]. 微生物学杂志, 2009, 29(3): 11-17. YANG Y Y, LI Z J, LIANG X H, et al. Influences of bacillus on metabolic functions of microbial communities in fishponds[J]. Journal of Microbiology, 2009, 29(3): 11-17. DOI:10.3969/j.issn.1005-7021.2009.03.003 |
| [3] |
马悦欣, 徐高蓉, 张恩鹏, 等. 仿刺参幼参急性口围肿胀症的细菌性病原[J]. 水产学报, 2006, 30(3): 377-382. MA Y X, XU G R, ZHANG E P, et al. The etiology of acute preistome edema disease in cultured juveniles of Apostichopus japonicus[J]. Journal of Fisheries of China, 2006, 30(3): 377-382. |
| [4] |
黄华伟. 养殖刺参(Apostichopus japonicus)腐皮综合征的发生与异养菌区系间的关系[D]. 青岛: 中国海洋大学, 2007. HUANG H W. The relationship between the occurrence of skin ulcer syndrome in cultured Apostichopus japonicus and heterotrophic bacterial flora[D]. Qingdao: Ocean University of China, 2007. |
| [5] |
陈文博, 王洪军, 李勃, 等. 养参池底泥弧菌数量变动及微生态制剂对其影响[J]. 水产养殖, 2012, 33(10): 40-43. CHEN W B, WANG H J, LI B, et al. Effect of probiotics on total numbers of vibrio in sediment of apostichopus japonicascircle[J]. Journal of Aquaculture, 2012, 33(10): 40-43. DOI:10.3969/j.issn.1004-2091.2012.10.009 |
| [6] |
林海, 周刚, 李旭光, 等. 增氧对池塘沉积物酶活性的影响[J]. 水生态学杂志, 2011, 32(5): 100-104. LIN H, ZHOU G, LI X G, et al. Effects of different aerations on sediment enzymes activity and its relations with nutrients[J]. Journal of Hydroecology, 2011, 32(5): 100-104. |
| [7] |
何义进, 周群兰, 刘勃, 等. 不同增氧方式对中华绒螯蟹养殖池塘水质的影响[J]. 渔业现代化, 2009, 36(4): 23-26. HE Y J, ZHOU Q L, LIU B, et al. Effects of different aerating to the water quality in Eriocheir sinensis ponds[J]. Fishery Modernization, 2009, 36(4): 23-26. DOI:10.3969/j.issn.1007-9580.2009.04.005 |
| [8] |
翟海佳. 不同模式养殖池塘沉积物微生物群落结构分析[D]. 宁波: 宁波大学, 2012. ZHAI H J. Study on sediment microbial community structure in different model culture ponds[D]. Ningbo: Ningbo University, 2012. |
| [9] |
王祖峰. 仿刺参养殖池塘三种水质控制技术效果的比较[D]. 大连: 大连海洋大学, 2016. WANG Z F. A comparative study of three water quality controlling technologies effected in Apostichopus japonicus pond-aquaculture[D]. Dalian: Dalian Ocean University, 2016. |
| [10] |
CHEN J X. Present status and prospects of sea cucumber industry in China[M]//LOVATELLI A, CONAND C, PURCELL S, et al. Advances in Sea Cucumber Aquaculture and Management. Rome: FAO, 2004: 25-38.
|
| [11] |
林青, 张东升, 魏亚南, 等. 3种水质调控方式对刺参池塘浮游植物种群结构的影响[J]. 大连海洋大学学报, 2019, 34(4): 566-572. LIN Q, ZHANG D S, WEI Y N, et al. Effects of three water quality regulation methods on phytoplankton population structure in a sea cucumber culture pond[J]. Journal of Dalian Ocean University, 2019, 34(4): 566-572. |
| [12] |
魏亚南, 张东升, 林青, 等. 3种水质调控方式下参池沉积物酶活性的比较研究[J]. 水产科学, 2020, 39(2): 193-199. WEI Y N, ZHANG D S, LIN Q, et al. Effect of three types of water quality control methods on sediment enzyme activity in sea cucumber pond[J]. Fisheries Science, 2020, 39(2): 193-199. |
| [13] |
张扬. 不同盐度、底泥及扰动条件下再生水补水景观水体水质变化[D]. 西安: 西安建筑科技大学, 2017. ZHANG Y. Effect of different salinity, sediment and change rule on water quality variations of the pond replenished by reclaimed water[D]. Xi'an: Xi'an University of Architecture and Technology, 2017. |
| [14] |
顾海涛, 刘兴国, 何雅萍, 等. 微孔曝气式增氧机的性能及应用效果[J]. 渔业现代化, 2017, 44(3): 25-28. GU H T, LIU X G, HE Y P, et al. Performance and application effects of microporousaerators[J]. Fishery Modernization, 2017, 44(3): 25-28. DOI:10.3969/j.issn.1007-9580.2017.03.004 |
| [15] |
张春雪. 精养池塘底泥—水界面特征分析及底泥改良研究[D]. 武汉: 华中农业大学, 2013. ZHANG C X. The research of characteristic in water-sediment and remediation in intensive ponds sediment[D]. Wuhan: Huazhong Agricultural University, 2013. |
| [16] |
孙金凤. 增氧机扰动对池塘水体悬浮物、溶氧时空分布格局的影响[D]. 武汉: 华中农业大学, 2012. SUN J F. The impact of aerator perturbations on temporal and spatial distribution pattern of suspended matter and dissolved oxygen of pond water[D]. Wuhan: Huazhong Agricultural University, 2012. |
| [17] |
鲁晓倩. 三种水质调控方式下刺参池塘浮游植物群落结构及初级生产力的初步研究[D]. 大连: 大连海洋大学, 2017. LU X Q. A preliminary study on the community structure and primary productivity of phytoplankton in sea cucumber ponds under three water quality control methods[D]. Dalian: Dalian Ocean University, 2017. |
| [18] |
党子乔. 三种水质调控方式对海参池塘沉积物-水界面N、P通量的周年变化影响[D]. 大连: 大连海洋大学, 2017. DANG Z Q. The effect of three water quality control methods on the annual changes of N and P fluxes at the sediment-water interface of sea cucumber ponds[D]. Dalian: Dalian Ocean University, 2017. |
| [19] |
孙广伟, 周玮, 党子乔, 等. 3种水质调控方式下刺参池塘初级生产力的周年变化[J]. 水产学报, 2020, 44(4): 632-641. SUN G W, ZHOU W, DANG Z Q, et al. Annual change of primary productivity in Apostichopus japonicus ponds by three water quality regulations[J]. Journal of Fisheries of China, 2020, 44(4): 632-641. |
| [20] |
TIAN J W, FAN L, LIU H D, et al. A nearly uniform distributional pattern of heterotrophic bacteria in the Mariana Trench interior[J]. Deep Sea Research Part Ⅰ: Oceanographic Research Papers, 2018, 142: 116-126. DOI:10.1016/j.dsr.2018.10.002 |
| [21] |
田相利, 郑瑶瑶, 柳炳俊, 等. 草鱼混养系统细菌数量变动和群落功能多样性研究[J]. 中国海洋大学学报, 2012, 42(11): 19-27. TIAN X L, ZHENG Y Y, LIU B J, et al. Abundance dynamics and community functional diversity of bacteria in grass carp polyculture systems[J]. Periodical of Ocean University of China, 2012, 42(11): 19-27. |
| [22] |
迟爽, 刘海军, 刘冉, 等. 刺参养殖池塘底泥理化指标和细菌数量变化的检测[J]. 渔业科学进展, 2014, 35(1): 111-117. CHI S, LIU H J, LIU R, et al. Variation of physicochemical indices and bacterial numbers in sediment of Apostichopus japonicus culture ponds[J]. Progress in Fishery Sciences, 2014, 35(1): 111-117. DOI:10.3969/j.issn.1000-7075.2014.01.016 |
| [23] |
柳炳俊. 草鱼混养系统细菌数量变动与区系组成研究[D]. 青岛: 中国海洋大学, 2011. LIU B J. Studies on the bacterial quantity fluctuation and flora in grass carp polyculture system[D]. Qingdao: Ocean University of China, 2011. |
| [24] |
李彬, 荣小军, 廖梅杰, 等. 冬季刺参养殖环境与肠道内细菌菌群的研究[J]. 海洋科学, 2010, 34(4): 64-69. LI B, RONG X J, LIAO M J, et al. Bacteria community in the intestine and culture environment of Apostichopus japonicus in winter[J]. Marine Sciences, 2010, 34(4): 64-69. |
| [25] |
白洁, 时瑶, 宋亮, 等. 黄海西北部浮游细菌生物量分布特征及其与环境因子的关系[J]. 中国海洋大学学报, 2009, 39(4): 592-596. BAI J, SHI Y, SONG L, et al. Distribution character of bacterioplankton biomass and their relationship with environmental factors in the northwest of the Yellow Sea[J]. Periodical of Ocean University of China, 2009, 39(4): 592-596. |
| [26] |
贾俊涛, 吕艳, 李筠. 对虾围隔生态系底泥中细菌数量动态研究[J]. 动物医学进展, 2003, 24(4): 76-78. JIA J T, LYU Y, LI Y. The quantity dynamics of bacteria in the sediment of shrimp pond[J]. Progress in Veterinary Medicine, 2003, 24(4): 76-78. DOI:10.3969/j.issn.1007-5038.2003.04.023 |
| [27] |
郑晓叶, 郑天伦, 曹飞飞, 等. 不同密度青蛤养殖塘异养细菌和弧菌的数量变化[J]. 浙江农业学报, 2018, 30(10): 1647-1654. ZHENG X Y, ZHENG T L, CAO F F, et al. Quantity dynamics of heterotrophic bacteria and vibrio in cyclina sinensis rearing ponds[J]. Acta Agriculturae Zhejiangensis, 2018, 30(10): 1647-1654. DOI:10.3969/j.issn.1004-1524.2018.10.05 |
| [28] |
夏斌, 马绍赛, 崔毅, 等. 黄海绿潮(浒苔)暴发区温盐、溶解氧和营养盐的分布特征及其与绿潮发生的关系[J]. 渔业科学进展, 2009, 30(5): 94-101. XIA B, MA S S, CUI Y, et al. Distribution of temperature, salinity, dissolved oxygen, nutrients and their relationships with green tide in Enteromorpha prolifera outbreak area of the Yellow Sea[J]. Progress in Fishery Sciences, 2009, 30(5): 94-101. DOI:10.3969/j.issn.1000-7075.2009.05.016 |
| [29] |
乔方利, 马德毅, 朱明远, 等. 2008年黄海浒苔爆发的基本状况与科学应对措施[J]. 海洋科学进展, 2008, 26(3): 409-410. QIAO F L, MA D Y, ZHU M Y, et al. The basic situation and scientific countermeasures of the outbreak of Enteromorpha Yellow Sea in 2008[J]. Advances in Marine Science, 2008, 26(3): 409-410. DOI:10.3969/j.issn.1671-6647.2008.03.019 |
| [30] |
牛宇峰. 刺参养殖池塘异养菌数量变动及区系组成的初步研究[D]. 青岛: 中国海洋大学, 2009. NIU Y F. The primary studies on the bacteria flora in sea cucumber ponds[D]. Qingdao: Ocean University of China, 2009. |
| [31] |
SUI X L. The progress and prospects of studies on artificial propagation and culture of the sea cucumber, Apostichopusjaponicus[M]//LOVATELLI A. Advances in Sea Cucumber Aquaculture and Management. Rome: FAO, 2004: 273-276.
|
| [32] |
孙明洁, 张娜, 徐善良, 等. 两种弧菌感染大黄鱼免疫相关基因的SNP位点分析[J]. 上海海洋大学学报, 2019, 28(5): 772-781. SUN M J, ZHANG N, XU S L, et al. Analysis of SNP loci in immune-related genes of two species of vibrio infecting large yellow croaker (Larimichthyscrocea)[J]. Journal of Shanghai Ocean University, 2019, 28(5): 772-781. |
| [33] |
周劲风, 温琰茂, 李耀初. 养殖池塘底泥-水界面营养盐扩散的室内模拟研究: Ⅰ氮的扩散[J]. 农业环境科学学报, 2006, 25(3): 786-791. ZHOU J F, WEN Y M, LI Y C. Nutrients diffusion at the water-sediment interface of farming ponds by indoor simulation experiment: Ⅰdiffusion of nitrogenous compounds[J]. Journal of Agro-Environment Science, 2006, 25(3): 786-791. DOI:10.3321/j.issn:1672-2043.2006.03.048 |
| [34] |
张艳, 李秋芬, 王印庚, 等. 益生菌的研究现状及其在海水养殖中的应用[J]. 海洋水产研究, 2005, 26(6): 83-87. ZHANG Y, LI Q F, WANG Y G, et al. Research status and application of probiotics to mariculture[J]. Marine Fisheries Research, 2005, 26(6): 83-87. |
| [35] |
王亚南, 彭志英, 刘双江. 养殖场底泥中芽孢杆菌属细菌的生态学研究[J]. 湛江海洋大学学报, 2004, 24(4): 23-27. WANG Y N, PENG Z Y, LIU S J. Ecological study of bacillus in sediments of marine-cultural pond[J]. Journal of Zhanjiang Ocean University, 2004, 24(4): 23-27. |
| [36] |
关晓燕, 周遵春, 陈仲, 等. 应用PCR-DGGE指纹技术分析高温季节仿刺参养殖水环境中菌群多样性[J]. 海洋湖沼通报, 2010(1): 82-88. GUAN X Y, ZHOU Z C, CHEN Z, et al. Bacteria diversity analysis on the water of sea cucumber(Apostichopus japonicus)culture pond under high-temperature season based on PCR-DGGE fingerprint technology[J]. Transactions of Oceanology and Limnology, 2010(1): 82-88. |
| [37] |
王姣姣, 李丹, 宋坚, 等. 不同时期刺参养殖池塘海水菌群结构分析[J]. 中国农业科技导报, 2015, 17(2): 134-140. WANG J J, LI D, SONG J, et al. Structural analysis of seawater bacterial community from culture pond of Apostichopus japonicus at different periods[J]. Journal of Agricultural Science and Technology, 2015, 17(2): 134-140. |
| [38] |
李建松, 王广军, 龚望宝, 等. 草鱼养殖池塘蓝藻暴发时水体细菌群落特征分析[J]. 上海海洋大学学报, 2016, 25(4): 541-550. LI J S, WANG G J, GONG W B, et al. Characteristics of bacterial community of the grass carp pond when cyanobacterial blooming occurred[J]. Journal of Shanghai Ocean University, 2016, 25(4): 541-550. |