 2021,
Vol. 30
2021,
Vol. 30



2. 上海海洋大学 海洋动物系统分类与进化上海高校重点实验室, 上海 201306;
3. 上海海洋大学 水产动物遗传育种中心上海市协同创新中心, 上海 201306;
4. 上海城投原水有限公司, 上海 200125
青草沙水库位于上海市崇明区长兴岛西部,是目前世界上最大的河口江心避咸蓄淡型水库,也是上海市最大的饮用水水源地[1]。青草沙水库2007年6月正式开工建设,2010年12月投入运行[2]。水库在运行期间,依据长江口潮汐和盐度的变化趋势,采用“避污蓄清”“避咸蓄淡”的原则定期开闸纳水和排水,鱼类随之进出水库,水库与长江口之间存在鱼类的交换。自2018年起,在取水口外围建起了细密隔栅,水库与长江之间仅鱼类的卵、小型的仔稚鱼可以通过隔栅进出水库。
青草沙水库在运行之初,鱼类本底来自长江口,自2013年起,管理部门根据水库水生生物的变动,适量投放鲢(Hypophthalmichthys molitrix)、鳙(Hypophthalmichthys nobilis)等鱼类来调节水质。青草沙水库自建成以来,胡忠军等[3]对鱼类群落结构进行了初步调查,王绍祥等[4]2011年对青草沙水库鱼类群落组成、多样性及时空分布特征进行了分析,李丽等[5]在2011年采用碳氮稳定同位素的方法分析了优势种刀鲚食性组成和转变。2015至2019年对鱼类进行了季节性的调查和鱼类群落结构的年际变化分析,探讨青草沙水库运行以来鱼类群落结构的变动趋势和演替方向,为研究青草沙水库鱼类演替规律和管理措施提供科学依据。
1 材料与方法 1.1 调查区域和渔具2015至2019年按春、夏、秋、冬四季分别对青草沙水库进行了为期5年的鱼类调查。根据水库渔业管理队、双声呐鱼探仪探测的鱼类分布区及相关文献信息,在全库布设7个有代表性3层刺网网点和围库1周若干个地笼网网点(图 1)。3层刺网长度均为长100 m、高7 m,网目孔径有2种规格:外层直径15 cm、中间层直径8 cm;外层直径10 cm、中间层直径6 cm。25 m有结地笼网(76节60门、网框宽40 cm×高30 cm)、50 m有结地笼网(网框宽100 cm×高100 cm,网眼对角线8 mm)等不同规格,地笼网主要放置在岸边和湖心岛周围等浅水区。刺网前一天上午布设、次日清晨收取;地笼网放置7 d后收取;所有渔获物带回实验室进行分析[6]。

|
●刺网采样点S1-S7;▲地笼网采样点。 ●sampling sites s1-s7 of gillnet; ▲sampling sites of cage. 图 1 青草沙水库调查站点示意图 Fig. 1 Sampling sites of Qingcaosha reservoir |
用量鱼板(精度1 cm)、游标卡尺(精度0.01 cm),配合两脚规对样本进行全长、体长等数据的测量;用天平称量体质量(精度0.1 g),小个体用分析天平称量(精度0.01 g),写上编号、样本批次、采样日期与采样渔具。根据《中国动物志·硬骨鱼纲》(鲤形目·上卷)、《中国动物志·硬骨鱼纲》(鲤形目·中卷)[7]、《中国动物志·硬骨鱼纲·鲈形目(四)》、《上海鱼类志》[8]、《Fishes of the World》[9]等相关分类学参考书籍,对样本进行种类鉴定。
1.2 数据分析 1.2.1 优势种基于在青草沙水库的鱼类样本采样情况及所获取的数据特征,计算Pinkas相对重要性指数(Index of Relative Importance,IRI),分析鱼类群落的优势种,IRI的计算式[10]为
 (1)
(1)
式中:IRI为相对重要性指数;N为某一种鱼类样本个体数占渔获物总个体数的百分比,%;W为某一种鱼类样本质量占渔获物总质量的百分比,%;F某一种鱼类样本出现站点数占调查站点总数的百分比,%。
1.2.2 多样性指数物种多样性指数包括物种丰富度和均匀度,是用来描述一个群落多样性的统计量,直观地表现出生物群落结构多样性的高低[11]。可用来衡量青草沙水库鱼类多样性高低和变化趋势,主要有以下3个函数:
(1) Shannon-Wiener多样性指数的计算公式[12]为
 (2)
(2)
式中: S为采集样本种类总数; pi为第i种鱼类个体数与渔获物总个体数的比值。
(2) Pielou均匀度指数, 用于计算群落中不同鱼类的均匀性,计算公式[13]为
 (3)
(3)
式中:H′为Shannon-Wiener多样性指数;Hmax为lnS,S为样本中鱼类种类总数。
(3) Margalef丰富度指数衡量一个群落中存在物种数目,并进一步分析该群落物种丰度程度,计算公式[14]为
 (4)
(4)
式中:S为所采集样本中种类总数;N为样本中鱼类总个体数。
1.2.3 群落稳定性分析(ABC曲线)鱼类群落结构稳定性分析使用Warwick的丰度-生物量比较ABC曲线(abundance-biomass comparison curves)[15]。鱼类数量优势度曲线和生物量优势度曲线在一个坐标系中同时存在,通过比较两条曲线在坐标系中的位置来分析群落受干扰程度。用W统计值(W-statistic) 作为ABC曲线方法的一个统计量,公式为
 (5)
(5)
式中:Bi为第i种鱼类生物量累计百分比;Ai为第i种鱼类数量累计百分比;S为样本中鱼类种类总数。
1.2.4 群落相似性分析等级聚类(cluster)是分析鱼类群落结构特征的方法,cluster聚类基于Bray-Curtis相似性系数来建立样本间的相似性矩阵[16]。
 (6)
(6)
式中:S表示鉴定分析的物种数; xij和xim分别代表群落中第i种鱼类在第j个和第m个站点的丰度,物种丰度数据均先进行lg(x+1)转换。
将聚类分析得到的每个分类组别进行ANOSIM(analysis of similarities)差异显著性检验,用R值来表示差异显著性,R值范围在0到1之间,越接近数值1表示组内样本间相似性高于组间样本相似性,R值接近0,表示组内与组间相似性显著。SIMPER(similarity of percentage analysis)相似性百分比分析,进一步检验不同样本的组内和组间的相似性,贡献率≥2%的种类定义为主要特征种[17]。
1.3 数据处理采用PRIMER 5.0软件[18]、R语言3.6.2、SPSS软件[19]对数据进行分析。
2 结果 2.1 种类组成及其年际变化2015—2019年共获得5 158尾鱼类样本,鉴定为51种,隶属于为9目16科41属,分别为:鲤形目34种,占种类总数的66.67%;鲈形目8种,占比15.69%;鲇形目3种,占比5.89%;其余为鲟形目、鲱形目、鳗鲡目、颌针鱼目、鲻形目、鲽形目等均为1种。
2015—2019各年间,调查到的鱼类种类、分类类群年份间存在差异。2015年种类数最少,仅调查到26种鱼类,隶属于7目8科26属;2018年种类数最多,31种,隶属于7目12科28属。2019和2015年调查结果相比,调查渔获物由7目下降到4目,没有调查到鲽形目、鳗鲡目、鲻形目、鲽形目(表 1)。

|
表 1 2015—2019年各年间青草沙水库鱼类分类概况 Tab.1 Faunal composition of fishes in Qingcaosha Reservoir from 2015 to 2019 |
青草沙地处长江口,水域受到淡水径流和海水入侵的交互作用,生境复杂,参照ELLIOTT等[20]对河口地区鱼类分类,青草沙水库鱼类可分为4种类型:河口定居鱼类(ES),如半滑舌鳎(Cynoglossus semilaevis)等,生物量占总渔获量的0.8%;降海洄游鱼类(CA),如花鲈(Lateolabrax maculates)、日本鳗鲡(Anguilla japenica)等,丰度为3.1%;溯河洄游鱼类(AW),如间下

|
图 2 2015—2019年青草沙水库鱼类生态类型 Fig. 2 Ecological types of fish in Qingcaosha Reservoir from 2015 to 2019 |
根据鱼类主要食物类型,可将青草沙鱼类分为4种食性类型[21]:肉食性鱼类、杂食性鱼类、滤食性鱼类、植食性鱼类。其中,滤食性鱼类鲢和鳙,丰度占总量11%,生物量占总渔获量71%(图 3)。

|
图 3 2015—2019年青草沙水库鱼类食性类型 Fig. 3 Feeding habits of fish in Qingcaosha Reservoir from 2015 to 2019 |
2015—2019年间,青草沙水库降海洄游型鱼类,如中华鲟等,丰度占比由2015年的7.4%下降到2019年的0.8%(图 4a);溯河洄游型鱼类,如间下

|
图 4 2015—2019年青草沙水库渔获物中不同生态类群的年际变化 Fig. 4 interannual changes of different ecological groups in Qingcaosha Reservoir from 2015 to 2019 |
2015—2019年,青草沙水库优势度大于100的鱼类有20种(表 3),鲢优势度最高(IRI为5 187),其次是鲫(IRI为2 905),鳙优势度第三(IRI为2 399)。有些小型鱼类优势度也很高,如优势度排名第4的大鳍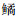

|
表 2 2015—2019年青草沙水库历年优势度(IRI)前5的鱼类 Tab.2 species of the top five dominant specie in Qincaosha Reservoir from 2015 to 2019 |

|
表 3 A、B组内相似性和物种贡献率 Tab.3 Similarity and species contribution rate in groups A and B |
2015年到2019年调查期间,鲢、鳙、鲫等3种鱼类在所有调查年份中均为优势种;棒花鱼作为优势种出现在3个年份(2015、2016、2018年),大鳍
青草沙水库2015—2019年,Margalef丰富度指数、Shannon-Wiener指数H′和Pielou均匀度指数这3个指数变动范围均比较小,分别为这3.76~4.30、2.55~2.70和0.76~0.83。Margalef丰富度指数2018年最高,为4.30;2015年最低,为3.76。Pielou均匀度指数2015年为0.83,2019年下降至0.76。Shannon指数2015年为2.70,2019年下降至2.55。均匀度指数和香农多样性指数,均有所下降(图 5)。将青草沙水库每年的3个多样指数进行相关性分析,多样性指数H和丰富度指数D相关性显著,皮尔逊检验相关系数为0.984(P=0.001,P<0.01),多样性指数H和均匀度指数J相关性也显著,皮尔逊检验相关系数为0.997(P=0.001,P<0.01)。

|
图 5 青草沙水库鱼类群落多样性指数年际变化 Fig. 5 Interannual variation of diversity indices of the fish community in Qingcaosha Reservoir |
根据原始数据建立相似性矩阵,对群落进行Cluster聚类分析来分析青草沙水库不同年份鱼类群落的结构特征[22],结果显示相似性水平为53%,2015、2016年聚为第一支(简称“A支”);2017、2018、2019聚为第二支(简称“B支),B支中又以2017、2018年相似性最高(图 6)。

|
图 6 青草沙水库2015—2019年鱼类丰度的Bray-Curtis相似性聚类图 Fig. 6 Bray-Curtis similarity cluster analysis of fish abundance from 2015 to 2019 in Qingcaosha reservoir |
ANOSIM分析检验聚类分析得到的A、B两支显著性,结果显示R=0.25,P=0.4(P>0.05),差异不显著。通过SIMPER相似性百分比计算出对组内相似性和组间相异性贡献较大的鱼类,根据贡献率的高低确定造成不同年份差异的关键物种。
SIMPER分析结果显示,A支平均相似性为66.75%,B支平均相似性为62.68%,A组鱼类对组内相似性贡献率高低依次为棒花鱼、大鳍
2015—2019年调查期间,青草沙水库每年的生物量优势度曲线始终在丰度优势度曲线上方,丰度比生物量具有更高的多样性[23],表明青草沙水库的群落结构受干扰程度很低。ABC曲线用W值来量化,2015年到2019年的(W)值依次为0.311、0.205、0.233、0.208、0.246,青草沙水库的W值无单一规律,鱼类群落结构处于一种波动调整状态,但生物量曲线始终处于丰度曲线的上方,表明群落结构比较稳定(图 7)。

|
图 7 2015—2019年青草沙水库ABC曲线 Fig. 7 ABC curves of 2015—2019 in Qingcaosha Reservoir |
研究期间青草沙水库调查到51种鱼类,淡水鲤形目种类占比高达66.67%,是青草沙水库鱼类的主体。胡忠军等[24]2010年在水库正式供水前调查到鱼类18种,淡水鲤形目占比72.2%。淡水鱼类的比例比正式供水前增加,表明水库淡水区系特征变得更明显。邱永松[25]指出生境物种与其生境环境显著相关,尤其受水体盐度和温度影响较大,与水体深度也有一定关联,这与青草沙的情况也是相符的,正式供水前后青草沙水库盐度发生变化、水深增加,鱼类组成发生变化。
优势度直观地反应出群落中的优势种。调查期间,优势度前3的物种依次是鲢(5 187)、鲫(2 905)、鳙(2 399),且鲢、鳙、鲫等3种鱼类在每年调查中均为优势种。胡忠军等[24]调查的优势种为贝氏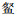
王雪辉等[26]指出,鱼类群落结构主要受到生境变化的影响,从生态类群来看,青草沙水库降海洄游、溯河洄游、河口型鱼类丰度占比减少,淡水定居型数量增多,整个青草沙水库偏向淡水区系鱼类特性。
刘媛媛[27]2014年对长江口繁殖盛期的仔稚鱼进行种类组成调查,结果表明鲈形目鱼类丰度占比最多,其次是鲤形目,半咸水鱼类比例最高。张涛等[28]对长江口最大的滩涂湿地崇明东滩鱼类群落组成进行研究,结果表明鲈形目鱼类最多,群落优势种为中国花鲈、窄体舌鳎(Crnogrossus gracilis)、刀鲚等河口型和洄游型鱼类。刘凯等[29]对崇明岛北滩进行鱼类群落研究表明北滩的江海洄游型和河口型鱼类占优势,优势种为凤鲚(Coilia mystus)、梅童鱼(Collichthys lucidus)、小黄鱼(Pseudoscinaena polyactis)等, 且北滩的鱼类趋向于小型化,生物多样性有下降趋势[30]。龚小玲等[31]对长江口九段沙湿地进行鱼类调查研究,调查到的42种鱼类中,鲈形目鱼类最多,河口型鱼类占绝对优势,优势种为斑尾刺虾虎鱼、四指马鲅(Eleutheronema tetradactylum)等。以上4个长江口邻近水域的鱼类物种组成中,鲈形目鱼类均占优势,从生态类群上看,河口型和近海型鱼类比例较高,而从长江口取水的青草沙水库,鱼类生态类群以淡水鲤科鱼类为主,与通畅的长江口已形成差异。
韩婵等[32]2012年对上海淡水湖泊淀山湖进行鱼类群落结构调查表明,淀山湖鲤科鱼类占优势,且小型鱼类数量占优势,淀山湖增殖放流的鲢、鳙优势度高,这与青草沙水库的调查结果相似。青草沙水库鱼类结构组成已由过去的长江口鱼类群落结构特征向长江下游淡水型水库、湖泊鱼类结构特征转化,人为干预对青草沙水库鱼类结构的影响也进一步形成。
3.3 多样性指数和群落稳定性在2015至2019年调查期间,青草沙水库的鱼类群落结构Margalef指数波动无明显规律,Shannon-Wiener指数,均有下降趋势,但变动范围很小。Pielou均匀度指数变化不显著,这个结果与朱鑫华等[33]和金斌松[34]的研究结果一致: 当外界环境发生变化的时候,鱼类群落的均匀度指数变化很小,即均匀度对环境的变化不敏感。李圣法等[35]研究认为鱼类物种多样性指数的变化和该群落优势鱼类关联较大,青草沙水库每年优势种都有一定的变化,人为干扰活动强度也与物种多样性显著相关。2019年多样性指数较2015年相比略微下降,可能是与青草沙水库淡水鱼类增多,也可能是与调查等很多因素相关。鱼类群落生态多样性指数与很多因素有关,除了环境因素外,在采样时,网具网眼大小,深度等也会影响多样性结果。
ABC曲线在同一坐标系中反映了鱼类群落中生物量和丰度的变化趋势[36]。实质上和传统进化中r选择和k选择理论背景是一致的:k选择是以生长慢、性成熟晚的大个体种类为主[37];当群落受到干扰时,以r选择的物种(生长快、个体小的种类) 为主。2015—2019年调查期间,青草沙水库的生物量优势度曲线每年都处于丰度优势度曲线上方,W值在0.205~0.311波动,调查期间5年综合W值为0.246,W值均为正值,表明青草沙水库以生长慢、性成熟晚的大个体鱼类为主。但青草沙水库的两条曲线有交叉的趋势,W值在逐年减小,水库受干扰的程度在逐年增加,青草沙水库的鱼类群落结构有向生长速度快、性成熟周期短的小型鱼类演替的趋势。
放养鲢、鳙,避咸取淡、取水口设置隔栏等人工干预对青草沙水库鱼类结构的影响慢慢形成,青草沙水库鱼类偏向于淡水区系,洄游鱼类减少,人为放养鲢、鳙是其优势度高的主要原因,鲫在库内自繁使其优势度逐渐升高。鱼类多样性指数平稳无异常波动,现阶段青草沙鱼类群落结构稳定,但有向小型鱼类演替的趋势。
| [1] |
顾金山, 陆晓如, 顾玉亮. 上海青草沙水源地原水工程规划[J]. 给水排水, 2009, 35(1): 50-54. GU J S, LU X R, GU Y L. Layout of Shanghai Qingcaosha drinking water source engineering[J]. Water & Wastewater Engineering, 2009, 35(1): 50-54. DOI:10.3969/j.issn.1002-8471.2009.01.015 |
| [2] |
宋少红. 长江口青草沙水源地原水工程可行性研究[D]. 上海: 上海交通大学, 2008: 53. SONG S H, Feasibility study on raw water project of Qingcaosha water source in Changjiang estuary[D]. Shanghai: Shanghai Jiao Tong University, 2008: 53. |
| [3] |
胡忠军, 史先鹤, 吴昊, 等. 上海青草沙水库食物网结构特征分析[J]. 水生态学杂志, 2019, 40(2): 47-54. HU Z J, SHI X H, WU H, et al. Food web structure in Qingcaosha reservoir of Shanghai, China[J]. Journal of Hydroecology, 2019, 40(2): 47-54. |
| [4] |
王绍祥, 高春霞, 田思泉, 等. 青草沙水库中上层鱼类群落组成及多样性分析[J]. 上海海洋大学学报, 2014, 23(4): 594-601. WANG S X, GAO C X, TIAN S Q, et al. Community composition and diversity of pelagic fishes in Qingcaosha Reservoir[J]. Journal of Shanghai Ocean University, 2014, 23(4): 594-601. |
| [5] |
李丽, 杜蓉, 刘其根, 等. 长江口青草沙水库刀鲚食性转变[J]. 中国水产科学, 2019, 26(4): 765-773. LI L, DU R, LIU Q G, et al. The ontogenetic dietary shift of Japanese grenadier anchovy (Coilia nasus) in the Qingcaosha Reservoir near the Yangtze River Estuary[J]. Journal of Fishery Sciences of China, 2019, 26(4): 765-773. |
| [6] |
沃佳, 徐宾铎, 薛莹, 等. 胶州湾鱼类群落优势种组成的时空变化[J]. 中国水产科学, 2017, 24(5): 1091-1098. WO J, XU B Z, XUE Y, et al. Temporo-spatial heterogeneity of dominant fish species in the Jiaozhou Bay community[J]. Journal of Fishery Sciences of China, 2017, 24(5): 1091-1098. |
| [7] |
陈宜瑜. 中国动物志硬骨鱼纲鲤形目(中卷)[M]. 北京: 科学出版社, 1998. CHEN Y Y. China fauna osteichthyes cypriniformes (in volume)[M]. Beijing: Science Press, 1998. |
| [8] |
温家华, 徐征和, 武玮, 等. 大汶河流域水生态系统健康评价研究[J]. 南水北调与水利科技, 2018, 16(3): 118-124. WEN J H, XU Z H, WU W, et al. Study on water ecosystem evaluation in Dawen River Basin[J]. South-to-North Water Transfers and Water Science & Technology, 2018, 16(3): 118-124. |
| [9] |
NELSON J S. Fishes of the world[M]. 4th ed. Hoboken: John Wiley & Sons, 2006.
|
| [10] |
PINKAS L, OLIPHANT M S, IVERSON I L K. Food habits of albacore, bluefin tuna, and bonito in California waters[J]. Fish Bulletin, 1971, 152: 1-105. |
| [11] |
杜建国, 陈彬, 卢振彬, 等. 泉州湾海域鱼类多样性及营养级变化[J]. 生物多样性, 2010, 18(4): 420-427. DU J G, CHEN B, LU Z B, et al. Changes of fish diversity and trophic levels in Quanzhou Bay[J]. Biodiversity Science, 2010, 18(4): 420-427. |
| [12] |
MORRIS M, WOODS L M, RACHE B. A novel ecological methodology for constructing ethnic-majority life tables in the absence of individual ethnicity information[J]. Journal of Epidemiology & Community Health, 2015, 69(4): 361-367. |
| [13] |
孙军, 刘东艳. 多样性指数在海洋浮游植物研究中的应用[J]. 海洋学报, 2004, 26(1): 62-75. SUN J, LIU D Y. The application of diversity indices in marine phytoplankton studies[J]. Acta Oceanologica Sinica, 2004, 26(1): 62-75. DOI:10.3321/j.issn:0253-4193.2004.01.007 |
| [14] |
MARGALEF R. On certain unifying principles in ecology[J]. The American Naturalist, 97(897): 357-374.
|
| [15] |
WARWICK R M. A new method for detecting pollution effects on marine macrobenthic communities[J]. Marine Biology, 1986, 92(4): 557-562. DOI:10.1007/BF00392515 |
| [16] |
杨刚. 长江口鱼类群落结构及其与重要环境因子的相关性[D]. 上海: 上海海洋大学, 2012: 67. YANG G. Fish community structure and correlation to major environmental factors in Yangtze River estuary[D]. Shanghai: Shanghai Ocean University, 2012: 67. |
| [17] |
刘飞, 娄必云, 刘焕章, 等. 赤水河赤水市江段鱼类群落结构季节变化特征[J]. 淡水渔业, 2015, 45(1): 35-41. LIU F, LOU B Y, LIU H Z, et al. Seasonal variations of fish assemblage in Chishui Reach of the Chishui River[J]. Freshwater Fisheries, 2015, 45(1): 35-41. DOI:10.3969/j.issn.1000-6907.2015.01.007 |
| [18] |
沈新强, 史赟荣, 晁敏, 等. 夏、秋季长江口鱼类群落结构[J]. 水产学报, 2011, 35(5): 700-710. SHEN X Q, SHI Y R, CHAO M, et al. Fish community structure of the Yangtze River Estuary in summer and autumn[J]. Journal of Fisheries of China, 2011, 35(5): 700-710. |
| [19] |
MCCORMICK K, SALCEDO J. IBM SPSS data preparation[M]//MCCORMICK K, SALCEDO J. SPSSreg Statistics for Data Analysis and Visualization. Indianapolis, Indiana: John Wiley & Sons, 2017: 303-324.
|
| [20] |
ELLIOTT M, WHITFIELD A K, POTTER I C, et al. The guild approach to categorizing estuarine fish assemblages: a global review[J]. Fish and Fisheries, 2007, 8(3): 241-268. DOI:10.1111/j.1467-2679.2007.00253.x |
| [21] |
窦硕增. 鱼类摄食生态研究的理论及方法[J]. 海洋与湖沼, 1996, 27(5): 556-561. DOU S Z. Feeding ecology of fish-a critical review on theories, methods and their application[J]. Oceanologia et Limnologia Sinica, 1996, 27(5): 556-561. DOI:10.3321/j.issn:0029-814X.1996.05.016 |
| [22] |
张鹏弛. 南黄海冷水团大型底栖动物群落结构长期变化[D]. 临汾: 山西师范大学, 2017: 66. ZHANG P C. Long-term changes of the macrobenthic community structure in the cold water mass of the southern Yellow Sea[D]. Linfen: Shanxi Normal University, 2017: 66. |
| [23] |
董鹏, 张海波, 叶仙森, 等. 象山港中部海域大型底栖动物群落组成及多样性特征[J]. 上海海洋大学学报, 2015, 24(3): 430-440. DONG P, ZHANG H B, YE X S, et al. Community structure and biodiversity characteristics of macrobenthos in the middle of Xiangshan Bay[J]. Journal of Shanghai Ocean University, 2015, 24(3): 430-440. |
| [24] |
胡忠军, 吴昊, 陈立侨, 等. 长江口青草沙水库正式供水前的鱼类群落结构特征[J]. 水产学报, 2012, 36(8): 1234-1241. HU Z J, WU H, CHEN L Q, et al. Structure of fish assemblages before drinking-water supplies in Qingcaosha Reservoir of Yangtze River Estuary[J]. Journal of Fisheries of China, 2012, 36(8): 1234-1241. |
| [25] |
邱永松. 南海北部大陆架鱼类群落的区域性变化[J]. 水产学报, 1988, 12(4): 303-313. QIU Y S. The regioal changes of fish community on the northern continental shelf of South China Sea[J]. Journal of Fisheries of China, 1988, 12(4): 303-313. |
| [26] |
王雪辉, 邱永松, 杜飞雁, 等. 北部湾鱼类群落格局及其与环境因子的关系[J]. 水产学报, 2010, 34(10): 1579-1586. WANG X H, QIU Y S, DU F Y, et al. Fish community pattern and its relation to environmental factors in the Beibu Gulf[J]. Journal of Fisheries of China, 2010, 34(10): 1579-1586. |
| [27] |
刘媛媛. 长江口鱼类繁殖盛期仔稚鱼种类组成及优势种分布研究[D]. 上海: 上海海洋大学, 2016. LIU Y Y. The categories composition of fish larvae and juveniles and distribution of dominant species in reproductive periodin in the Yangtze estuary[D]. Shanghai: Shanghai Ocean University, 2016. |
| [28] |
张涛, 庄平, 刘健, 等. 长江口崇明东滩鱼类群落组成和生物多样性[J]. 生态学杂志, 2009, 28(10): 2056-2062. ZHANG T, ZHUANG P, LIU J, et al. Species composition and biodiversity of fish community in Chongming Dongtan of Yangtze River estuary[J]. Chinese Journal of Ecology, 2009, 28(10): 2056-2062. |
| [29] |
刘凯, 徐东坡, 张敏莹, 等. 崇明北滩鱼类群落生物多样性初探[J]. 长江流域资源与环境, 2005, 14(4): 418-421. LIU K, XU D P, ZHANG M Y, et al. Preliminary studies on biodiversity of fish community on north beach of Chongming island[J]. Resources and Environment in the Yangtze Basin, 2005, 14(4): 418-421. DOI:10.3969/j.issn.1004-8227.2005.04.005 |
| [30] |
张衡, 何文珊, 童春富, 等. 崇西湿地冬季潮滩鱼类种类组成及多样性分析[J]. 长江流域资源与环境, 2007, 16(3): 308-313. ZHANG H, HE W S, TONG C F, et al. Species composition and biodiversity of the intertidal fishes in Chongxi wetland in winter in Changjiang estuary[J]. Resources and Environment in the Yangtze Basin, 2007, 16(3): 308-313. DOI:10.3969/j.issn.1004-8227.2007.03.009 |
| [31] |
龚小玲, 张晓懿, 朱敏, 等. 长江口九段沙湿地潮沟鱼类组成及其多样性[J]. 上海海洋大学学报, 2011, 20(4): 517-524. GONG X L, ZHANG X Y, ZHU M, et al. Fish species composition and diversity in intertidal creek of Jiuduansha wetland in the Yangtze River estuary[J]. Journal of Shanghai Ocean University, 2011, 20(4): 517-524. |
| [32] |
韩婵, 高春霞, 田思泉, 等. 淀山湖鱼类群落结构多样性的年际变化[J]. 上海海洋大学学报, 2014, 23(3): 403-410. HAN C, GAO C X, TIAN S Q, et al. Analysis of annual variations for fish community structure in Dianshan Lake[J]. Journal of Shanghai Ocean University, 2014, 23(3): 403-410. |
| [33] |
朱鑫华, 吴鹤洲, 徐凤山, 等. 黄渤海沿岸水域游泳动物群落多样性及其相关因素的研究[J]. 海洋学报, 1994, 16(3): 102-112. ZHU X H, WU H Z, XU F S, et al. Studies on the diversity of swimming fauna community and its related factors in the coastal waters of the Yellow Sea and the Bohai Sea[J]. Acta Oceanologica Sinica, 1994, 16(3): 102-112. |
| [34] |
金斌松. 长江口盐沼潮沟鱼类多样性时空分布格局[D]. 上海: 复旦大学, 2010. JIN B S. Temporal and spatial distribution patterns of fish diversity in salt marsh creeks of the Yangtze River Estuary[D]. Shanghai: Fudan University, 2010. |
| [35] |
李圣法, 程家骅, 李长松, 等. 东海中部鱼类群落多样性的季节变化[J]. 海洋渔业, 2005, 27(2): 113-119. LI S F, CHENG J H, LI C S, et al. Seasonal changes on fish community diversity in the middle part of the East China Sea[J]. Marine Fisheries, 2005, 27(2): 113-119. DOI:10.3969/j.issn.1004-2490.2005.02.005 |
| [36] |
JAHANI N, NABAVI S M B, DEHGHAN MADISEH S, et al. Qualitative assessment of organic pollution from aquaculture activities on benthic organisms using ABC (Abundance Biomass Curves) in Ghazale creek (Persian Gulf)[J]. Iranian Scientific Fisheries Journal, 2011, 19(4): 43-54. |
| [37] |
YEMANE D, FIELD J G, LESLIE R W. Exploring the effects of fishing on fish assemblages using Abundance Biomass Comparison (ABC) curves[J]. ICES Journal of Marine Science, 2005, 62(3): 374-379. DOI:10.1016/j.icesjms.2005.01.009 |
2. Shanghai University Key Laboratory of Marine Animal Taxonomy and Evolution, Shanghai Ocean University, Shanghai 201306, China;
3. Shanghai Collaborative Innovation for Aquatic Animal Grenetics and Breeding, Shanghai Ocean University, Shanghai 201306, China;
4. Shanghai Chentou Raw Water Co. Ltd, Shanghai 200125, China