 2021,
Vol. 30
2021,
Vol. 30

2. 南方海洋科学与工程广东省实验室南海资源大数据中心, 广东 湛江 524013;
3. 集美大学 水产学院, 福建 厦门 361021;
4. 广东省南海深远海渔业管理与捕捞工程技术研究中心, 广东 湛江 524088;
5. 广东海洋大学 深圳研究院海洋渔业信息化技术中心, 广东 深圳 518120
北部湾位于南海北部,海南岛以西(17°00′N~21°45′N和105°40′E~110°10′E)海域,为天然半封闭海湾[1],其渔业资源丰富,是我国的传统优良渔场之一[2]。近年来关于北部湾资源调查和研究[3-9]结果表明,北部湾海域渔业资源呈衰退趋势,开展以常见经济鱼类为重点的研究是制定和实施资源恢复相关政策和措施的基础。
斑鳍白姑鱼(Pennahia pawak)作为北部湾较为稳定的资源种类,具有重要的生态地位和经济效益[6]。近年来,国内不少学者对斑鳍白姑鱼的基础生物学和资源动态分布进行了研究,如王雪辉等[10]对南海北部的斑鳍白姑鱼的体长-体质量关系进行了初步分析,孙典荣等[11]初步分析了海南岛附近斑鳍白姑鱼的季节性分布,何雄波等[12]研究了斑鳍白姑鱼的种群结构、资源密度及其分布,颜云榕等[13-14]基于鳞片研究了斑鳍白姑鱼年龄特征,并对其繁殖习性和摄食习性做了分析,侯刚等[15]研究了北部湾4种白姑鱼属(Pennahia)鱼类,包括斑鳍白姑鱼在内的耳石形态识别。这些研究结果为渔业资源评估提供了基础资料,而鱼类的生长和死亡作为影响资源数量变动的基本原因之一,也是渔业资源评估必不可少的重要环节[16]。但是目前关于北部湾斑鳍白姑鱼生长与死亡等生物学参数及其变化的研究比较缺乏,仅颜云榕等[14]基于鳞片读取的年龄数据对北部湾斑鳍白姑鱼的生长参数进行了估算,有必要进一步开展和完善北部湾斑鳍白姑鱼生长和死亡等重要生物学参数的相关研究。除了鳞片和耳石等硬质样本外,通过获取具有代表性的渔获组成数据,也能对生长和死亡等重要参数进行估算[17]。FiSAT软件的体长频数分析法因方便、快捷等特点被广泛应用于鱼类生长死亡参数的估算[18],且能够较好地反映鱼类资源开发状况。
本研究根据2008—2009和2018—2019年北部湾资源调查样本,基于体长频率法,应用FiSATⅡ渔业资源评估软件,对斑鳍白姑鱼的种群结构、生长、死亡和开发率等参数进行研究,探讨北部湾斑鳍白姑鱼群体生物学参数的变化,为渔业资源合理开发和管理提供参考。
1 材料与方法 1.1 样品与数据来源样本来自于2008—2009和2018—2019年4个季度在北部湾北海和东方渔港的上岸渔获物(以底拖网为主,网目尺寸约为40 cm),采样点位置见图 1,以随机采样的方式,采集斑鳍白姑鱼样品共992尾(表 1)。按照《海洋调查规范》在实验室内测定体长和体质量等生物学指标,体长精确至0.1 cm,体质量精确至0.1 g。

|
图 1 北部湾斑鳍白姑鱼采样位点图 Fig. 1 Sampling sites of P. pawak in Beibu Gulf |

|
表 1 北部湾斑鳍白姑鱼采样信息 Tab.1 Sampling information of P. pawak in the Beibu Gulf |
体长与体质量关系采用幂函数[19]表示:
 (1)
(1)
式中:W为体质量,g;L为体长,cm;a、b为参数,a为条件因子,b为异速生长因子。b=3,表示该鱼类个体呈等速生长;b>3,表示鱼类个体呈正异速生长;b<3,表示鱼类个体呈负异速生长[20-21]。
描述斑鳍白姑鱼的生长选用von Bertalanffy生长方程[16]:
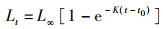 (2)
(2)
式中:t为年龄;Lt为t龄时的体长;L∞为渐近体长;K为接近渐进体长的相对速度;t0为理论起始生长的年龄。L∞和K采用FiSAT Ⅱ软件的体长频率分析(electronic length frequency analysis Ⅰ,ELEFAN Ⅰ)[22-23]方法,以1 cm为组间距获取的体长频数,拟合生长曲线进行估算。t0的估算应用PAULY[24]提出的经验公式:
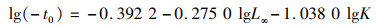 (3)
(3)
根据体长变换渔获曲线法(length-converted catch curve)估算总死亡系数(Z)[16],自然死亡系数(M)由PAULY[25]提出的经验公式进行估算:
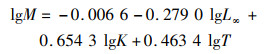 (4)
(4)
式中:T为北部湾年平均海水表层温度,2008—2009年平均海水表层温度为25.6 ℃,2018—2019年平均海水表层温度为26.2 ℃。海表面温度(sea surface temperature,SST)数据来自美国国家海洋大气局(https://oceanwatch.pifsc.noaa.gov/),选取的经纬度范围为18°N~22°N和105°E~110°E。
捕捞死亡系数(F)和开发率(E)[16]则根据总死亡系数和自然死亡系数计算,公式分别为
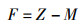 (5)
(5)
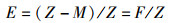 (6)
(6)
式中:F为捕捞死亡系数;E为开发率; Z为总死亡系数;M为自然死亡系数。
开捕体长(Lc)根据体长渔获曲线拟合出的直线方程进行估算。推算线性回归中未被使用的各点相对应的期望值ln(N/Δt),并计算各点的观测值与期望值的比,以及比值累积率,选取累积率达到50%的点,其所对应的体长即为平均选择体长的估计量(Lc),也称为开捕体长[26]。
2 结果 2.1 群体结构北部湾斑鳍白姑鱼2008—2009和2018—2019年的体长分别为8.3~24.6 cm和7.0~21.1 cm,平均体长分别为14.83和12.59 cm;优势体长分别为11~17 cm(75.8%)和10~15 cm(73.6%),见图 2。与2008—2009年相比,2018—2019年的体长范围变小,平均体长下降了15.1%。北部湾斑鳍白姑鱼2008—2009和2018—2019年的体质量分别为14.2~401.4 g和7.5~216.3 g,平均体质量分别为85.88和52.91 g;优势体质量分别为30~100 g(68.3%)和10~80 g(85.7%),见图 3。与2008—2009年相比,2018—2019年的体质量范围缩小,平均体质量下降了38.4%。

|
图 2 北部湾斑鳍白姑鱼体长分布频率 Fig. 2 Body length distribution of P. pawak in the Beibu Gulf |

|
图 3 北部湾斑鳍白姑鱼体质量分布频率 Fig. 3 Body mass distribution of P. pawak in the Beibu Gulf |
2008—2009年北部湾斑鳍白姑鱼个体的体长均值在冬季最高(16.05 cm)、春季(14.95 cm)和夏季(14.93 cm)次之、秋季最低(13.43 cm),其体质量均值在冬季最高(105.63 g)、夏季(86.22 g)和春季(84.50 g)次之、秋季最低(67.45 g),见图 4。

|
图 4 2008—2009年北部湾斑鳍白姑鱼体长、体质量季节分布箱线图 Fig. 4 Boxplot of body length and body mass of P. pawak in the Beibu Gulf from 2008 to 2009 |
2018—2019年北部湾斑鳍白姑鱼个体的体长均值在夏季最高(13.44 cm)、冬季(13.39 cm)和春季(12.21 cm)次之、秋季最低(11.85 cm),其体质量均值在冬季(63.67 g)最高、夏季(57.53 g)和春季(51.25 g)次之、秋季最低(43.66 g),见图 5。

|
图 5 2018—2019年北部湾斑鳍白姑鱼体长、体质量分布箱线图 Fig. 5 Boxplot of body length and body mass of P. pawak in the Beibu Gulf from 2018 to 2019 |
采用幂函数拟合分析北部湾斑鳍白姑鱼体长与体质量关系,2008—2009年体长与体质量关系式为W=0.036L2.84(R2=0.927),2018—2019年体长与体质量关系式为W=0.024L3.00(R2=0.949),见图 6。

|
图 6 北部湾斑鳍白姑鱼体长与体质量的关系 Fig. 6 Body length-body mass relationships of P. pawak in the Beibu Gulf |
通过分析不同时段斑鳍白姑鱼体长-体质量关系参数(表 2),2个时段(2008—2009和2018—2019)的条件因子a值分别为0.036和0.024,异速生长因子b值分别为2.84和3.00。2008—2009年斑鳍白姑鱼的b值在夏季最高为2.94,冬季最低为2.79,依次为夏季>秋季>春季>冬季,均小于3,表现为负异速生长;2018—2019年其b值在春季最高为3.20,夏季最低为2.63,依次为春季>秋季>冬季>夏季,其中春季和秋季的b值大于3,呈正异速生长,夏季和冬季的b值小于3,呈负异速生长;2008—2009年斑鳍白姑鱼的a值在冬季最高为0.043,依次为冬季>春季>夏季=秋季,2018—2019年斑鳍白姑鱼a值在夏季最高为0.059,春季最低0.015,依次为夏季>冬季>秋季>春季。

|
表 2 斑鳍白姑鱼的体长-体质量关系参数 Tab.2 Parameters of body length-body mass relationships of P. pawak |
利用ELEFAN方法估算斑鳍白姑鱼2个时段(2008—2009和2018—2019)的渐进体长L∞分别为24.15 cm和22.05 cm,生长速率K分别为0.39和0.32,2个时段个体的生长参数L∞和K均减小;根据公式3估算理论起始生长的年龄t0,结果分别为-0.35和-0.56(表 3)。2008—2009和2018—2019年2个时段北部湾斑鳍白姑鱼体长的von Bertalanffy生长方程分别为Lt=24.15[1-e-0.39(t+0.35)]和Lt=22.05[1-e-0.32(t+0.56)]。应用长度变换渔获曲线法进行线性回归,拟合结果:2008—2009年的线性方程为ln(N/Δt)=-1.39t+9.068,R2=0.956;2018—2019年的线性方程为ln(N/Δt)=-1.17t+8.213,R2=0.943。估算得到斑鳍白姑鱼的总死亡系数(Z即为方程的斜率)分别为1.39和1.17。根据公式4估算自然死亡系数分别为0.98和0.90,捕捞死亡系数分别为0.41和0.28,开发率分别为0.29和0.24。另外,基于变换体长渔获量曲线拟合关系进行进一步推算2个时段北部湾斑鳍白姑鱼的开捕体长,结果分别为12.56 cm和10.29 cm(图 7)。对比发现,2018—2019年北部湾斑鳍白姑鱼的总死亡系数、自然死亡系数、捕捞死亡系数、开发率以及开捕体长均减小。

|
表 3 北部湾斑鳍白姑鱼生长死亡参数对比分析 Tab.3 Comparative analysis of parameters for P. pawak in the Beibu Gulf |

|
图 7 基于变换体长渔获曲线估算斑鳍白姑鱼开捕体长 Fig. 7 Catchable size from the proportional retention of length converted catch curve |
在较大捕捞压力下,多数鱼类的表型性状会产生适应性变化,如体长和性成熟的演化。有研究表明,北部湾重要经济鱼类逐渐被低值、小型的鱼类所替代[6],并且多种经济鱼类已经出现个体小型化和性成熟提前等现象[27-30]。1997—2019年间数据对比(表 4)显示,斑鳍白姑鱼的平均体长在1997—2002和2008—2009年间增大,在2008—2009年度最大,在2010—2011和2018—2019年减小。一方面,平均体长在1997—2002和2008—2009年间增大的原因可能是1999年我国采取休渔政策等限制捕捞压力的措施取得了一定效果,同时燃油价格上涨,北部湾的捕捞强度有所下降,使鱼类资源在1999年之后近十年得到有效恢复[7]。另一方面,由于越南无休渔政策,作业渔船不断增加,多年来北部湾渔业一直承受着较高的捕捞强度[27],北部湾海域的斑鳍白姑鱼资源可能已存在衰退,小个体群体居多,反映在2010—2011和2018—2019年的群体平均体长减小。研究结果表明,2018—2019和2010—2011年各个季度平均体长、体质量均低于2008—2009年,进一步验证了斑鳍白姑鱼各个季度小个体鱼群居多。以上分析表明,1997—2019年北部湾斑鳍白姑鱼的体长范围和体质量范围都在缩小,大个体明显减少,整体上呈小型化趋势,当然也可能存在一定的采样偏差,采集了较多的小个体样本,从而对本研究结果产生一定影响。

|
表 4 北部湾斑鳍白姑鱼体长和体质量参数的年际变化 Tab.4 Interannual variation of body length and body mass parameter of P. pawak from the Beibu Gulf |
鱼类的体长-体质量关系为渔业资源评估模型提供了重要参数[16]。基于体长与体质量幂函数关系估算的生长参数a的大小在一定程度上反映鱼类生存环境的优劣[16],2个时段北部湾斑鳍白姑鱼的生长参数a由0.036下降至0.024,说明斑鳍白姑鱼的生境有所恶化。生长参数b反映了鱼类体长与体质量增长的不均匀性[31],其影响因素是多方面的,如发育的阶段[31]、种群的不同[32]、环境变化[33]、捕捞强度[10]等。北部湾斑鳍白姑鱼接近等速生长,但其生长受到各种因素的制约,会发生一定变化。2008—2009年斑鳍白姑鱼的生长参数b为2.84,与1997—1999年比较明显减小[10],可能是持续较大的捕捞压力所致。STERGIOU等[34]、王雪辉等[10]的研究曾指出在较大捕捞压力下,鱼类个体的体长生长速度增加,生长参数b有减小的趋势。2018—2019年北部湾斑鳍白姑鱼的b值为3.00,与王雪辉等[10]研究结果基本一致。因此,斑鳍白姑鱼趋向等速生长。
对比2008—2009和2018—2019年北部湾斑鳍白姑鱼各季度生长参数b,发现冬季群体均为负异速生长。与之相对应,也有研究[12]表明在冬季北部湾斑鳍白姑鱼资源密度较大,这可能与温度、饵料水平有关。2018—2019年夏季的生长参数b最低(2.63),呈负异速生长,这可能与8月休渔结束致使捕捞压力突增以及饵料状况有关。
3.3 生长、死亡及开发率生长参数K和L∞是鱼类群体结构变化的指标[35-36],对比2个时段个体的生长参数K,2018—2019年的生长速度相较于2008—2009年有略微下降,2018—2019年度斑鳍白姑鱼L∞下降了8.7%;开捕体长作为渔业资源管理参考的重要指标[16],2018—2019年实际的开捕体长也减小了18.1%,进一步表明斑鳍白姑鱼倾向小个体,这与本研究斑鳍白姑鱼种群结构小型化的结果一致。根据GULLAND[37]提出的一般鱼类最适开发率为0.5,2008—2009年开发率(0.29)大于2018—2019年开发率(0.24),且均小于0.5,表明在10年间斑鳍白姑鱼资源介于轻度开发的状态。2个时段斑鳍白姑鱼的自然死亡系数相对较高,分别为0.98和0.90,陈作志等[38]分别对1964、1992、1997年北部湾白姑鱼(Argyrosomus argentatus)的死亡参数进行了估算,其自然死亡系数为0.93~1.25,它与斑鳍白姑鱼为同属种类,本研究的斑鳍白姑鱼与白姑鱼的自然死亡系数相似。另外,2008—2009和2018—2019年的自然死亡系数分别为相应年度捕捞死亡系数的2.4倍和3.2倍,由此可推断自然死亡是斑鳍白姑鱼死亡的主要原因。自然死亡系数受鱼类本身的年龄、资源量、捕食者数量、饵料丰度和自然环境等因素影响[39]。就已有的研究来看,一方面生存环境变化及栖息地水质污染等原因可能导致自然死亡率较高,另一方面北部湾渔业长期保持较高的捕捞强度,个体较大、经济价值较高的优势种逐渐减少,低值、中小型的种类增多,其海洋生态系统营养级有下降的趋势[6]。斑鳍白姑鱼的饵料生物以小型鱼类和虾类等为主[13],这些饵料生物同时也被白姑鱼属的其他种类[40]、带鱼科鱼类[41]、多齿蛇鲻(Saurida tumbil)[42]、宝刀鱼(Chirocentrus dora)[43]、竹䇲鱼(Trachurus japonicus)[44-45]等多种鱼类摄食,种间存在较大的饵料竞争,斑鳍白姑鱼等中小型鱼类共同被更高营养级的种类所捕食,当更优质的捕食者减少时,摄食、繁殖能力更强的种类会迅速发展,从而造成斑鳍白姑鱼的饵料减少。生长和繁殖两者呈明显的负相关,当摄入的能量减小,为维持种群繁衍和稳定,用于生长的能量减少,体型将可能呈现小型化[46]。总之,饵料丰度、人类捕捞以及栖息地环境变化可能是斑鳍白姑鱼保持较高的自然死亡率和个体小型化的主要原因,由于本文研究内容有限,具体的情况有待进一步研究。
| [1] |
乔延龙, 林昭进. 北部湾地形、底质特征与渔场分布的关系[J]. 海洋湖沼通报, 2007(s1): 232-238. QIAO Y L, LIN Z J. The relationship between the main features of land form, the distribution of bottom sediment and fishery distribution[J]. Transactions of Oceanology and Limnology, 2007(s1): 232-238. |
| [2] |
CHEN Z Z, QIU Y S, XU S N. Changes in trophic flows and ecosystem properties of the Beibu Gulf ecosystem before and after the collapse of fish stocks[J]. Ocean & Coastal Management, 2011, 54(8): 601-611. |
| [3] |
蔡研聪, 徐姗楠, 陈作志, 等. 南海北部近海渔业资源群落结构及其多样性现状[J]. 南方水产科学, 2018, 14(2): 10-18. CAI Y C, XU S N, CHEN Z Z, et al. Current status of community structure and diversity of fishery resources in offshore northern South China Sea[J]. South China Fisheries Science, 2018, 14(2): 10-18. DOI:10.3969/j.issn.2095-0780.2018.02.002 |
| [4] |
傅昕龙. 北部湾西北部近海鱼类资源分布和群落结构特征及其影响因素[D]. 上海: 上海海洋大学, 2018. FU X L. Distribution of fish populations and structure of fish communities in the coastal waters of northwest Beibu Gulf and their influential factors[D]. Shanghai: Shanghai Ocean University, 2018. |
| [5] |
李渊, 张静, 张然, 等. 南沙群岛西南部海域和北部湾口海域底层游泳动物多样性[J]. 中国水产科学, 2016, 23(1): 177-187. LI Y, ZHANG J, ZHANG R, et al. Diversity of demersal nekton in the southwestern sea of the Nansha Islands and the mouth of Beibu Bay[J]. Journal of Fishery Sciences of China, 2016, 23(1): 177-187. |
| [6] |
王雪辉, 邱永松, 杜飞雁, 等. 北部湾秋季底层鱼类多样性和优势种数量的变动趋势[J]. 生态学报, 2012, 32(2): 333-342. WANG X H, QIU Y S, DU F Y, et al. Dynamics of demersal fish species diversity and biomass of dominant species in autumn in the Beibu Gulf, northwestern South China Sea[J]. Acta Ecologica Sinica, 2012, 32(2): 333-342. |
| [7] |
王雪辉, 邱永松, 杜飞雁, 等. 北部湾鱼类多样性及优势种的时空变化[J]. 中国水产科学, 2011, 18(2): 427-436. WANG X H, QIU Y S, DU F Y, et al. Spatio-temporal variability of fish diversity and dominant species in the Beibu Gulf[J]. Journal of Fishery Sciences of China, 2011, 18(2): 427-436. |
| [8] |
陈作志, 邱永松, 贾晓平, 等. 捕捞对北部湾海洋生态系统的影响[J]. 应用生态学报, 2008, 19(7): 1604-1610. CHEN Z Z, QIU Y S, JIA X P, et al. Effects of fishing on the marine ecosystem of Beibu Gulf[J]. Chinese Journal of Applied Ecology, 2008, 19(7): 1604-1610. |
| [9] |
乔延龙, 陈作志, 林昭进. 北部湾春、秋季渔业生物群落结构的变化[J]. 中国水产科学, 2008, 15(5): 816-821. QIAO Y L, CHEN Z Z, LIN Z J. Changes of community structure of fishery species during spring and autumn in BeibuGulf[J]. Journal of Fishery Sciences of China, 2008, 15(5): 816-821. DOI:10.3321/j.issn:1005-8737.2008.05.014 |
| [10] |
王雪辉, 杜飞雁, 邱永松. 南海北部主要经济鱼类体长与体重关系[J]. 台湾海峡, 2006, 25(2): 262-266. WANG X H, DU F Y, QIU Y S. Length-weight relationships of important commercial fishes in northern South China Sea[J]. Journal of Oceanography in Taiwan Strait, 2006, 25(2): 262-266. DOI:10.3969/j.issn.1000-8160.2006.02.017 |
| [11] |
孙典荣, 李渊, 林昭进, 等. 海南岛近岸海域鱼类群落结构研究[J]. 中国海洋大学学报, 2011, 41(4): 33-38. SUN D R, LI Y, LIN Z J, et al. Community structure of fish in the coastal waters of Hainan Island[J]. Periodical of Ocean University of China, 2011, 41(4): 33-38. |
| [12] |
何雄波, 陶雅晋, 侯刚, 等. 北部湾斑鳍白姑鱼群体结构特征与时空分布[J]. 广东海洋大学学报, 2015, 35(3): 35-42. HE X B, TAO Y J, HOU G, et al. Population structure and spatio-temporal distribution of Pennahia pawak in the Beibu Gulf, South China Sea[J]. Journal of Guangdong Ocean University, 2015, 35(3): 35-42. DOI:10.3969/j.issn.1673-9159.2015.03.006 |
| [13] |
颜云榕. 北部湾主要鱼类摄食生态及食物关系的研究[D]. 青岛: 中国科学院研究生院(海洋研究所), 2010. YAN Y R. Feeding ecology and food relations of the main fishes in the Beibu Gulf, South China Sea[D]. Qingdao: The Institute of Oceanology, Chinese Academy of Sciences, 2010. |
| [14] |
颜云榕, 侯刚, 卢伙胜, 等. 北部湾斑鳍白姑鱼的年龄与生长[J]. 中国水产科学, 2011, 18(1): 145-155. YAN Y R, HOU G, LU H S, et al. Age and growth of pawak croaker Pennahia pawak in Beibu Gulf[J]. Journal of Fishery Sciences of China, 2011, 18(1): 145-155. |
| [15] |
侯刚, 刘丹丹, 冯波, 等. 基于地标点几何形态测量法识别北部湾4种白姑鱼矢耳石形态[J]. 中国水产科学, 2013, 20(6): 1293-1302. HOU G, LIU D D, FENG B, et al. Using landmark-based geometric morphometrics analysis to identify sagittal otolith of four fish species[J]. Journal of Fishery Sciences of China, 2013, 20(6): 1293-1302. |
| [16] |
詹秉义. 渔业资源评估[M]. 北京: 中国农业出版社, 1995. ZHAN B Y. Fish stock assessment[M]. Beijing: China Agriculture Press, 1995. |
| [17] |
刘涛, 戴小杰, 高春霞. 印度洋大眼金枪鱼生长和死亡参数的估算[J]. 上海海洋大学学报, 2015, 24(1): 102-107. LIU T, DAI X J, GAO C X. Estimation of growth and mortality parameters of bigeye tuna (Thunnus obesus) in the Indian Ocean[J]. Journal of Shanghai Ocean University, 2015, 24(1): 102-107. |
| [18] |
叶婷, 王迎宾, 周丛羽. 鱼类体长频率数据结构对生长参数估算的影响分析[J]. 水产科学, 2014, 33(5): 277-282. YE T, WANG Y B, ZHOU C Y. Analysis of effects of fish length frequency data on estimates of growth parameters[J]. Fisheries Science, 2014, 33(5): 277-282. DOI:10.3969/j.issn.1003-1111.2014.05.003 |
| [19] |
RICKER W E. Linear regressions in fishery research[J]. Journal of the Fisheries Research Board of Canada, 1973, 30(3): 409-434. DOI:10.1139/f73-072 |
| [20] |
FROESE R. Cube law, condition factor and weight-length relationships: history, meta-analysis and recommendations[J]. Journal of Applied Ichthyology, 2006, 22(4): 241-253. DOI:10.1111/j.1439-0426.2006.00805.x |
| [21] |
PITCHER T J, HART P J B. Fisheries ecology[M]. London: Croom Helm, 1982.
|
| [22] |
PAULY D, DAVID N. ELEFAN I, a BASIC Program for the objective extraction of growth parameters from length-frequency data[J]. Meeresforsch, 1981, 28(4): 205-211. |
| [23] |
MUNRO J L, PAULY D. A simple method forcomparing the growth of fishes and invertebrates[J]. Fishbyte, 1983, 1(1): 5-6. |
| [24] |
PAULY D. Some simple methods for the assessment of tropical fish stocks[J]. FAO Fisheries Technical Paper, 1983, 234: 52. |
| [25] |
PAULY D. On the interrelationships between natural mortality, growth parameters, and mean environmental temperature in 175 fish stocks[J]. ICES Journal of Marine Science, 1980, 39(2): 175-192. DOI:10.1093/icesjms/39.2.175 |
| [26] |
何宝全, 李辉权. 珠江河口棘头梅童鱼的资源评估[J]. 水产学报, 1988, 12(2): 124-134. HE B Q, LI H Q. Stock Assessment of Collichthys lucidus in pearl river esturary[J]. Journal of Fisheries of China, 1988, 12(2): 124-134. |
| [27] |
王雪辉, 邱永松, 杜飞雁, 等. 基于长度贝叶斯生物量法估算北部湾二长棘鲷种群参数[J]. 水产学报, 2020, 44(10): 1654-1662. WANG X H, QIU Y S, DU F Y, et al. Using length-based Bayesian biomass method to estimate Parargyrops edita population parameters in the Beibu Gulf, South China Sea[J]. Journal of Fisheries of China, 2020, 44(10): 1654-1662. |
| [28] |
耿平. 北部湾典型鱼类种群生长、死亡及开发状态的年际变化研究[D]. 上海: 上海海洋大学, 2019. GENG P. A study of inter-annual changes in growth, mortality and exploitation rate of representative fish stocks in BeibuGulf[D]. Shanghai: Shanghai Ocean University, 2019. |
| [29] |
彭露. 南海北部近海金线鱼的年龄、生长及数量分布[D]. 上海: 上海海洋大学, 2019. PENG L. Age, growth and quantity distribution of Nemipterusvirgatus in the northern coast of the South China Sea[D]. Shanghai: Shanghai Ocean University, 2019. |
| [30] |
张魁, 陈作志, 王跃中, 等. 北部湾短尾大眼鲷群体结构及生长、死亡和性成熟参数估计[J]. 热带海洋学报, 2016, 35(5): 20-28. ZHANG K, CHEN Z Z, WANG Y Z, et al. Population structure of Priacanthus macracanthus in the Beibu Gulf, and parameters for its growth, mortality and maturity[J]. Journal of Tropical Oceanography, 2016, 35(5): 20-28. |
| [31] |
黄真理, 常剑波. 鱼类体长与体重关系中的分形特征[J]. 水生生物学报, 1999, 23(4): 330-336. HUANG Z L, CHANG J B. Fractal characteristics of length-weight relation ship in fish[J]. Acta Hydrobiologica Sinica, 1999, 23(4): 330-336. DOI:10.3321/j.issn:1000-3207.1999.04.006 |
| [32] |
徐海龙, 谷德贤, 乔秀亭, 等. 渤海湾主要渔业资源长度与体质量关系分析[J]. 南方水产科学, 2014, 10(1): 57-63. XU H L, GU D X, QIAO X T, et al. Analysis of length-weight relationship for major fishing species from Bohai Bay[J]. South China Fisheries Science, 2014, 10(1): 57-63. DOI:10.3969/j.issn.2095-0780.2014.01.009 |
| [33] |
朱立新, 侯刚, 梁振林. 基于贝叶斯方法的黄海北部鳀鱼体长与体重关系式参数估计[J]. 中国水产科学, 2015, 22(4): 757-769. ZHU L X, HOU G, LIANG Z L. Parameter estimation of the weight-length relationship of Japanese anchovy in the north Yellow Sea using Bayesian methods[J]. Journal of Fishery Sciences of China, 2015, 22(4): 757-769. |
| [34] |
STERGIOU K I, MOUTOPOULOS D K. A Review of length-weight relationships of fishes from greek marine waters[J]. Naga, 2001, 24(1/2): 23-39. |
| [35] |
李策, 李新辉, 李跃飞, 等. 基于单位补充量模型的西江赤眼鳟种群资源利用现状评价[J]. 中国水产科学, 2019, 26(1): 151-160. LI C, LI X H, LI Y H, et al. Exploitation status of Squaliobarbus curriculus in the Xijiang River based on the analysis of the yield per recruit and spawning biomass per recruit models[J]. Journal of Fishery Sciences of China, 2019, 26(1): 151-160. |
| [36] |
叶金清, 徐兆礼, 陈佳杰, 等. 基于生长和死亡参数变化的官井洋大黄鱼资源现状分析[J]. 水产学报, 2012, 36(2): 238-246. YE J Q, XU Z L, CHEN J J, et al. Resources status analysis of large yellow croaker in Guanjinyang using von Bertalanffy growth equation and fishing mortality parameters[J]. Journal of Fisheries of China, 2012, 36(2): 238-246. |
| [37] |
GULLAND J A. Fish stock assessment: a manual of basic methods[M]. New York: FAO/Wiley, 1983.
|
| [38] |
陈作志, 邱永松, 黄梓荣. 南海北部白姑鱼生长和死亡参数的估算[J]. 应用生态学报, 2005, 16(4): 712-716. CHEN Z Z, QIU Y S, HUANG Z R. Estimation of growth and mortality parameters of Argyrosomus argentatus in northern South China Sea[J]. Chinese Journal of Applied Ecology, 2005, 16(4): 712-716. DOI:10.3321/j.issn:1001-9332.2005.04.025 |
| [39] |
官文江, 高峰, 雷林, 等. 渔业资源评估中的回顾性问题[J]. 上海海洋大学学报, 2012, 21(5): 841-847. GUAN W J, GAO F, LEI L, et al. Retrospective problem in fishery stock assessment[J]. Journal of Shanghai Ocean University, 2012, 21(5): 841-847. |
| [40] |
张波, 金显仕, 戴芳群. 长江口两种重要石首鱼类的摄食习性[J]. 动物学报, 2008, 54(2): 209-217. ZHANG B, JIN X S, DAI F Q. Feeding habits of the two sciaenid fishes near the Changjiang estuary[J]. Acta Zoologica Sinica, 2008, 54(2): 209-217. |
| [41] |
何雄波. 中国近海带鱼科(Trichiuridae)常见种空间分布、种群结构与营养生态研究[D]. 厦门: 集美大学, 2019. HE X B. Spatial distribution, population structure and trophic ecology of common Trichiuridae species in the coastal waters of China[D]. Xiamen: Jimei University, 2019. |
| [42] |
颜云榕, 王田田, 侯刚, 等. 北部湾多齿蛇鲻摄食习性及随生长发育的变化[J]. 水产学报, 2010, 34(7): 1089-1098. YAN Y R, WANG T T, HOU G, et al. Feeding habits and monthly and ontogenetic diet shifts of the greater lizardfish, Saurida tumbil in the Beibu Gulf of the South China Sea[J]. Journal of Fisheries of China, 2010, 34(7): 1089-1098. |
| [43] |
颜云榕, 杨厚超, 卢伙胜, 等. 北部湾宝刀鱼的摄食生态[J]. 生态学报, 2011, 31(3): 654-665. YAN Y R, YANG H C, LU H S, et al. Feeding ecology of dorab wolf-herring, Chirocentrus dorab from the Beibu Gulf[J]. Acta Ecologica Sinica, 2011, 31(3): 654-665. |
| [44] |
蒋日进, 金海卫, 周永东, 等. 东海竹䇲鱼的食性[J]. 应用生态学报, 2013, 24(7): 2015-2024. JIANG R J, JIN H W, ZHOU Y D, et al. Feeding habits of Trachurus japonicus in the East China Sea[J]. Chinese Journal of Applied Ecology, 2013, 24(7): 2015-2024. |
| [45] |
李忠炉, 张文旋, 何雄波, 等. 南海北部湾秋季蓝圆鲹与竹䇲鱼的摄食生态及食物竞争[J]. 广东海洋大学学报, 2019, 39(3): 79-86. LI Z L, ZHANG W X, HE X B, et al. Feeding ecology and feeding competition between Decapterus maruadsi and Trachurus japonicus in autumn in the Beibu Gulf, South China Sea[J]. Journal of Guangdong Ocean University, 2019, 39(3): 79-86. |
| [46] |
牛翠娟, 娄安如, 孙儒泳, 等. 基础生态学[M]. 2版. 北京: 高等教育出版社, 2007. NIU C J, LOU A R, SUN R Y. Fundamentals of ecology[M]. 2nd ed. Beijing: Higher Education Press, 2007. |
 ,
QIU Kangwen1,
ZHOU Xiao1,
ZHAO Chunxu2,3,
DENG Yujian1,
HE Xiongbo1,
YAN Yunrong1,2,4,5
,
QIU Kangwen1,
ZHOU Xiao1,
ZHAO Chunxu2,3,
DENG Yujian1,
HE Xiongbo1,
YAN Yunrong1,2,4,5
2. Marine Resources Big Data Center of South China Sea, Southern Marine Science and Engineering Guangdong Laboratory, Zhanjiang 524013, Guangdong, China;
3. College of Fisheries, Jimei University, Xiamen 361021, Fujian, China;
4. Guangdong Provincial Engineering and Technology Research Center of Far Sea Fisheries Management and Fishing of South China Sea, Zhanjiang 524088, Guangdong, China;
5. Center of Marine Fisheries Information Technology, Shenzhen Institute of Guangdong Ocean University, Shenzhen 518120, Guangdong, China