 2021,
Vol. 30
2021,
Vol. 30



长江口是我国沿海最大的河口区和河口渔场,入江口被崇明岛分成南支和北支2个水域。受台湾暖流、黄海水、长江冲淡水和潮汐等的共同影响,长江口的浮游生物种类丰富,群落结构复杂[1]。许多鱼类和甲壳类动物在河口区进行繁殖、生长和索饵,河口区也是许多洄游鱼类的重要通道[2]。凤鲚(Coilia mystus)和刀鲚(Coilia nasus)等经济鱼类以长江口作为重要的产卵场、保育场和庇护所[3-4]。
刀鲚捕捞是长江渔业的重要组成部分,在20世纪70年代其年捕捞量曾达到3 945 t。但近年来,由于长江生态环境的破坏、水域污染和过度捕捞,刀鲚资源量已严重下降。对于刀鲚仔稚鱼形态学、摄食习性、数量分布、日龄组成与生长等问题已经有许多学者[5-8]进行了研究,但针对长江口南支刀鲚仔稚鱼资源量变化的研究甚少,仅李安东等[9]对其数量变动进行了探讨,但缺乏年度间的相关比较。本研究在长江口南支水域定点调查的基础上,与李安东等[9]2014年和侯世挺等[4]2015年同期的调查结果作比较,分析刀鲚仔稚鱼资源量的年度变化,旨在为刀鲚早期资源的保护和恢复提供科学依据。
1 材料与方法 1.1 调查方法2018年6—10月,每月大潮期间,以大型仔稚鱼网(口径1.3 m、网目0.5 mm)在长江口南支水域设置的8个站位点(St.1~St.8,图 1)进行表层水平拖网采样调查,St.1~St.5为白天涨潮时采样,St.6~St.8为白天涨退潮采样,此外在St.8进行夜间涨退潮采样。采样渔船(“沪崇渔10688”,48.69 kW)的拖网速度为2 kn,每网次拖网时间10 min,以网口流量计(KC Denmark A/S Model 23.090)记录拖网流量值,在现场用5%的甲醛溶液固定采集到的样品,同时现场测定水温。

|
图 1 采样点分布 Fig. 1 Sampling stations |
在Olympus SZX7解剖镜下,从野外采集的样品中分离出刀鲚仔稚鱼,按照张冬良等[5]和KENDALL等[10]的方法划分刀鲚仔稚鱼的发育阶段并计数,使用游标卡尺或目测微尺进行体长测量[前弯曲期仔鱼和弯曲期仔鱼测量吻端至脊索末端的长度(NL),后弯曲期仔鱼和稚鱼测量标准体长(SL)]。
刀鲚仔稚鱼密度计算公式为
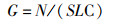 (1)
(1)
式中:G为单位体积水体中刀鲚仔稚鱼个体数即刀鲚仔稚鱼密度,尾/m3;N为每次拖网所采集到的刀鲚仔稚鱼个体数,尾;S为网口面积,m2;L为流量计转数;C为流量计校正值,C=0.3 [11]。以G表征刀鲚资源丰度,比较不同月份、不同站位点、不同年份以及昼夜涨退潮的刀鲚仔稚鱼密度。所有的统计分析均在Excel 2016、IBM SPSS Statistics 21和GraphPad Prism 7中完成。
2 结果 2.1 温度2018年6—10月,水温为24.8~30.9 ℃,8月平均水温最高,10月平均水温最低,平均水温为27.8 ℃。2014、2015和2018年各月的水温变化趋势相同,6—8月水温逐渐上升,8—10月水温逐渐下降。2018年的平均水温高于2015和2014年两年调查的平均水温。采用IBM SPSS Statistics 21进行差异性分析,表明2018年与2014和2015年水温差异性不显著(P>0.05,图 2)。

|
图 2 平均水温月变化 Fig. 2 Monthly changes of average water temperature |
本研究在长江口共拖网58次,采集刀鲚仔稚鱼4 611尾,平均密度为0.123尾/m3(表 1)。刀鲚仔稚鱼集中分布在St.1、St.2、St.5、St.6、St.8,其中:白天涨潮时St.1密度最高(0.563尾/m3),其次为St.6(0.206尾/m3);St.3、St.4、St.7以及St.8夜间涨退潮密度较低,均低于0.1尾/m3。白天涨潮时,刀鲚仔稚鱼主要出现在6月,平均密度(0.572尾/m3)高于其他各月,且以St.1密度最大(2.151尾/m3), 其次为St.6(0.951尾/m3), 7月刀鲚仔稚鱼出现量减少,平均密度为0.131尾/m3,8—10月刀鲚仔稚鱼密度较低(图 3)。退潮时,刀鲚仔稚鱼主要出现在6月的St.6,密度为0.232尾/m3(图 4)。

|
表 1 各站位涨、退潮仔稚鱼的密度变化 Tab.1 Variations of density of larval fish during flood and ebb tides in 2018 |

|
图 3 白天涨潮时刀鲚仔稚鱼密度分布 Fig. 3 Density distribution of Coilia nasus larvae and juveniles at flood tide during the day |

|
图 4 白天退潮时刀鲚仔稚鱼密度分布 Fig. 4 Density distribution of Coilia nasus larvae and juveniles at ebb tide during the day |
St.8调查结果显示,白天涨潮期共采集刀鲚仔稚鱼287尾,平均密度(0.191尾/m3)高于退潮期(0.005尾/m3);夜晚采集刀鲚仔稚鱼较少,涨潮期平均密度(0.005尾/m3)也高于退潮期(0.002尾/m3)。从昼夜比较看,白天涨潮期刀鲚仔稚鱼的平均密度大于夜晚涨潮期平均密度,白天退潮期密度同样大于夜晚退潮期密度(图 5)。

|
图 5 St.8刀鲚仔稚鱼昼夜涨退潮密度变化 Fig. 5 Average density of Coilia nasus larvae and juveniles at St.8 |
比较各站位的刀鲚仔稚鱼数量变化:2015年刀鲚仔稚鱼出现量较少,仅539尾,平均密度为0.019尾/m3,以St.1平均密度(0.043尾/m3)最高;而2014年采集刀鲚仔稚鱼1 270尾,平均密度为0.029尾/m3,St.8平均密度(0.079尾/m3)大于其他各站位点密度。2018年各站位间刀鲚仔稚鱼平均密度高于2014和2015年。分析表明,2018与2014、2015年各站位点平均密度差异均不显著(P>0.05)。由此可见,年度间刀鲚仔稚鱼在各站位的出现波动较小,见图 6。

|
图 6 刀鲚仔稚鱼各站位点密度年变化 Fig. 6 Annual changes of average density of Coilia nasus larvae and juveniles by stations |
2018年在长江口南支水域采集的刀鲚仔稚鱼体长范围为3.1~61.74 mm,但体长4~17 mm刀鲚仔鱼占95.26%。发育阶段从前弯曲期到稚鱼期,没有采集到卵黄囊期仔鱼,其中:前弯曲期仔鱼(3.1~18.12 mm)数量最多,占总渔获量的86.18%;弯曲期仔鱼(4.9~24.05 mm)和后弯曲期仔鱼(9.75~30.36 mm)分别占8.50%和4.60%;稚鱼期(24.22~61.7 mm)数量最少,仅占0.72%。见图 7。

|
图 7 刀鲚仔稚鱼数量和体长分布 Fig. 7 Number and body length of Coilia nasus larvae and juveniles |
2015年采集的刀鲚仔稚鱼体长范围为3.86~57.44 mm,前弯曲期、弯曲期和后弯曲期的仔鱼各占总渔获量30%左右。2014年刀鲚仔稚鱼的体长范围为3.56~50.88 mm,卵黄囊期和后弯曲期仔鱼数量较多,各占总渔获量的37.17%和20.16%。由此可见,3年采集的刀鲚仔稚鱼体长范围基本一致,但各发育阶段仔稚鱼占比存在差异,前弯曲期仔鱼在2018和2015年所占比例较高,而2014年卵黄囊期仔鱼占比较高。见图 8。

|
图 8 各年刀鲚仔稚鱼发育阶段组成 Fig. 8 Development stages composition of Coilia nasus larvae and juveniles |
比较2018、2015和2014年各发育阶段仔稚鱼出现时间和高峰期,卵黄囊期仔鱼在2014年6月大量出现,占该月仔稚鱼总数的60.31%,7月采集数量较少,后弯曲期仔鱼和稚鱼主要出现在8月;而2018和2015年均未采集到卵黄囊期的刀鲚,2015年7月采集到较多数量的前弯曲期、弯曲期和后弯曲期刀鲚,而2018年6月采集大量前弯曲期、弯曲期和后弯曲期的刀鲚,占本次调查总渔获量的81.98%,7—10月仅采集到少量刀鲚仔稚鱼。比较2018、2015和2014年发育阶段变化可知,各发育阶段的刀鲚仔稚鱼大量出现的时间有所提前,尤其是卵黄囊期仔鱼,因此推测刀鲚的产卵时间可能比往年早。见图 9。

|
图 9 刀鲚仔稚鱼发育阶段组成月变化 Fig. 9 Monthly changes of development stages composition of Coilia nasus larvae and juveniles |
2018年白天涨潮时:6月采集刀鲚仔稚鱼3 512尾,体长集中分布在6~12 mm,前弯曲期仔鱼数量最多,而其他各时期的刀鲚数量较少;7月前弯曲期仔鱼出现频率下降,但仍高于其他各发育阶段仔稚鱼,弯曲期仔鱼出现量增加;8月仅采集刀鲚仔稚鱼49尾,以后弯曲期仔鱼为主。白天退潮时:共采集339尾刀鲚仔稚鱼,其中6月采集数量最多,为268尾,各发育时期的仔稚鱼出现频率与白天涨潮时基本一致,但其数量低于白天涨潮。见图 10。

|
图 10 白天涨退潮刀鲚仔稚鱼体长和发育阶段组成 Fig. 10 Body length frequencies and developmental stages of Coilia nasus larvae and juveniles |
白天涨潮时:2014年6月出现频率最高的为卵黄囊期仔鱼,仔稚鱼体长集中分布在4~6 mm,前弯曲期仔鱼出现量较低;7月仅采集到19尾刀鲚仔稚鱼,各发育时期的仔稚鱼数量相差不大;8月采集的刀鲚仔稚鱼发育阶段主要为弯曲期、后弯曲期和稚鱼期为主。2015年各发育时期的刀鲚出现频率与2018年相似,6月以前弯曲期仔鱼为主。白天退潮时:2014年和2015年的6月均未采集到刀鲚仔稚鱼,且两年7月均以后弯曲期刀鲚为主。比较3年数据可知,刀鲚仔稚鱼在3年各月的发育阶段组成差异不大。见图 10。
3 讨论 3.1 环境因子对刀鲚仔稚鱼密度的影响仔稚鱼的分布受多种因素的影响,生态环境因子和仔稚鱼自身的生物特性影响其时空分布和数量变化,而温度是影响仔稚鱼数量分布的重要因素[12]。研究[13]表明,刀鲚受精卵孵化的最适宜水温为25~27 ℃,5月下旬至10月上旬为刀鲚的产卵时期,其中5月末至8月上旬为产卵盛期[8]。2018年6月长江口的水温(26.2 ℃)最适宜刀鲚受精卵孵化,刀鲚仔稚鱼大量出现,平均密度最高;6—8月,水温逐渐上升,8月水温最高(30.9 ℃),高于受精卵的最适孵化水温,因此仔稚鱼密度随水温的上升而逐月下降;8—10月,水温逐渐下降,但此时刀鲚的产卵盛期已过,仔稚鱼密度继续下降。2015年6月长江口的水温(23.3 ℃)低于刀鲚受精卵孵化的最适温度,受精卵孵化率较低,因此仔稚鱼的出现量较低,随着温度的上升,受精卵的孵化率提高,7—8月数量增加。
刘熠等[14]在长江下游的研究表明,沿岸地形和水体动力学差异可能是造成刀鲚仔稚鱼在不同江段的断面空间上密度分布不同的原因。本研究显示:2018、2015以及2014年刀鲚仔稚鱼主要分布在St.1和St.8,其中St.1位于倾斜狭窄的南支江段处,北支和南支水流在此交汇,水流湍急,造成泥沙沉积;而St.8位于下游入江处,江水的深度较浅,刀鲚仔稚鱼主要出现在这两个站位点可能与这些特殊的地理特征有关。侯世挺等[4]在长江口南支进行的研究也表明,仔稚鱼密度较高的两个站位点分别是St.8和St.1。
潮汐不仅可以调节水体间的差异和温度梯度,还可以使流速、流向、水深等发生改变,从而对鱼类的行为产生较大的影响[15]。研究[16-18]表明,仔稚鱼可以利用潮汐变化在涨潮时随水流进入河口区生长发育。WEINSTEIN等[19]提出了选择性潮汐输送机制,牙鲆(Paralichthys spp.)和波纹绒须石首鱼(Micropogonia sundulatus)的后期仔鱼在白天涨潮时移向较浅水层,退潮时移向较深水层,而在夜晚涨潮时牙鲆向较浅水层移动,波纹绒须石首鱼却停留在较深水层。李联飞等[20]在长江口南支水域进行的刀鲚仔稚鱼分层调查表明,涨潮时前弯曲期和弯曲期的刀鲚在表层的CPUE大于中层,而后弯曲期和稚鱼期的刀鲚则趋于中层分布。本研究对St.6、St.7和St.8的刀鲚密度调查显示,涨潮期仔稚鱼的平均密度大于退潮期,可能与刀鲚仔稚鱼的垂直分布有关。曾娇等[21]在长江口南支水域进行的潮汐对仔稚鱼密度影响的研究也表明各月仔稚鱼涨潮期密度均大于退潮期密度。
发育阶段靠前的仔稚鱼由于鳍条尚未发育完全,游泳能力较弱,会随着潮汐进行流动,随着仔稚鱼生长发育,仔稚鱼游泳能力增强,从而能自主选择方向[21-22]。比较2018、2015以及2014年采集的刀鲚仔稚鱼优势发育阶段,2018和2015年以前弯曲期为主,而2014年以卵黄囊期仔鱼为主。卵黄囊期仔鱼的体长集中在4~6 mm,前弯曲期仔鱼的体长集中在6~10 mm,3年间这两个发育阶段的数量差异较大,可能是因为卵黄囊期仔鱼和前弯曲期仔鱼的个体较小,容易受到潮汐、气候变化、水流和天敌等的影响,而体长相对较长的弯曲期仔鱼和后弯曲期仔鱼以及稚鱼在这3年间出现的时期和频率基本一致。葛珂珂等[7]和李联飞等[20]提出刀鲚仔稚鱼的体长达到一定范围或特定的发育阶段(弯曲期和后弯曲期)之后就会进入碎波带进行生长发育。本研究的卵黄囊期仔鱼和前弯曲期仔鱼数量较高,而弯曲期仔鱼、后弯曲期仔鱼以及稚鱼较少,这些体长较大的仔稚鱼可能已从长江口敞水水域进入到碎波带栖息。
3.2 刀鲚仔稚鱼资源的保护钟俊生等[23]和吴美琴等[24]的研究表明刀鲚仔稚鱼在长江口仔稚鱼群落中占优势地位,董文霞等[25]的研究也表明刀鲚种群依旧具有生长潜力。但2008—2015年,刀鲚成鱼在长江口汛期的总捕捞量呈下降趋势,仅在2010和2013年出现波动性回升,且个体趋于小型化[26-28],资源状况仍十分严峻。为保护长江流域水生生物资源,2015年12月农业农村部将长江流域的禁渔期从之前的3个月调整到4个月。从本研究的刀鲚仔稚鱼各发育阶段出现的时间和高峰期看,也可以推测刀鲚的产卵时间比以往有所提前,而禁渔期的延长将有利于刀鲚早期资源的保护。
自然死亡、环境污染、过度捕捞和鳗苗网等对刀鲚资源量的影响均非常明显。葛成冈等[28]的研究表明,长江口数量庞大的小网目(1 mm)鳗苗网对经济鱼类的仔稚幼鱼产生了直接影响,对洄游性鱼类幼体的损害极大,对刀鲚资源量的锐减造成了极大影响,因此这类网具应予以取缔。农业农村部已规定从2017年7月1日起长江干流水域的准用渔具及其最小尺寸并从2019年2月1日起停止发放刀鲚专项捕捞许可证的措施,必将对恢复刀鲚资源起到决定性的作用。
| [1] |
KNOX G A. Estuarine ecosystems: a systems approach[M]. Boca Raton, FL: CRC Press, 1986.
|
| [2] |
LASIAK T A. Nursery grounds of juvenile teleosts: evidence from the surf zone of King's Beach, Port Elizabeth[J]. South African Journal of Science, 1981, 77(9): 388-390. |
| [3] |
MODDE T, ROSS S T. Trophic relationships of fishes occurring within a surf zone habitat in the northern Gulf of Mexico[J]. Northwest Gulf Science, 1983, 6(2): 109-120. |
| [4] |
侯世挺, 钟俊生, 王晓东, 等. 长江口南支仔稚鱼数量随潮汐变化的研究[J]. 上海海洋大学学报, 2018, 27(2): 250-258. HOU S T, ZHONG J S, WANG X D, et al. The study on the amount changes of fish larvae during flood and ebb in the south branch of Yangtze River estuary[J]. Journal of Shanghai Ocean University, 2018, 27(2): 250-258. |
| [5] |
张冬良, 李黎, 钟俊生, 等. 长江口碎波带刀鲚仔稚鱼的形态学研究[J]. 上海海洋大学学报, 2009, 18(2): 150-154. ZHONG D L, LI L, ZHONG J S, et al. Morphological study on larvae and juveniles of Coilia nasus in the surf zone of Yangtze River estuary[J]. Journal of Shanghai Ocean University, 2009, 18(2): 150-154. |
| [6] |
蒋雪莲, 张宇, 钟俊生, 等. 长江口沿岸碎波带刀鲚仔稚鱼摄食习性与浮游动物分布的相关性研究[J]. 长江流域资源与环境, 2015, 24(9): 1507-1513. JIANG X L, ZHANG Y, ZHONG J S, et al. Study on relationship between distribution of zooplankton and Coilia nasus larvae feeding features in the surf zone of Yangtze River estuary[J]. Resources and Environment in the Yangtze Basin, 2015, 24(9): 1507-1513. DOI:10.11870/cjlyzyyhj201509010 |
| [7] |
葛珂珂, 钟俊生, 吴美琴, 等. 长江口沿岸碎波带刀鲚仔稚鱼的数量分布[J]. 中国水产科学, 2009, 16(6): 923-930. GE K K, ZHONG J S, WU M Q, et al. Distribution and abundance of larval Japanese grenadier anchovy Coilia nasus in the surf zone of the Yangtze River estuary[J]. Journal of Fishery Sciences of China, 2009, 16(6): 923-930. DOI:10.3321/j.issn:1005-8737.2009.06.013 |
| [8] |
葛珂珂, 钟俊生. 长江口沿岸碎波带刀鲚仔稚鱼的日龄组成与生长[J]. 水生生物学报, 2010, 34(4): 716-721. GE K K, ZHONG J S. Daily-age structure and growth characteristics of Coilia nasus larvae and juveniles in the surf zone of Yangtze River estuary[J]. Acta Hydrobiologica Sinica, 2010, 34(4): 716-721. |
| [9] |
李安东, 钟俊生, 罗一鸣, 等. 长江口南支水域刀鲚仔稚鱼数量变动的研究[J]. 上海海洋大学学报, 2015, 24(5): 745-753. LI A D, ZHONG J S, LUO Y M, et al. Distribution and abundance of Coilia nasus larvae and juveniles in the southern branch of Yangtze River estuary[J]. Journal of Shanghai Ocean University, 2015, 24(5): 745-753. |
| [10] |
KENDALL A W, AHLSTROM E H JR, MOSER H G. Early life history stages of fishes and their characters[M]. Lawrence: The American Society of Ichthyologists Herpetologists, Special Publications, 1984: 11-22.
|
| [11] |
国家海洋局908专项办公室. 海洋生物生态调查技术规程[M]. 北京: 海洋出版社, 2006: 908. Office of the State Oceanic Administration. Technical specification for ecological surveys of marine life[M]. Beijing: Ocean Press, 2006. |
| [12] |
蒋玫, 沈新强, 陈莲芳. 长江口及邻近水域春季鱼卵仔鱼分布与环境因子的关系[J]. 海洋环境科学, 2006, 25(2): 37-39, 44. JIANG M, SHEN X Q, CHEN L F. Relationship between with abundance distribution of fish eggs, larvae and environmental factors in the Changjiang estuary and vicinity waters in spring[J]. Marine Environmental Science, 2006, 25(2): 37-39, 44. DOI:10.3969/j.issn.1007-6336.2006.02.010 |
| [13] |
施永海, 张根玉, 张海明, 等. 刀鲚的全人工繁殖及胚胎发育[J]. 上海海洋大学学报, 2015, 24(1): 36-43. SHI Y H, ZHANG G Y, ZHANG H M, et al. Complete artificial propagation and embryonic development of Coilia nasus[J]. Journal of Shanghai Ocean University, 2015, 24(1): 36-43. |
| [14] |
刘熠, 任鹏, 杨习文, 等. 长江下游刀鲚(Coilia nasus)仔稚鱼的时空分布[J]. 湖泊科学, 2020, 32(2): 506-517. LIU Y, REN P, YANG X W, et al. Spatial and temporal distribution of larvae and juvenile Coilia nasus in the lower reaches of the Yangtze River[J]. Journal of Lake Sciences, 2020, 32(2): 506-517. |
| [15] |
乔云贵, 黄洪亮, 黄妙芬, 等. 潮汐对鱼类游泳行为影响的研究进展[J]. 江苏农业科学, 2012, 40(3): 9-12. QIAO Y G, HUANG H L., HUANG M F, et al. Research progress on the effect of tide on fish swimming behavior[J]. Jiangsu Agricultural Sciences,, 2012, 40(3): 9-12. DOI:10.3969/j.issn.1002-1302.2012.03.004 |
| [16] |
KINGSFORD M J, CHOAT J H. Influence of surface slicks on the distribution and onshore movements of small fish[J]. Marine Biology, 1986, 91(2): 161-171. DOI:10.1007/BF00569432 |
| [17] |
SHANKS A L. Surface slicks associated with tidally forced internal waves may transport pelagic larvae of benthic invertebrates and fishes shoreward[J]. Marine Ecology Progress Series, 1983, 13(2/3): 311-315. |
| [18] |
LAPRISE R, DODSON J J. Ontogeny and importance of tidal vertical migrations in the retention of larval smelt Osmerus mordax in a well-mixed estuary[J]. Marine Ecology Progress Series, 1989, 55(2/3): 101-111. |
| [19] |
WEINSTEIN N, MICHAEL P. Population dynamics of an estuarine-dependent fish, the spot (Leiostomus xanthurus), along a tidal creek-seagrass meadow coenocline[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1983, 40(10): 1633-1638. DOI:10.1139/f83-189 |
| [20] |
李联飞, 钟俊生, 曾娇, 等. 潮汐对长江口南支刀鲚仔稚鱼垂直分布及昼夜变化的影响[J]. 上海海洋大学学报, 2020, 29(1): 74-82. LI L F, ZHONG J S, ZENG J, et al. Vertical distribution and changes during day and night of Coilia nasus larvae and juveniles depending on flood and ebb tide in southern branch of Yangtze River estuary[J]. Journal of Shanghai Ocean University, 2020, 29(1): 74-82. |
| [21] |
曾娇. 长江口南支涨退潮对仔稚鱼垂直分布及漂流机制的影响研究[D]. 上海: 上海海洋大学, 2019. ZENG J. Effects of tide on vertical distribution and drifting mechanisms of the fish larvae and juveniles in the southern branch of the Yangtze estuary[D]. Shanghai: Shanghai Ocean University, 2019. |
| [22] |
唐明英, 黄德林, 黄立章, 等. 草、青、鲢、鳙鱼卵水力学特性试验及其在三峡库区孵化条件初步预测[J]. 水利渔业, 1989(4): 26-30. TANG M Y, HUANG D L, HUANG L Z, et al. Hydraulic characteristics test of grass crap, black crap, silver crap and bighead carp eggs and preliminary prediction of hatching conditions in the Three Gorges reservoir area[J]. Reservoir Fisheries, 1989(4): 26-30. |
| [23] |
钟俊生, 郁蔚文, 刘必林, 等. 长江口沿岸碎波带仔稚鱼种类组成和季节性变化[J]. 上海水产大学学报, 2005, 14(4): 375-382. ZHONG J S, YU W W, LIU B L, et al. Seasonal occurrences of fish larvae and juveniles in the surf zone of the Yangtze River estuary[J]. Journal of Shanghai Fisheries University, 2005, 14(4): 375-382. |
| [24] |
吴美琴, 钟俊生, 葛珂珂, 等. 长江口沿岸碎波带仔稚鱼分布的季节性变动[J]. 渔业科学进展, 2010, 31(1): 1-7. WU M Q, ZHONG J S, GE K K, et al. Seasonal fluctuation of fish larvae and juveniles distribution in the surf zone of the Yangtze River estuary[J]. Progress in Fishery Sciences, 2010, 31(1): 1-7. DOI:10.3969/j.issn.1000-7075.2010.01.001 |
| [25] |
董文霞, 唐文乔, 王磊. 长江刀鲚繁殖群体的生长特性[J]. 上海海洋大学学报, 2014, 23(5): 669-674. DONG W X, TANG W Q, WANG L. Growth characteristics of reproductive population of Coilia nasus in the Yangtze River[J]. Journal of Shanghai Ocean University, 2014, 23(5): 669-674. |
| [26] |
田思泉, 田芝清, 高春霞, 等. 长江口刀鲚汛期特征及其资源状况的年际变化分析[J]. 上海海洋大学学报, 2014, 23(2): 245-250. TIAN S Q, TIAN Z Q, GAO C X, et al. Analyzing of annual changes for the stock status of Coilia nasus in fishing season in Yangtze River estuary[J]. Journal of Shanghai Ocean University, 2014, 23(2): 245-250. |
| [27] |
刘凯, 段金荣, 徐东坡, 等. 长江口刀鲚渔汛特征及捕捞量现状[J]. 生态学杂志, 2012, 31(12): 3138-3143. LIU K, DUAN J R, XU D P, et al. Present situation of Coilia nasus population features and yield in Yangtze River estuary waters in fishing season[J]. Chinese Journal of Ecology, 2012, 31(12): 3138-3143. |
| [28] |
葛成冈, 钟俊生, 葛珂珂, 等. 长江口鳗苗网兼捕鱼类组成分析及其管理建议[J]. 上海海洋大学学报, 2013, 22(3): 391-397. |