 2020,
Vol. 29
2020,
Vol. 29



2. 上海海洋大学 农业农村部鱼类营养与环境生态研究中心, 上海 201306;
3. 上海水产养殖工程技术研究中心, 上海 201306
浮游植物作为江河湖库等水体生态系统的主要初级生产者,是浮游动物、底栖动物以及鱼类等生物类群的主要食物来源,同时也是水生态系统中能量流动和物质循环的驱动因子[1],在一定程度上可决定物质循环和能量流动的方式[2-3]。浮游植物对水质变化敏感,环境因子变化对浮游植物生理活动及其群落结构(如物种组成和数量)和功能的影响很大,其群落结构特征可以较准确地反映水体的水质状况及其动态变化,是水环境质量的重要指示类群及其评价的重要指标[4-9]。
河道对城市的防洪排涝、森林防火、引水灌溉、水运通航、休憩观赏和生态平衡等具有十分重要的作用,城市河流作为资源和环境的载体,关系到城市的生存和发展,是影响城市风格和环境质量的重要因素[10]。城市河道由于生物群落结构简单,食物链不合理,易受到外界各种人为或非人为因素的干扰,往往缺乏自我净化与维持的能力[5],导致水环境质量现状差,水生态系统非常脆弱。迄今为止,国内外对内陆水体浮游植物群落及其在水环境质量评价方面的研究主要集中在湖泊、水库和河流[8, 11-19],对城市河道开展的较少[5, 9-10, 20-24]。
快速的城市化给城市带来了一些生态环境问题,环城绿带作为抑制城市蔓延和促进生态环境保护的有效办法受到了广泛关注[25]。上海在国内首次提出了环城绿带思想并于1995年开始建设环城绿带[25]。目前,学者集中对上海环城绿带的林带植物和土壤进行一些研究[26],但鲜见对该绿带河道水生生物方面的研究。本研究于2017年对上海环城绿带11条休憩型河道的浮游植物进行了调查,比较了沉水植物对河道浮游植物群落结构特征的影响,并分析其结构特征与环境因子的关系和进行水质生物学评价,以期为上海市河道生态修复以及水环境质量评价提供基础数据和参考依据。
1 材料与方法 1.1 研究区概况根据上海市河道分类情况,沿中环绿带选取11条生态休憩型河道作为采样站点,每条河道设置1个采样点(图 1)。其中S3位于宝山区,S5位于普陀区,S7位于嘉定区,S9、S10、S11位于长宁区,S15、S18、S21、S24和S25位于浦东新区。S3、S11、S18和S24水体中均种植有沉水植物,S11除了有沉水植物外还进行了河道淤泥清除工程,在有水草河段的中心位置采样,其他7个站点为开放型河道,游客可在周边休憩玩耍。

|
图 1 上海市环城绿带休憩型河道浮游植物采样站点分布 Fig. 1 Distribution of phytoplankton sampling stations in river courses for leisure of green belt around city in Shanghai |
于2017年5月、7月、10月、12月中旬采集浮游植物样品。浮游植物定量样品采集表底混合水样1 000 mL,加1.5%的鲁哥试液固定,静置24~48 h后浓缩至30~50 mL,在显微镜下进行种类鉴定[26],然后计算浮游植物的物种组成、生物密度和生物量。
采集浮游植物样品时,测定并记录溶解氧(DO)、水温(WT)、pH和透明度(SD)。依据《水和废水监测分析方法》对总氮(TN)、总磷(TP)、高锰酸盐指数(CODMn)、氨氮(NH3-N)、五日生化需氧量(BOD5)进行分析测定,叶绿素a采用丙酮萃取分光光度计法[27]。同时,采样河道较浅(0.74±0.06 m),通过直接观察及底栖动物和沉积物采集记录河道是否有沉水植物存在。经计算,中环绿带11条河流的水温平均为(19.56±0.73) ℃,pH为7.92±0.05,SD为(0.54±0.03) m,DO为(7.84±0.27) mg/L,TN为(2.47±0.16) mg/L,TP为(0.15±0.02) mg/L,NH3-N为(1.06±0.10) mg/L,BOD5为(5.88±0.26) mg/L,CODMn为(7.88±0.34) mg/L。
1.3 多样性计算及水质评价采用Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(J)和Margalef丰富度指数(R)来评价河道水质[6]:
 (1)
(1)
 (2)
(2)
 (3)
(3)
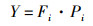 (4)
(4)
式中:Pi=ni/N,为第i种藻类的个数;N为所有藻类总个数;S为样品中藻类种类数;Fi为第i种藻类在各站点出现的频率,将优势度指数Y>0.02定为优势种[5]。
将H′划分为4级:0~1为重污染,1~2为中污染,2~3为轻污染,H′>3为清洁或无污染[5];将J划分为4级:0~0.3为重度污染,0.3~0.5为中度污染,0.5~0.8为轻度污染,0.8~1.0为清洁[5];将R划分为5级:0~1为多污型,1~2为α-中污型,2~3为β-中污型,3~4为寡污型,R>5为清洁无污染水质[22]。
采用综合营养状态指数(TLI)法进行水体富营养化程度评价,选择Chl.a、TP、TN、SD和CODMn等5个参数计算TLI:
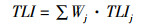 (5)
(5)
式中:Wj代表第j种参数的营养状态指数的相关权重,TLIj代表第j种参数的营养状态指数。TLI(Chl.a)=10×(2.5+1.086×lnChl.a),TLI(TP)=10×(9.436+1.624×lnTP),TLI(TN)=10×(5.453+1.694×lnTN),TLI(SD)=10×(5.118-1.94×lnSD),TLI(CODMn)=10×(0.109+2.661×lnCODMn)。TLI<30为贫营养,30~50中营养,50~60轻度富营养化,60~70中度富营养化,>70重度富营养化。
1.4 统计分析采用双因素方差分析探讨浮游植物现存量和多样性的季节变化和群落类型间的差异性,数据分析前采用对数、平方根或立方根转换以满足正态分布和方差齐性。若存在交互作用,则控制季节采用非参数检验(Mann-Whitley U test)探讨群落类型间的差异,或控制群落类型采用单因素方差分析探讨季节间差异。多重比较采用Duncan氏检验。
采用聚类分析(CA)和多维尺度分析(MDS)探讨沉水植物对浮游植物群落的影响,运行Primer 5.0软件进行CA分析和MDS分析。采用压力系数(Stress)检验MDS的拟合优度,当stress≤0.1时,说明具有较好的拟合度;如果stress < 0.05时,认为该图形具有代表性。同时采用ANOSIM检验群落间是否存在显著差异,并应用相似性百分比分析(SIMPER)探讨浮游植物物种对群落类型内相似性和类型间相异性的平均贡献率。以上所有分析基于出现频次≥2且相对密度≥1%的物种矩阵,对相对密度数据进行反正弦平方根转换,并以欧式距离作为相异性指标。
运行Canoca 4.53软件分析浮游植物物种组成与环境因子的关系,去趋势对应分析(DCA)结果显示使用冗余分析(RDA)更为合理。河道是否有沉水植物的数据也参与RDA分析,有沉水植物赋值1,无沉水植物河道赋值0。DCA和RDA分析前对每个站点的浮游植物密度进行平均,选择出现频次≥2且N%≥1%的物种,并对物种密度和pH除外的环境因子数据进行对数转换。RDA运行中对环境因子进行前向选择和蒙特卡洛检验(999次)。
2 结果 2.1 浮游植物群落物种组成共调查到浮游植物8门83属142种,其中绿藻门32属61种,占43.0%;硅藻门有23属40种,占28.2%;蓝藻门13属19种,占13.4%;其他门共占比15.5%,包括裸藻门、甲藻门、隐藻门、黄藻门和金藻门,分别有5属9种、4属5种、3属5种、2属2种和1属1种;有6个优势种,其中微小平裂藻优势度最高(表 1)。

|
表 1 浮游植物不同群落类型优势种 Tab.1 The dominant species in different phytoplankton community types |
聚类分析表明,中环绿带休憩型河道的浮游植物群落可以划分为2种类型(图 2),有沉水植物河道的S3、S11、S18和S24站点聚为一类(WSM),其他无沉水植物河道的7个站点聚为一类(NWSM)。MDS分析显示压力系数为0.1,证明图形有较好的拟合度,此群落划分可行。ANOSIM也揭示浮游植物群落在有无水草河道间存在显著差异(R=0.651,P < 0.01)。SIMPER分析表明,WSM类型内平均相似性为46.2%,其中尖尾蓝隐藻(Chroomonas acuta,34.3%)、小球藻(Chlorella vulgaris,10.2%)、伪鱼腥藻(Pseudoanabaena,8.9%)、直链藻(Melosira,7.4%)、小环藻(Cyclotella sp.,5.6%)、啮噬隐藻(Cryptomonas erosa,5.5%)的贡献较大;NWSM群落类型内平均相似性为57.9%,其中微小平裂藻(Merismopedia tenuissima,14.1%)、伪鱼腥藻(14.1%)、颤藻属(Oscillatoria sp.,12.4%)、直链藻(8.0%)、小球藻(6.8%)和尖尾蓝隐藻(5.1%)的贡献率较高。两种群落类型间的平均相异性为59.0%,主要分歧种为尖尾蓝隐藻(14.0%)、微小平裂藻(11.3%)和颤藻属(7.9%)。

|
图 2 浮游植物物种组成空间聚类分析 Fig. 2 Spatial clustering analysis of phytoplankton species composition |
WSM类型河道绿藻门和硅藻门物种数较多均有22种,NWSM类型河道绿藻门物种数最多有58种,其次为硅藻门37种,其他浮游植物种类在WSM和NWSM类型河道间相差不大,但在WSM类型河道中发现清洁水体指示物种,即金藻门锥囊藻属1种(Dinobryon sp.),而在NWSM类型河道未发现金藻门。WSM类型河道有4个优势种,尖尾蓝隐藻优势度最大;NWSM类型河道有5个优势种,其中微小平裂藻优势度最高(表 1)。
2.3 浮游植物现存量季节变化及其在不同类型河道间的差异性浮游植物年均密度为(17.10±0.64)×106 ind/L,蓝藻为密度绝对优势门类,占总密度的77.1%,其次为硅藻(9.0%)和绿藻(8.9%)(图 3)。年均生物量为(3.28±0.58) mg/L,硅藻生物量最高(38.3%),其次为甲藻(21.9%)和蓝藻(18.3%)。见图 3。

|
Ba: Bacillariophyta, Xa: Xanthophyta, Py: Pyrrophyta, Chr: Chrysophyta, Cy: Cyanophyta, Chl: Chlorophyta, Eu: Euglenophyta, Cr: Cryptophyta 图 3 浮游植物各个门类密度和生物量比例 Fig. 3 The proportions of the density and biomass of different phytoplankton phyla |
蓝藻门中的优势种包括微小平裂藻、颤藻和伪鱼腥藻,其密度在夏季较高,秋季其次,春冬季较低;小球藻夏冬秋三季较高,春季较低;尖尾蓝隐藻、小环藻和直链藻秋冬季较高,春夏季较低;啮噬隐藻秋季较高,其他三季较低(图 4)。

|
Mte.微小平裂藻Merismopedia tenuissima; Osc.颤藻属1种Oscillatoria sp.; Pse.伪鱼腥藻Pseudoanabaena sp.; Cvu.小球藻Chlorella vulgaris; Cyc.小环藻Cyclotella sp.; Mel.直链藻Melosira; Cac.尖尾蓝隐藻Chroomonas acuta; Cer.啮噬隐藻Cryptomonas erosa 图 4 河道主要浮游植物物种密度的月份变化 Fig. 4 Monthly density change of the dominant phytoplankton species in our studied rivers |
双因素方差分析表明,密度在有无水草类型河道(F=49.127,P < 0.001)和季节(F=7.803,P < 0.001)间均存在显著差异,生物量在有无水草类型河道间存在显著差异(F=10.569,P=0.002 < 0.01),但在季节间无显著差异(F=0.970,P=0.417>0.05),同时两者在不同河道类型和季节间均不存在交互作用(密度:F=2.126,P=0.114;生物量:F=0.519,P=0.672)。
WSM类型河道的浮游植物密度[(1.81±0.57)×106 ind/L]显著低于NWSM类型河道[(25.84±9.73)×106 ind/L],WSM类型河道中隐藻(占39.6%)、蓝藻(24.1%)、硅藻(19.2%)密度较高,NWSM类型河道中蓝藻密度占绝对优势(79.2%),其次为绿藻(8.8%)和硅藻(8.6%)。WSM类型河道生物量[(2.00±1.01) mg/L]显著低于NWSM类型河道[(4.00±0.67) mg/L],WSM类型河道中甲藻门生物量占优势(占70.5%),硅藻(12.1%)和隐藻(9.0%)次之,NWSM类型河道硅藻生物量最高(占45.8%),其次为蓝藻(22.9%)和绿藻(14.0%)。
Duncan氏多重比较表明,夏季浮游植物密度(48.95±23.05×106 cell/L)显著高于春季(1.99±0.66×106 cell/L)和冬季(6.14±1.78×106 cell/L),秋季(11.33±0.5.51 ×106 cell/L)显著高于春季,其他季节两两间无显著差异。
2.4 多样性指数时空变化双因素方差分析表明,R(F=57.517,P < 0.001)和H′(F=8.834,P=0.005 < 0.01)在不同类型河道间存在显著差异,无季节性差异(R:F=1.606,P=0.205>0.005;H′:F=1.323,P=0.282>0.05);J在不同类型河道间无显著差异(F=0.821,P=0.371),但存在显著的季节性差异(F=5.885,P=0.002 < 0.01);R在河道类型和季节间无交互作用(F=0.767,P=0.520>0.005),但H′(F=6.128,P=0.002 < 0.01)和J(F=6.692,P=0.001 < 0.01)存在显著的交互作用(图 5)。

|
图 5 浮游植物多样性在群落类型间差异及月份变化 Fig. 5 Differences of diversity indices of phytoplankton between community types and their seasonal changes |
NWSM类型河道的R显著高于WSM类型河道。曼-惠特尼U检验显示,春季WSM类型河道的H′显著高于NWSM(Mann-Whitley U test,Z=2.646,P=0.006 < 0.01),秋季亦然(Z=2.646,P=0.006 < 0.01),夏季(Z=1.701,P=0.109>0.05)和冬季(Z=1.890,P=0.073>0.05)两个季节H′在类型河道类型间无差异。单因素方差分析表明,WSM类型河道的J不存在显著季节差异(F=2.503,P=0.109>0.05),但NWSM类型河道季节间差异显著(F=15.512,P < 0.001);Duncan氏多重比较表明,NWSM类型河道夏季的J值明显低于其他3个季节,其他季节两两间差异不显著(图 5)。
2.5 浮游植物现存量与环境因子的相关性分析相关性分析表明,浮游植物密度与BOD5、CODMn、NH3-N、TN和TP显著正相关,生物量与BOD5和TN显著正相关(表 2)。

|
表 2 浮游植物现存量和多样性指数与环境因子间的相关系数 Tab.2 Correlation coefficients between standing crop and diversity and environmental factors |
RDA前2轴特征值分别为0.560和0.066,解释了上海河道浮游植物群落总变异的62.5%。轴1和轴2浮游植物物种与环境因子的相关性分别达到了0.970和0.804,表明物种与环境因子具有较强的相关性。第1轴主要与BOD5负相关(-0.916)、与沉水植物正相关(0.835),第2轴与沉水植物(0.408)和BOD5(0.263)均正相关。大部分浮游植物物种出现在BOD5比较高的水域,在有沉水植物的河道大部分物种的密度比较低。
2.7 水质评价环城绿带休憩型有沉水植物河道的R为0.866±0.090、H′为1.670±0.123、J为0.678±0.042,分别显示为这类河道处在多污型、中污染状态和轻度污染。无沉水植物河道R为1.653±0.059、H′为2.105±0.102,J为0.642±0.032,分别说明此类河道分别处于α-中污型、轻污染和轻度污染。
综合来看,有和无沉水植物河道浮游植物群落的R平均为1.367±0.076、H′为1.947±0.084、J为0.655±0.025,分别显示研究河道处于α-中污型、中污染状态和轻污染状态。WSM类型河道的TLI值为55.35,表明水质为轻度富营养化;NWSM类型河道TLI为68.30,指示水质为中度富营养化。

|
Aha:集星藻Actinastrum hantzschii,Ana:鱼腥藻属Anabaena,Cac:尖尾蓝隐藻Chroomonas acuta,Cer:啮噬隐藻Cryptomons erosa,Cte:四足十字藻Crucigenia tetrapedia,Cyc:小环藻Cyclotella sp,Cvu:小球藻Chlorella vulgaris,Eug:裸藻属Euglena,Gla:华美十字藻Grucigenia lauterbornii,Mva:变异直链藻Melosira varians,Mic:微囊藻属Microcystis,Mte:微小平裂藻Merismopedia tenuissima,Mel:直链藻Melosira,Nav:舟形藻属Navicula,Osc:颤藻属Oscillatoria,Opr:巨颤藻Oscillatoria princeps,Pha:扁裸藻Phacus,Spi:螺旋藻属Spirulina,Pse:伪鱼腥藻属Pseudanabaena,Syn:黄群藻属Synura,Squ:四尾栅藻Scenedesmusquadricauda,Tra:囊裸藻属Trachelomonas 图 6 浮游植物物种-环境因子冗余分析排序图 Fig. 6 Ordination map of redundancy analysis(RDA) of phytoplankton species-environmental factors |
综合营养状态指数(TLI)显示,本研究调查河道水体处于轻度富营养-中度富营养化状态,与浮游植物Shannon物种多样性指数(H′)及丰富度指数(R)评价的中污染状态基本吻合,但均匀度指数显示河道处于轻污染程度,说明浮游植物的均匀度指数不适合评价上海市环城绿带休憩型河道的水环境质量。微小平裂藻、尖尾蓝隐藻、啮噬隐藻和小球藻均可成为富营养化或污染较严重水体的优势种[12-14, 21, 29-30],颤藻属、伪鱼腥藻属、小环藻属以及直链藻属或者这些属的一些物种也能在富营养化或污染较严重水体中占优势[11-12, 29-32],以上这些种属均为本研究河道浮游植物群落的优势种属,这说明水质评价的指示物种法与TLI及H′和R评价结果较为吻合。
从不同类型河道来看,有沉水植物河道浮游植物的H′评价得到中污染状态与TLI评价得到的轻度富营养化比较一致,无沉水植物河道浮游植物的R评价得到的α-中污型污染程度与TLI评价的中度富营养化比较符合,说明适合使用H′评价上海市有水草河道的水体质量、使用R评价无沉水植物河道的污染程度。
通过比较发现,不同学者报道[5, 21]的上海市河道浮游植物优势种属、污染指示种不尽一致,一方面可能与调查的年份特别是季节不一样所导致,如李为星等[21]和姜雪芹等[5]分别是在2011年8月和2007—2008年冬季采样,而本文是季节调查;另一方面可能与城市河道是不稳定生态系统有关[5]。李为星等[21]曾提出尖尾蓝隐藻、啮蚀隐藻和梅尼小环藻等9个浮游植物物种为上海市城郊河道污染指示种,本文发现蓝隐藻、啮蚀隐藻和小环藻1种是有沉水植物河道的优势种、非无沉水植物河道的优势种,因此说明这3个物种为有沉水植物富营养化河道污染指示种。尽管在前两篇文献报道中伪鱼腥藻不是上海市富营养化河道的优势种,但本文结果显示该物种为无沉水植物富营养化河道的优势种,说明该物种也能作为上海市河道污染指示种。
3.2 沉水植物对河道浮游植物群落结构的影响沉水植物可以通过化感效应、遮蔽光照、降低水体氮磷浓度、降低蓝藻对其它浮游植物物种的竞争优势,以及为浮游动物提供庇护所而增加对浮游植物的摄食压力等直接或间接作用,改变浮游植物的群落结构如物种组成、密度、生物量以及物种多样性[33-35]。本研究表明,有沉水植物河道的浮游植物群落(WSM)密度和生物量均显著低于无沉水植物河道(NWSM),RDA分析也显示大多数浮游植物物种密度与水草负相关(图 5),说明水草能显著抑制浮游植物的生长和繁殖[34-36]。
水草存在与否显著影响浮游植物群落的物种组成特别是优势种组成,大量研究表明,水草修复前后,浮游植物群落物种组成会发生较大的变化,优势种一般从蓝藻门和绿藻门物种为主转变为以隐藻门、硅藻门和/或绿藻门物种为主[31, 37-40],或蓝藻门物种如微囊藻占优势转变为绿藻门物种如小球藻占优势[35]。本研究结果与此类似,WSM河道的物种组成与NWSM河道存在显著差异,WSM类型河道优势种包括隐藻门的尖尾蓝隐藻和啮噬隐藻、绿藻门的小球藻和硅藻门的小环藻;NWSM类型河道优势种包括蓝藻门的微小平裂藻、颤藻和伪鱼腥藻,绿藻门的小球藻和硅藻门的直链藻,其中微小平裂藻、颤藻和尖尾蓝隐藻为两种河道类型的主要分歧种,微小平裂藻和颤藻为NWSM类型河道的主要特征种,尖尾蓝隐藻为WSM类型河道的主要特征种。平裂藻、颤藻等蓝藻是小型藻类,个体细胞微小,单位体重表面积大,对光照和营养盐等资源的响应时间短[38],这可能导致了本文调查的无水草且富营养化程度更高河道中的微小平裂藻和颤藻比其它浮游植物物种更具竞争优势,特别是在水温较高的季节能快速增长从而导致蓝藻水华,夏季这两种藻类的密度达58.8×106 cell/L,占该季节藻类总密度的77.5%,年均密度也高达全年总密度的60.5%。
隐藻门物种个体较小、沉降速率低、繁殖能力高、生长速率快、对光照的要求比较低、且其能在水层中迁移,获取营养物质的能力较强,这些特征使该门中的一些代表性物种如尖尾蓝隐藻在某些特定季节和环境条件下具有竞争优势。在光照强度比较低的冬春季和秋冬季,尖尾蓝隐藻分别为武汉月湖[37]和安庆新洲水域[17]浮游植物群落的优势种,甚至能成为一些富营养化湖泊如上海淀山湖和滴水湖4个季节的优势种[13-14]。大型水生植被的重建,消耗了水体中氮磷等营养物质,同时抑制了浮游植物的光合作用,尖尾蓝隐藻成为我国水生植物重建湖泊或种植水体如惠州西湖、武汉月湖及上海青草沙水库实验池的优势种[31, 37-39]。本文RDA显示,尖尾蓝隐藻密度与沉水植物有一定的正相关关系(图 5),说明沉水植物的存在有利于它的生长,其占了沉水植物河道(WSM)浮游植物年均密度的34.9%,秋季和冬季比例更高,分别占了57.6%和38.3%。
不少大型水生植物修复湖泊和种植实验小水体发现,水生植物成功种植或重建后浮游植物的Shannon物种多样性(H′)、丰富度指数(R)和均匀度指数(J)显著或明显高于种植或重建前[36, 39-40],或H′显著高于种植或重建前[38],或大型水生植物修复示范区R高于未修复区[31],但也有报道表明,仅H′存在显著差异,而R无显著变化[35]。与这些研究不同的是,本研究中WSM类型河道的R显著低于NWSM类型河道,与滇池藻类生态系统浮游细菌物种丰富度显著高于草型生态系统[41]类似。这可能与以下原因有关,一方面有水草河道的沉水植物种植时间短、生态系统不稳定,适合在草型系统中的浮游植物物种还没有发展起来,但适合在藻型水体中生长生存的物种消失的比较快;另一方面,本研究中有水草河道是小型河道,采样点周围水域沉水植物覆盖度大,生境较为单一,而上述文献研究者是在较大型水体如整个湖泊或湖泊的某块区域中种植水草,而在其他区域存在未种植水草的区域,生境异质性较高,从而导致上述文献所报道的有水草区域的R高于或显著高于无水草区域。
3.3 其他理化因子对浮游植物群落结构的影响一般而言,生物包括浮游植物群落现存量(总密度和生物量)随营养盐浓度的变化呈单峰分布,即开始随着营养盐浓度的增加而增加,当营养盐浓度达到某一阈值浮游植物现存量最高,之后逐渐下降[42];但不同的物种对营养盐浓度的响应阈值不一致,污染敏感物种的营养盐阈值较低、耐污染物种的较高。BOD5、CODMn、NH3-N、TN和TP能指示水体营养盐浓度,是表征水体污染程度的重要环境因子。很多研究表明,浮游植物群落总密度和/或总生物量与以上一种或者多种环境因子显著正相关[35, 40, 43-45];一些采用多元统计分析的研究报道,浮游植物物种的时空分布受到多种环境因子如BOD5及沉水植物等环境因子的影响[46-47]。与此一致,本文研究河道的浮游植物总密度和生物量也与BOD5、CODMn、NH3-N、TN和TP显著正相关,浮游植物主要物种均与BOD5呈正相关关系,表明这些物种的污染耐受能力较强,吉林查干湖丰水期浮游植物优势属也倾向于生活在高磷素、高BOD5的生境中[46]。
| [1] |
ROUSSEAUX C S, GREGG W W. Interannual variation in phytoplankton primary production at a global scale[J]. Remote Sensing, 2014, 6(1): 1-19. |
| [2] |
CHEW L L, CHONG V C, TANAKA K, et al. Phytoplank-ton fuel the energy flow from zooplankton to small nekton in turbid mangrove waters[J]. Marine Ecology Progress Series, 2012, 469: 7-24. DOI:10.3354/meps09997 |
| [3] |
ZHANG P Y, TANG H J, GONG Z J, et al. Phytoplankton abundance constrains planktonic energy subsidy to benthic food web[J]. Journal of Ecosystem & Ecography, 2014, 4(1): 1-5. |
| [4] |
SUIKKANEN S, LAAMANEN M, HUTTUNEN M. Long-term changes in summer phytoplankton communities of the open northern Baltic Sea[J]. Estuarine, Coastal and Shelf Science, 2007, 71(3/4): 580-592. |
| [5] |
姜雪芹, 禹娜, 毛开云, 等. 冬季上海市城区河道中浮游植物群落结构及水质的生物评价[J]. 华东师范大学学报(自然科学版), 2009(2): 78-87, 140. JIANG X Q, YU N, MAO K Y, et al. Assessment of algal diversity and water quality in the four urban rivers of Shanghai in winter[J]. Journal of East China Normal University (Natural Science), 2009(2): 78-87, 140. |
| [6] |
王俊莉, 刘冬燕, 古滨河, 等. 基于浮游植物群落的安徽太平湖水环境生态评价[J]. 湖泊科学, 2014, 26(6): 939-947. WANG J L, LIU D Y, GU B H, et al. Environmental assessment of Lake Taiping (Anhui Province) based on a phytoplankton community analysis[J]. Journal of Lake Sciences, 2014, 26(6): 939-947. |
| [7] |
王艺兵, 侯泽英, 叶碧碧, 等. 鄱阳湖浮游植物时空变化特征及影响因素分析[J]. 环境科学学报, 2015, 35(5): 1310-1317. WANG Y B, HOU Z Y, YE B B, et al. The characteristics of spatial and temporal variations of phytoplankton in Poyang Lake and their influencing factors[J]. Acta Scientiae Circumstantiae, 2015, 35(5): 1310-1317. |
| [8] |
朱为菊, 庞婉婷, 尤庆敏, 等. 淮河流域春季浮游植物群落结构特征及其水质评价[J]. 湖泊科学, 2017, 29(3): 637-645. ZHU W J, PANG W T, YOU Q M, et al. Phytoplankton community structure and the evaluation of water quality in spring, Huaihe River Basin[J]. Journal of Lake Sciences, 2017, 29(3): 637-645. |
| [9] |
孙玉平, 于恒国, 周钦, 等. 典型富营养化城市河流-浙江温瑞塘河的浮游植物群落类型与季节变化[J]. 湖泊科学, 2018, 30(2): 375-384. SUN Y P, YU H G, ZHOU Q, et al. Seasonal variation of phytoplankton communities in Wenruitang River-A typical eutrophic urban river, Zhejiang Province[J]. Journal of Lake Sciences, 2018, 30(2): 375-384. |
| [10] |
方慷, 刘存歧, 杨军. 大清河水系保定段城市河道浮游植物群落结构及水质评价[J]. 水生态学杂志, 2013, 34(3): 25-31. FANG K, LIU C Q, YANG J. Assessment on phytoplankton community structure and water quality of Baoding city watercourse in Daqinghe Basin[J]. Journal of Hydroecology, 2013, 34(3): 25-31. |
| [11] |
李秋华, 陈丽丽, 夏品华, 等. 贵州百花湖麦西河河口浮游植物群落结构及与环境因子关系[J]. 湖泊科学, 2011, 23(4): 612-618. LI Q H, CHEN L L, XIA P H, et al. Structure of phytoplankton community and its relationship with environmental factors at the estuary of Maixi River in Baihua Reservoir, Guizhou Province[J]. Journal of Lake Sciences, 2011, 23(4): 612-618. |
| [12] |
王丽卿, 施荣, 季高华, 等. 淀山湖浮游植物群落特征及其演替规律[J]. 生物多样性, 2011, 19(1): 48-56. WANG L Q, SHI R, JI G H, et al. Phytoplankton community structure and its succession in Dianshan Lake[J]. Biodiversity Science, 2011, 19(1): 48-56. |
| [13] |
童琰, 马明睿, 林青, 等. 滴水湖浮游植物时空分布动态及其影响因子[J]. 长江流域资源与环境, 2012, 21(12): 1499-1506. TONG Y, MA M R, LIN Q, et al. Dynamics of spatial and temporal distribution of phytoplankton and related affecting factors in the Dishui Lake[J]. Resources and Environment in the Yangtze Basin, 2012, 21(12): 1499-1506. |
| [14] |
徐春燕, 俞秋佳, 徐凤洁, 等. 淀山湖浮游植物优势种生态位[J]. 应用生态学报, 2012, 23(9): 2550-2558. XU C Y, YU Q J, XU F J, et al. Niche analysis of phytoplankton's dominant species in Dianshan Lake of East China[J]. Chinese Journal of Applied Ecology, 2012, 23(9): 2550-2558. |
| [15] |
胡忠军, 莫丹玫, 周小玉, 等. 千岛湖浮游植物群落结构时空分布及其与环境因子的关系[J]. 水生态学杂志, 2017, 38(5): 46-54. HU Z J, MO D M, ZHOU X Y, et al. Spatial and seasonal distribution of phytoplankton in Qiandao lake and relationship with environmental factors[J]. Journal of Hydroecology, 2017, 38(5): 46-54. |
| [16] |
严文逸, 王绍祥, 顾静, 等. 青草沙水库浮游藻类群落组成及其与环境因子的关系[J]. 上海海洋大学学报, 2017, 26(1): 75-84. YAN W Y, WANG S X, GU J, et al. Composition of planktonic algae community and its relationship with environmental factors in Qingcaosha Reservoir[J]. Journal of Shanghai Ocean University, 2017, 26(1): 75-84. |
| [17] |
张家路, 王银平, 蔺丹清, 等. 安庆新洲水域浮游植物群落结构特征[J]. 上海海洋大学学报, 2019, 28(5): 680-688. ZHANG J L, WANG Y P, LIN D Q, et al. Characteristics of community structure of phytoplankton of Xinzhou waters in Anqing[J]. Journal of Shanghai Ocean University, 2019, 28(5): 680-688. |
| [18] |
张家路, 王银平, 蔺丹清, 等. 扬中夹江浮游植物种群特征及其对环境因子的响应[J]. 上海海洋大学学报, 2019, 28(4): 607-615. ZHANG J L, WANG Y P, LIN D Q, et al. Phytoplankton community structure in Jiajiang in Yangzhou Estuary and its relation to environmental factors[J]. Journal of Shanghai Ocean university, 2019, 28(4): 607-615. |
| [19] |
张真, 胡忠军, 史先鹤, 等. 千岛湖浮游植物群落结构及功能群的划分[J]. 上海海洋大学学报, 2019, 28(1): 37-48. ZHANG Z, HU Z J, SHI X H, et al. Community structures and functional groups of phytoplankton in Qiandao Lake[J]. Journal of Shanghai Ocean university, 2019, 28(1): 37-48. |
| [20] |
万蕾, 朱伟, 操家顺, 等. 苏州重污染河道水体浮游植物消长规律初探[J]. 长江流域资源与环境, 2006, 15(2): 237-243. WANG L, ZHU W, CAO J S, et al. Growth of phytoplankton in polluted river in Suzhou city[J]. Resources and Environment in the Yangtze Basin, 2006, 15(2): 237-243. |
| [21] |
李为星, 徐玉萍, 王丽卿, 等. 上海市城郊河道夏季浮游植物群落结构及水质评价[J]. 上海海洋大学学报, 2013, 22(3): 357-363. LI W X, XU Y P, WANG L Q, et al. Assessment of phytoplankton community structure and water quality in the seven suburb rivers of Shanghai in summer[J]. Journal of Shanghai Ocean University, 2013, 22(3): 357-363. |
| [22] |
周振明, 陈朝述, 刘可慧, 等. 漓江桂林市区段夏季浮游植物群落特征与水质评价[J]. 生态环境学报, 2014, 23(4): 649-656. ZHOU Z M, CHEN C S, LIU K H, et al. Phytoplankton community and water quality in guilin city section of Lijiang River in Summer, China[J]. Ecology and Environmental Sciences, 2014, 23(4): 649-656. |
| [23] |
罗梅, 刘昔, 陈国梁, 等. 城市景观河道浮游植物与水质评价[J]. 环境工程学报, 2016, 10(12): 7380-7386. LUO M, LIU X, CHEN G L, et al. Phytoplankton and water quality evaluation in a riverway of urban landscape[J]. Chinese Journal of Environmental Engineering, 2016, 10(12): 7380-7386. |
| [24] |
马婕, 申利亚, 何培民, 等. 苏州城区河道浮游植物功能群演替特征及其对环境因子的响应[J]. 上海海洋大学学报. SHEN L Y, HE P M, et al. Succession characteristics and responses to environmental factors of phytoplankton functional groups in river courses in the urban area of Suzhou City[J]. Journal of Shanghai Ocean university. DOI:10.12024/JSOU.20191202886 |
| [25] |
祁舒展, 姜卫兵, 魏家星. 我国环城绿带的发展:问题、对策与展望[J]. 中国城市林业, 2017, 15(3): 11-15. QI S Z, JIANG W B, WEI J X. Ring road greenbelts in China:problems, countermeasures and prospects[J]. Journal of Chinese Urban Forestry, 2017, 15(3): 11-15. |
| [26] |
孙诗雨, 王云. 上海环城绿带百米林带植物群落特征调查[J]. 上海交通大学学报(农业科学版), 2018, 36(6): 8-14. SUN S Y, WANG Y. Investigation and study on the features of plant communities of Shanghai Green Belt[J]. Journal of Shanghai Jiaotong University(Agricultural Science), 2018, 36(6): 8-14. |
| [27] |
章宗涉, 黄祥飞. 淡水浮游生物研究方法[M]. 北京: 科学出版社, 1991. ZHANG Z S, HUANG X F. Freshwater plankton research methods[M]. Beijing: Science Press, 1991. |
| [28] |
李振国, 卢军, 王国祥, 等. 分光光度法测定浮游植物叶绿素a的比较研究[J]. 中国环境监测, 2006, 22(2): 21-23. LI Z G, LU J, WANG G X, et al. Comparison of measurement of phytoplankton chlorophyll-a concentration by spectrophotometry[J]. Environmental Monitoring in China, 2006, 22(2): 21-23. |
| [29] |
吴洁, 虞左明. 西湖浮游植物的演替及富营养化治理措施的生态效应[J]. 中国环境科学, 2001, 21(6): 540-544. WU J, YU Z M. The succession of phytoplankton and the ecological effects of eutrophication control measures in Hangzhou West Lake[J]. China Environmental Science, 2001, 21(6): 540-544. |
| [30] |
郭坤, 杨德国, 彭婷, 等. 湖北省长湖浮游植物优势种生态位分析[J]. 湖泊科学, 2016, 28(4): 825-834. GUO K, YANG D G, PENG T, et al. Ecological niche analysis of dominant species of phytoplankton in Lake Changhu, Hubei Province[J]. Journal of Lake Sciences, 2016, 28(4): 825-834. |
| [31] |
陈亮, 张修峰, 刘正文. 惠州西湖浮游植物群落对生态系统修复的响应[J]. 武汉植物学研究, 2010, 28(4): 453-459. CHEN L, ZHANG X F, LIU Z W. The response of a phytoplankton community to ecosystem restoration in Huizhou West Lake[J]. Journal of Wuhan Botanical Research, 2010, 28(4): 453-459. |
| [32] |
蓝于倩, 袁一文, 彭亮, 等. 江谷水库鱼类网箱养殖富营养化及浮游植物功能群的指示作用[J]. 生态环境学报, 2015, 24(6): 1028-1036. LAN Y Q, YUAN Y W, PENG L, et al. Cage cultural eutrophication and phytoplankton functional groups as Bio-indicators in Jianggu Reservoir (Zhaoqing City, Guangdong Province)[J]. Ecology and Environmental Sciences, 2015, 24(6): 1028-1036. |
| [33] |
高健, 周敏, 闵婷婷, 等. 惠州西湖生态修复对浮游植物功能类群的影响[J]. 生态科学, 2013, 32(5): 540-545, 563. GAO J, ZHOU M, MIN T T, et al. Response of the phytoplankton functional groups to ecological restoration in Huizhou Lake[J]. Ecological Science, 2013, 32(5): 540-545, 563. |
| [34] |
姚远, 贺锋, 胡胜华, 等. 沉水植物化感作用对西湖湿地浮游植物群落的影响[J]. 生态学报, 2016, 36(4): 971-978. YAO Y, HE F, HU S H, et al. Effects of allelopathy of submerged macrophytes on the phytoplankton community collected from the west part of the West Lake wetland in Hangzhou, China[J]. Acta Ecologica Sinica, 2016, 36(4): 971-978. |
| [35] |
ZENG L, HE F, ZHANG Y, et al. How submerged macrophyte restoration promotes a shift of phytoplankton community in a shallow subtropical lake[J]. Polish Journal of Environmental Studies, 2017, 26(3): 1363-1373. DOI:10.15244/pjoes/68228 |
| [36] |
张明珠, 余帆洋, 唐清华, 等. 白云湖水生态构建示范区浮游植物群落初步研究[J]. 环境科学与技术, 2015, 38(6): 88-92. ZHANG M Z, YU F Y, TANG Q H, et al. Preliminary study of phytoplankton in ecological construction demonstration area of Baiyun Lake[J]. Environmental Science & Technology, 2015, 38(6): 88-92. |
| [37] |
邓平, 马建敏, 吴晓辉, 等. 武汉月湖水生植被重建过程中浮游植物的动态变化[J]. 湖泊科学, 2007, 19(5): 552-557. DENG P, MA J M, WU X H, et al. Dynamics of phytoplankton in the process of the aquatic macrophyte rehabilitation in Lake Yuehu, Wuhan[J]. Journal of Lake Sciences, 2007, 19(5): 552-557. |
| [38] |
闵婷婷, 刘正文, 李传红. 惠州西湖生态修复对浮游植物的影响[J]. 生态环境学报, 2011, 20(4): 701-705. MIN T T, LIU Z W, LI C H. Comparative studies of phytoplankton communities in restored and un-restored areas in Huizhou West Lake[J]. Ecology and Environmental Sciences, 2011, 20(4): 701-705. |
| [39] |
刘耘彤, 何文辉, 张慧, 等. 立体复合生态操纵水体浮游植物群落结构特征及水质评价[J]. 上海海洋大学学报, 2013, 22(2): 253-259. LIU Y T, HE W H, ZHANG H, et al. Characteristics of phytoplankton community structure and evaluation of water quality of multi-restoration ecological waterbody[J]. Journal of Shanghai Ocean University, 2013, 22(2): 253-259. |
| [40] |
代志刚, 蔺庆伟, 易科浪, 等. 杭州西湖浮游植物群落对沉水植物恢复的响应[J]. 水生态学杂志, 2017, 38(5): 35-45. DAI Z G, LIN Q W, YI K L, et al. Response of phytoplankton community to restoration of submerged macrophytes in West Lake, Hangzhou[J]. Journal of Hydroecology, 2017, 38(5): 35-45. |
| [41] |
WANG Y J, LI H B, XING P, et al. Contrasting patterns of free-living bacterioplankton diversity in macrophyte-dominated versus phytoplankton blooming regimes in Dianchi Lake, a shallow lake in China[J]. Chinese Journal of Oceanology and Limnology, 2017, 35(2): 336-349. DOI:10.1007/s00343-016-5277-9 |
| [42] |
PEARSON T H, ROSENBERG R. Macrobenthic succession in relation to organic enrichment and pollution of the marine environment[J]. Oceanography and Marine Biology, An Annual Review, 1978, 16: 229-311. DOI:10.1111/j.1610-0387.2007.06252.x |
| [43] |
夏霆. 通江城市河道浮游植物分布研究——以镇江市古运河为例[J]. 长江流域资源与环境, 2010, 19(11): 1249-1255. XIA T. Phytoplankton distribution of an urban river connected with the Yangtze River——A case study of the ancient canal, Zhenjiang, China[J]. Resources and Environment in the Yangtze Basin, 2010, 19(11): 1249-1255. |
| [44] |
宋芳芳, 刘娅琴, 宋祥甫, 等. 浮游植物群落对不同沉水植物生境的响应[J]. 上海农业学报, 2014, 30(4): 34-41. SONG F F, LIU Y Q, SONG X F, et al. Responses of phytoplankton to different submerged plant habitats[J]. Acta Agriculturae Shanghai, 2014, 30(4): 34-41. |
| [45] |
郑灿, 段杰仁, 石伟, 等. 星海湖浮游植物群落结构及与水环境因子的关系[J]. 水产学杂志, 2020, 33(1): 46-52. ZHENG C, DUAN J R, SHI W, et al. Phytoplankton community structure and its relationship with water environmental factors in Xinghai Lake[J]. Chinese Journal of Fisheries, 2020, 33(1): 46-52. |
| [46] |
李然然, 章光新, 张蕾. 查干湖湿地浮游植物与环境因子关系的多元分析[J]. 生态学报, 2014, 34(10): 2663-2673. LI R R, ZHANG G X, ZHANG L. Multivariate analysis of the relations between phytoplankton assemblages and environmental factors in Chagan Lake Wetland[J]. Acta Ecologica Sinica, 2014, 34(10): 2663-2673. |
| [47] |
王振方, 张玮, 杨丽, 等. 异龙湖不同湖区浮游植物群落特征及其与环境因子的关系[J]. 环境科学, 2019, 40(5): 2249-2257. WANG Z F, ZHANG W, YANG L, et al. Characteristics of phytoplankton community and its relationship with environmental factors in different regions of Yilong Lake, Yunnan Province, China[J]. Environmental Science, 2019, 40(5): 2249-2257. |
2. Center for Fish Nutrition and Environmental Ecology, Department of Agriculture and Rural Affairs, Shanghai Ocean University, Shanghai 201306, China;
3. Shanghai Engineering Research Center of Aquaculture, Shanghai Ocean University, Shanghai 201306, China