 2022,
Vol. 31
2022,
Vol. 31

2. 江苏省海洋水产研究所,江苏 南通 226000
潮间带是海岸带的重要组成部分,在潮汐作用下,潮间带频繁受到变化较大的物理和化学因素的交替影响,同时潮间带也是受到人类活动影响较严重的区域[1]。黄海海岸线长,海岸类型多样,多为粉沙淤泥质海岸,并且南黄海滩涂区域位于北亚热带与暖温带的过渡地带,从北到南,潮间带气候、资源类型、潮间带环境以及沿岸社会经济都有显著的差异,造成潮间带生物资源利用的多样性和地域分布的差异性[2]。大型底栖动物是潮间带生态系统的重要组成部分,对生态系统的稳定与平衡起着重要的作用。与此同时,大型底栖动物的群落组成与结构变化对自然环境和人类干扰极为敏感,某些环节动物是监测海岸带生态环境变化的良好指示生物[3]。相关学者对黄海南部沿海潮间带开展了大型底栖动物群落组成、生态位、物种多样性、次级生产力、功能群等方面的研究[4-7],除20世纪80年代江苏省海岸资源调查外,对黄海南部潮间带大型底栖动物的群落组成和多样性研究相对较少。本文于2019年春、夏、秋3个季节在黄海南部沿海滩涂断面实地采样调查,研究区域北起赣榆枳汪南至启东连兴港,研究潮间带大型底栖动物的群落组成和多样性的时空变化,为黄海南部浅海潮间带大型底栖动物生态特征及海岸带生态环境变化的研究提供基础资料。
1 材料与方法 1.1 调查站位和时间2019年5月(春季)、8月(夏季)、10月(秋季)在南黄海潮间带开展大型底栖动物调查。从北往南依次是在赣榆枳汪、连云港东连岛、滨海废黄河口、射阳电厂、大丰竹港、东台梁垛河、东台条子泥北、南通新港、启东东元、启东连兴港,每个断面设高潮区、中潮区、低潮区3个站点,共计30个站点(图 1)。

|
图 1 南黄海潮间带断面图 Fig. 1 Cross section of intertidal zone in southern Yellow Sea |
潮间带大型底栖动物调查多选择在大潮退潮时进行,采样时用GPS定位,每次调查分别在断面的高、中、低潮区利用0.25 m×0.25 m×0.30 m的定量框采集,每个点采集4个平行样,采用1.0 mm网目的过筛器淘洗泥样,将全部生物装入编号的样品瓶中,用体积浓度为70%的乙醇固定后带回实验室,进行种类鉴定、计数、称重,操作均按照《海洋监测规范第7部分:近海污染生态调查和生物监测(GB 17378.7—2007)》要求,并对所获得数据进行统计分析。
1.3 数据分析处理潮间带数据通过Excel软件处理后,应用Shannon-Wiener(H′)多样性指数[8],Margalef丰富度指数(d) [9],Peilou均匀度指数(J) [10]等参数处理分析。各参数的计算公式如下:
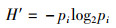 (1)
(1)
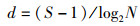 (2)
(2)
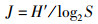 (3)
(3)
式中:Pi为第i种的数量;S为种类数;N为采泥样品中所有种类的总个体数。
采用相对重要性指数(IRI)[11]作为研究潮间带生物在群落中所占的重要性:
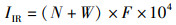 (4)
(4)
式中:IIR为相对重要性指数;N为某一种的个数占总数的百分比;W为某一种的质量占总质量的百分比;F为某一种出现的站次数占调查总站数的百分比。规定IRI大于1 000为优势种,100~1 000为主要种。大型底栖动物时空分布差异性分析使用IBM SPSS Statistics 22软件包进行聚类分析和多维尺度分析(MDS),典型种和分歧种计算运用Primer 5.0软件包。
2 结果与分析 2.1 种类组成及分布2019年5—10月3个季节10个断面共采集到潮间带大型底栖动物9门103种。其中:环节动物46种,占全部物种的44.66%;软体动物34种,占33.01%;甲壳动物15种,占14.56%;腕足动物、刺胞动物和棘皮动物动物各2种,各占1.94%;纽形动物和脊索动物各1种,各占0.97%。可见环节动物、软体动物和甲壳动物是组成南黄海潮间带大型底栖动物的主要类群。物种组成在季节和空间上均存在差异,夏季种类数最多(70种),春季次之(59种),秋季最少(56);枳汪物种数最多(43种),竹港最少(13种),见表 1和图 2。

|
表 1 南黄海潮间带大型底栖动物种类组成 Tab.1 Species composition of macrobenthos in intertidal zone of southern Yellow Sea |

|
图 2 黄海南部潮间带大型底栖动物种类组成与分布 Fig. 2 Species composition and distribution of Macrobenthos in intertidal zone of southern Yellow Sea |
南黄海潮间带各断面平均生物量为197.08 g/m2,平均生物密度为122 ind./m2,其中: 春季平均生物量为171.76 g/m2,平均生物密度为124 ind./m2;夏季平均生物量为244.97 g/m2,平均生物密度为115 ind./m2;秋季平均生物量为174.51 g/m2,平均生物密度为126 ind./m2;各潮带平均生物量以夏季最高,秋季次之,春季最低。各潮带生物密度以秋季最高,春季次之,夏季最低。
南黄海北部连云港和枳汪两断面各站位平均生物量为651.67 g/m2,平均生物密度为358 ind./m2。南黄海中部滨海港、射阳电厂等断面各站位平均生物量为49.50 g/m2,平均生物密度为60 ind./m2。南黄海南部连兴港、东元、梁垛河等断面各站位平均生物量为117.38 g/m2,平均生物密度为65 ind./m2。南黄海北部两个断面平均生物量和生物密度最高,中部平均生物量和生物密度均低于北部和南部。见表 2。

|
表 2 不同季节潮间带生物量与生物密度的时空变化 Tab.2 Spatiotemporal changes of biomass and density in different seasons and tidal zone |
通过计算得到南黄海潮间带各断面大型底栖动物群落Shannon-Wiener(H′)生物多样性指数为1.17~2.91,平均为2.14。丰度指数(d)为0.85~2.13,平均为1.36。均匀度指数(J)为0.57~1.84,平均为0.86;东连岛多样性水平最高,东元多样性最低。整体上(表 3),北部(2.57)群落多样性水平高于南部(1.94)和中部(2.12),南黄海潮间带大型底栖动物多样性指数一般,潮间带大型底栖动物群落物种丰富度较高,个体分布比较均匀。多样性指数夏季(2.33)高于春季(2.08)和秋季(2.01);丰度指数夏季(1.51)高于春季(1.35)和秋季(1.22);均匀度指数春季(0.92)也高于夏季(0.85)和秋季(0.81)。

|
表 3 南黄海潮间带大型底栖动物各断面多样性指数 Tab.3 Diversity index of macrobenthos in different sections of intertidal zone in southern Yellow Sea |
本文将优势度IRI≥1 000定位优势种,100~1 000为主要种[11-12],南黄海潮间带大型底栖动物没有IRI≥1 000的优势种,优势度在100~1 000的主要种有四角蛤蜊、文蛤、加州齿吻沙蚕、泥螺和长吻沙蚕等(表 4)。春季主要种为环节动物和软体动物,到夏季温度升高,主要种为软体动物、环节动物、腕足动物和甲壳动物,秋季主要种为软体动物、环节动物和腕足动物。

|
表 4 南黄海潮间带各季节潮间带大型底栖动物相对重要指数 Tab.4 Relative importance index of macrobenthos in intertidal zone of southern Yellow Sea |
四角蛤蜊(Mactra veneriformis)是南黄海潮间带的主要经济品种,分布范围广,除东连岛和连兴港外均有调查到,且主要分布于中潮区和低潮区。四角蛤蜊平均生物量为16.01 g/m2,占潮间带总平均生物量的8.12%,中潮区平均生物量最高为29.63 g/m2,低潮区平均生物量为10.51 g/m2,高潮区平均生物量最低为7.89 g/m2;各季节中夏季平均生物量最高,为23.35 g/m2,春季生物量最低,为9.03 g/m2。夏季东元断面中潮区四角蛤蜊生物量最高,为286.37 g/m2, 见图 3。

|
图 3 南黄海近岸潮间带四角蛤蜊生物量水平分布 Fig. 3 Horizontal biomass distribution of mactraveneriformis in intertidal zone of southern Yellow Sea |
聚类分析图和多维尺度分析图见图 4,典型种和分歧种见表 5。聚类图显示按欧氏距离10%可划分为3个类群:第一个类群包括东连岛和枳汪的3个潮带,站点位于南黄海北部,其海岛潮间带生境多样性较高,大型底栖动物种类较多,生物密度和生物量最高,属于泥沙质潮间带,其典型种包括双齿围沙蚕、疣荔枝螺、史氏背尖贝、圆球股窗蟹、褶牡蛎、菲律宾蛤仔和中华近方蟹,对组内平均相似性的贡献累计达到72%。第二个类群包括东元、新港、连兴港和梁垛河,都位于南黄海南部,同时为江苏省文蛤、泥螺等经济贝类的重要增养殖水域,密度稍低于第一个群,典型种包括文蛤、海豆芽、泥螺、长吻沙蚕、加州齿吻沙蚕、浅古铜吻沙蚕和宽身大眼蟹,对组内平均相似性贡献率累计达到75.14%。第三个类群以滨海港、竹港、射阳电厂和条子泥北的潮带为主,为粉砂淤泥质岸滩,该群内生物量和生物密度低于前两个群,其典型种包括加州齿吻沙蚕、四角蛤蜊、浅古铜吻沙蚕、彩虹明樱蛤、托氏螺、泥螺和长锥虫,对组内相似性贡献率累计达到71.26%。3个区域的典型种各不相同,北部和中部组间分歧种主要为加州齿吻沙蚕、四角蛤蜊、泥螺、浅古铜吻沙蚕、彩虹明樱蛤和褶牡蛎等,对组间相异性贡献率达到52.79%;中部和南部组间分歧种主要为加州齿吻沙蚕、四角蛤蜊、托氏螺、琥珀刺沙蚕、宽身大眼蟹、泥螺、长吻沙蚕、海豆芽和文蛤,对组间相异性贡献率达到45.91%;北部和南部组间分歧种主要为加州齿吻沙蚕、双齿围沙蚕、海豆芽、文蛤、长吻沙蚕、圆球股窗蟹和褶牡蛎,对组间相异性贡献率为32.14%。

|
BH.滨海港;ZW.枳汪;DLD.东连岛;SYDC.射阳电厂;ZG.竹港;LDH.梁垛河;XG.新港;TZNB.条子泥北;DY.东元;LXG.连兴港;Spr.春季;Sum.夏季;Aut.秋季;右图虚线显示MDS分析划分的3个主要的群。 Province BH.Binhai port; ZW.Zhiwang; DLD.Donglian Island; SYDC.Sheyang power plant; ZG.Zhugang; LDH.Liangduo River; XG.Xinggang; TZNB.TiaoziNibei; Dy.Dongyuan; LXG.Lianxing port; Spr.spring; Sum.summer; Aut.autumn; The dotted line on the right shows the three main groups divided by MDS analysis. 图 4 南黄海潮间带大型底栖动物群落系统聚类(左)及MDS分析图(右) Fig. 4 Cluster analysis(right) and MDS analysis(left) of macrobenthos community in intertidal zone of southern Yellow Sea |

|
表 5 南黄海潮间带大型底栖动物各区域典型种和分歧种 Tab.5 Typical and divergent species of macrobenthos in different regions of intertidal zone in southern Yellow Sea |
从MDS排序图中不同程度的潮间带分布可以看出,MDS的排序结果与SPSS聚类分析的结果一致,南黄海潮间带大型底栖动物群落结构空间上的变化差异大于季节上的差异。
3 讨论 3.1 南黄海底栖动物群落结构的空间变化分析2019年3个季节大型底栖动物调查结果显示,南黄海北部大型底栖动物群落生物量、生物密度和多样性水平高于南部和中部。造成大型底栖动物群落空间差异的主要因素为底质类型[13-14],其中: 北部东连岛具有独特的海岛环境,岩石和砂质底质条件为甲壳动物和软体动物提供了优质的生境,主要典型种为疣荔枝螺、褶牡蛎和圆球股窗蟹等;中部滨海港多为粉砂淤泥质底质,潮间带地势平坦,但近年来海上风电项目开发、沿海滩涂围垦[15]和环境污染等人类活动导致了当地底栖动物的物种数、生物量和生物密度远低于东连岛和新港,对环境变化不太敏感的环节动物成为了中部的典型种;南部新港沿岸地形低平,高潮区土质坚实,表层为粉砂黏土层,下部为砂层及粉砂土层,低潮区为粉砂带,该生境适宜文蛤和泥螺等软体动物生存。不同的底质条件和生境造成南黄海大型底栖动物群落结构空间上的差异,北部和南部的潮间带更适合大型底栖动物的生存繁衍。
3.2 南黄海底栖动物群落结构的季节变化分析南黄海2019年大型底栖动物物种数、生物量和生物密度变化不同: 底栖动物种类数季节变化顺序为夏季>春季>秋季;生物量夏季高于秋季,春季最低;生物密度秋季高于春季,夏季最低。南黄海潮间带自然环境的特点是,既属亚热带的性质,又兼备温带的某些特征。因此在潮间带动物的种类组成中,属于南方的变性暖水种范畴的有:泥螺、褶牡蛎、文蛤、长竹蛏、圆球股窗蟹、双齿围沙蚕等[16]。在食物网中,软体动物营养级随季节变化不大,3个季节主要种中软体动物优势度较高;优势种计算中,春季软体动物和环节动物为主要种,到夏季软体动物依然占主要优势;甲壳动物优势度在夏季高于环节动物,可能是其存在种间竞争,夏季温度较高,甲壳类快速生长,需要大量摄食,由于环节动物行动缓慢,竞争能力弱于甲壳动物,而且春季环节动物摄食的物种多为早期发育阶段的小型生物个体,活动能力不强,随着这些物种的生长发育,夏季活动能力加强,不易被环节动物捕食[17]。夏季个体较大,所以生物量在高于春季和秋季的情况下,生物密度低于春季和秋季。1982年南黄海潮间带大型底栖动物生物量和生物密度季节变化是秋季大于春季,与2019年潮间带底栖动物结构季节变化趋势相近,但是生物量低于2019年调查结果。该结果可能是采样手段、调查海域、调查时间的不同导致。对比季节变化发现,断面季节上的差异小于空间分布上的差异。
3.3 南黄海潮间带群落结构与历史资料比较分析与相近海域历史资料相比,本研究南黄海北部和南部的潮间带生物量和生物多样性较历史资料有所增加。连云港赣榆和东连岛泥沙质滩涂平均生物量和物种数为651.67 g/m2和61种;生物量高于连云港海州湾2014年李军等[18]研究的66.34 g/m2,物种数(61种)高于海州湾(57种),物种多样性指数与海州湾(3.26)差异较小,南黄海北部区域生物量和物种数高于历史资料,其主要种都为软体动物和甲壳动物。相对而言南黄海北部海岛潮间带地形变化较大,生境丰富,所以底栖动物多样性较高,生物量高于江苏中部和南部。与张虎等[19]2014年江苏中南部相比,本研究南黄海南部连兴港、新港、梁垛河、东元物种数为67种与其68种相差较小,生物密度和生物量为65.35 ind./m2和117.37 g/m2,两次研究相差较小(65.51 ind./m2、94.31 g/m2),但物种多样性高于江苏中南部(1.78),群落主要种都为软体动物和环节动物。样品采集时间、站点分布和环境因素变化等诸多方面造成该结果。
3.4 南黄海潮间带群落结构与其他海域比较本次南黄海潮间带3个季节10个断面共采集大型底栖动物103种,优势种以软体动物和环节动物为主。物种数和生物量远高于陶世如等[20]研究的长江口横沙岛和长兴岛潮间带(13种和37.95 g/m2),调查中出现的优势种多以甲壳动物和软体动物为主;物种数低于张宝琳等[21-24]研究的胶州湾幸岛潮间带大型底栖动物的物种数(199种),但生物量高于张宝琳等调查得到的生物量(60.01 g/m2),胶州湾调查中出现的优势种多以环节动物为主;物种数高于冯明等[25]研究的浙江韭山列岛岩相潮间带大型底栖动物的物种数(25种),但生物量低于冯明等研究得到的生物量(1 228.4 g/m2),浙江韭山列岛调查中出现的优势种多为软体动物。南黄海位于长江口、浙江韭山列岛和胶州湾之间区域,多样的底质类型和跨越3个纬度的空间距离等诸多因素造成其与周边海域潮间带结构存在差异性。
4 结论通过对南黄海潮间带大型底栖动物群落的调查和分析,结果表明:共调查发现潮间带生物9门103种,其中夏季物种数最多,春季次之,秋季最少。夏季平均生物量高于春季和秋季,秋季平均生物密度高于春季和夏季,南黄海潮间带大型底栖动物群落结构在季节间存在显著的差异。黄海南部潮间带大型底栖动物多样性指数一般,潮间带大型底栖动物群落物种丰富度较高,个体分布比较均匀。通过聚类分析和多维尺度分析方法对10个断面进行相似性比较,可将10个断面划分为北部、中部和南部3个区域进行空间上比较,北部平均生物量高于南部和中部,北部平均生物密度高于南部和中部,且北部群落多样性水平高于南部和中部,南黄海潮间带大型底栖动物群落结构空间上存在的差异大于季节上的差异。本研究可为南黄海浅海潮间带大型底栖动物生态特征及海岸带生态环境变化的研究提供基础资料。
| [1] |
胡桂坤, 秦璐璐, 李郁郁, 等. 基于ABC曲线的天津潮间带生物群落受扰动的分析[J]. 天津科技大学学报, 2019, 34(5): 57-62. HU G K, QIN L L, LI Y Y, et al. Disturbance status of intertidal macrobenthos in Tianjin based on ABC curves[J]. Journal of Tianjin University of Science & Technology, 2019, 34(5): 57-62. |
| [2] |
刘群, 殷勇. 江苏沿海滩涂地貌及资源开发利用途径[J]. 河南科学, 2010, 28(11): 1482-1490. LIU Q, YIN Y. The tidal flat geomorphology of Jiangsu coast area and ways of resources development and exploitation[J]. Henan Science, 2010, 28(11): 1482-1490. DOI:10.3969/j.issn.1004-3918.2010.11.030 |
| [3] |
王金宝, 李新正, 王洪法, 等. 山东长岛潮间带大型底栖动物生态特征研究[J]. 海洋科学, 2018, 42(10): 41-52. WANG J B, LI X Z, WANG H F, et al. Macrobenthic ecology in the interridal zones of Changdao Islands in Shandong Province[J]. Marine Sciences, 2018, 42(10): 41-52. DOI:10.11759/hykx20180803002 |
| [4] |
袁健美, 张虎, 汤晓鸿, 等. 江苏南部潮间带大型底栖动物功能群研究[J]. 海洋渔业, 2019, 41(1): 43-52. YUAN J M, ZHANG H, TANG X H, et al. Macrozoobenthic functional groups in intertidal zone of southern Jiangsu province[J]. Marine Fisheries, 2019, 41(1): 43-52. DOI:10.3969/j.issn.1004-2490.2019.01.006 |
| [5] |
袁健美, 张虎, 汤晓鸿, 等. 江苏潮间带大型底栖动物群落组成及次级生产力[J]. 生态学杂志, 2018, 37(11): 3357-3363. YUAN J M, ZHANG H, TANG X H, et al. Community structure and secondary production of macrobenthos in intertidal zone of Jiangsu[J]. Chinese Journal of Ecology, 2018, 37(11): 3357-3363. |
| [6] |
张虎, 汤晓鸿, 郭仲仁, 等. 江苏中南部潮间带大型底栖动物时空分布特征[J]. 海洋渔业, 2014, 36(3): 208-215. ZHANG H, TANG X H, GUO Z R, et al. Temporal and spatial distribution of benthic macrofauna in intertidal mudflat of central and southern Jiangsu coast[J]. Marine Fisheries, 2014, 36(3): 208-215. DOI:10.3969/j.issn.1004-2490.2014.03.003 |
| [7] |
童远瑞, 孟文新. 江苏沿海潮间带的贝类区系研究[J]. 南京大学学报(自然科学版), 1985, 22(1): 111-119. TONG Y R, MENG W X. Studies on molluscan fauna of Jiangsu coastal sea[J]. Journal of Nanjing University (Natural Sciences), 1985, 22(1): 111-119. |
| [8] |
SHANNON CE, WEAVER W. The mathematical theory of communication[J]. Physics Today, 1950, 3(9): 31-32. DOI:10.1063/1.3067010 |
| [9] |
MARGALEFR. Temporal succession and spatial heterogeneity inphytoplankton[M]//BUZZATI-TRAVERSO A A. Perspectives in Marine Biology. Berkeley: University of California Press, 1958: 323-349.
|
| [10] |
PIELOU EC. Ecological diversity[M]. New York: John Wiley, 1975: 16-51.
|
| [11] |
江海华, 彭欣, 肖国强, 等. 温岭海岛潮间带大型底栖生物的群落结构[J]. 海洋学研究, 2014, 32(3): 78-86. JIANG H H, PENG X, XIAO G Q, et al. Community structure of intertidal macrobenthos in islands of Wenling, Zhejiang, China[J]. Journal of Marine Sciences, 2014, 32(3): 78-86. DOI:10.3969/j.issn.1001-909X.2014.03.011 |
| [12] |
王航俊, 邹清, 郜钧璋, 等. 玉环国家级海洋公园夏季潮间带大型底栖生物群落结构研究[J]. 海洋环境科学, 2020, 39(1): 127-132. WANG H J, ZOU Q, GAO J Z, et al. Study on the macrobenthic community structure of the intertidal zone in the Yuhuan National Marine Park in summer[J]. Marine Environmental Science, 2020, 39(1): 127-132. |
| [13] |
高爱根, 杨俊毅, 曾江宁, 等. 海州湾潮间带大型底栖动物的分布特征[J]. 海洋学研究, 2009, 27(1): 22-29. GAO A G, YANG J Y, ZENG J N, et al. Distribution of the intertidal macrobenthos in the Haizhouwan Bay[J]. Journal of Marine Sciences, 2009, 27(1): 22-29. DOI:10.3969/j.issn.1001-909X.2009.01.004 |
| [14] |
季相星, 姜毅, 王晨波, 等. 连云港市泥砂质潮间带大型底栖动物群落调查[J]. 环境监控与预警, 2014, 6(6): 41-44. JI X X, JIANG Y, WANG C B, et al. Primary study on community structure of macrobenthos in the Argillo-arenaceousintertidal zone in Lianyungang[J]. Environmental Monitoring and Forewarning, 2014, 6(6): 41-44. DOI:10.3969/j.issn.1674-6732.2014.06.012 |
| [15] |
王静, 徐敏, 张益民. 滩涂围垦养殖的生态损益分析——以江苏条子泥滩涂围垦养殖为例[J]. 南京师范大学学报(自然科学版), 2012, 35(2): 113-119. WANG J, XU M, ZHANG Y M. Ecological profit and loss analysis of tidal flat reclamation——tidal flat reclamation of Tiaozini Sand as a case study[J]. Journal of Nanjing Normal University (Natural Science Edition), 2012, 35(2): 113-119. |
| [16] |
王颖. 南黄海辐射沙脊群环境与资源[M]. 北京: 海洋出版社, 2014. WANG Y. Environment and resources of radial sand ridges in the South Yellow Sea[M]. Beijing: Ocean Press, 2014. |
| [17] |
刘春云, 姜少玉, 宋博, 等. 烟台养马岛潮间带大型底栖动物食物网结构特征[J]. 海洋与湖沼, 2020, 51(3): 467-476. LIU C Y, JIANG S Y, SONG B, et al. Food web structure of macrobenthos in the intertidal zone of Yangma Island, Yantai, China[J]. Oceanologia et Limnologia Sinica, 2020, 51(3): 467-476. |
| [18] |
李军, 季相星, 姜玲, 等. 海州湾潮间带大型底栖动物多样性研究[J]. 环境监控与预警, 2017, 9(4): 45-48. LI J, JI X X, JIANG L, et al. Biodiversity of macrobenthos in the intertidal zone of Haizhou Bay[J]. Environmental Monitoring and Forewarning, 2017, 9(4): 45-48. |
| [19] |
张虎, 刘培廷, 汤建华, 等. 南通市大型海洋排污海区潮间带底栖生物研究[J]. 水产科学, 2009, 28(7): 409-412. ZHANG H, LIU P T, TANG J H, et al. Intertidal macrobenthos in large marine sewage waters in Nantong[J]. Fisheries Science, 2009, 28(7): 409-412. DOI:10.3969/j.issn.1003-1111.2009.07.012 |
| [20] |
陶世如, 姜丽芬, 吴纪华, 等. 长江口横沙岛、长兴岛潮间带大型底栖动物群落特征及其季节变化[J]. 生态学杂志, 2009, 28(7): 1345-1350. TAO S R, JIANG L F, WU J H, et al. Community characteristics and seasonal changes of macrozoobenthos in intertidal zones of Hengsha and Changxing Islands at Yangtze River estuary[J]. Chinese Journal of Ecology, 2009, 28(7): 1345-1350. |
| [21] |
张宝琳, 王洪法, 李宝泉, 等. 胶州湾辛岛潮间带大型底栖动物生态学调查[J]. 海洋科学, 2007, 31(1): 60-64. ZHANG B L, WANG H F, LI B Q, et al. The ecology of the macrobenthic community in the intertidal zone of Xindao, Jiaozhou Bay[J]. Marine Sciences, 2007, 31(1): 60-64. |
| [22] |
王洪法, 李宝泉, 张宝琳, 等. 胶州湾红石崖潮间带大型底栖动物群落生态学研究[J]. 海洋科学, 2006, 30(9): 52-57. WANG H F, LI B Q, ZHANG B L, et al. The ecological research of the macrobenthic community in intertidal zone of Hongshiya, Jiaozhou Bay[J]. Marine Sciences, 2006, 30(9): 52-57. |
| [23] |
李宝泉, 张宝琳, 刘丹运, 等. 胶州湾女姑口潮间带大型底栖动物群落生态学研究[J]. 海洋科学, 2006, 30(10): 15-19. LI B Q, ZHANG B L, LIU D Y, et al. The ecological study of the macrobenthic community in interdidal zone of Nugukou, Jiaozhou Bay[J]. Marine Sciences, 2006, 30(10): 15-19. |
| [24] |
李新正, 李宝泉, 王洪法, 等. 胶州湾潮间带大型底栖动物的群落生态[J]. 动物学报, 2006, 52(3): 612-618. LI X Z, LI B Q, WANG H F, et al. Macrobenthic community of the intertidal zone of Jiaozhou Bay[J]. Acta Zoologica Sinica, 2006, 52(3): 612-618. |
| [25] |
冯明, 韩庆喜, 严润玄. 浙江韭山列岛岩礁潮间带大型底栖动物群落特征和功能性状[J]. 水生态学杂志, 2021, 40(5): 1469-1477. FENG M, HAN Q X, YAN R X. Community characteristics and functional traits of macrobenthos in intertidal zone of Jiushan archipelago in Zhejiang Province[J]. Journal of Ecology, 2021, 40(5): 1469-1477. |
 ,
ZHANG Hu2
,
ZHANG Hu2 ,
YUAN Jianmei2,
SHI Jiawei2,
BEN Chengkai2,
HU Haisheng2,
XIAO Yueyue2,
ZU Kaiwei2,
TANG Xiaohong2
,
YUAN Jianmei2,
SHI Jiawei2,
BEN Chengkai2,
HU Haisheng2,
XIAO Yueyue2,
ZU Kaiwei2,
TANG Xiaohong2
2. Jiangsu Institute of Marine Fisheries, Nantong 226000, Jiangsu, China