 2021,
Vol. 30
2021,
Vol. 30



2. 农业农村部大洋渔业开发重点实验室, 上海 201306;
3. 国家远洋渔业工程技术研究中心, 上海 201306;
4. 大洋渔业资源可持续开发教育部重点实验室, 上海 201306;
5. 农业农村部大洋渔业资源环境科学观测实验站, 上海 201306
柔鱼(Ommastrephes bartramii)是大洋性头足类经济种类,广泛分布于西北太平洋海域,是我国鱿钓渔船的重要捕捞对象[1-2]。柔鱼是一种“生态机会主义”物种,其中心渔场分布与资源量大小极易受到海洋气候和环境条件的影响[3]。已有研究表明,柔鱼资源丰度的变化与环境因子有密切的关系,例如海表面温度(sea surface temperature,SST)[4]、叶绿素a(chlorophyll a,Chl.a)[5]质量浓度、海表面高度(sea surface height,SSH)[6]、海表面盐度(sea surface salinity,SSS)[7]等;此外,柔鱼资源丰度的变化还会受到厄尔尼诺和拉尼娜事件[8]的显著影响。厄尔尼诺和拉尼娜事件起源于太平洋中东部赤道海域出现的一股异常暖或冷的水团[9],其交替循环对北太平洋渔业尤其是短生命周期的头足类种群变化起着至关重要的作用[10]。已有研究[11]表明拉尼娜事件发生时,SST相对正常年份降低,西北太平洋海域柔鱼适宜栖息地面积增加,导致柔鱼产量变高,厄尔尼诺事件发生时则相反。所以,分析不同强度厄尔尼诺和拉尼娜事件对柔鱼资源丰度的影响,为柔鱼资源评估和科学管理提供技术支撑。
灰色系统理论是一门不确定系统理论学科,其优点在于允许样本数量较少且服从任意分布,这对于缺乏数据的渔业科学领域来说有很大的应用前景[12]。目前,在资源量预测[13-14]、渔场时空分布[15]、渔业水质评价[16]和CPUE标准化[17]等方面的研究已取得了较好的成果。本研究采用灰色系统中的灰色关联聚类对北太平洋柔鱼各年、各月资源丰度进行聚类,分析各类群特征差异以及资源丰度与气候、环境因子的关系,了解变化规律,为柔鱼资源的可持续利用提供科学依据。
1 材料与方法 1.1 渔业数据渔业生产数据来自上海海洋大学鱿钓技术组,时间为1998—2017年5—12月,空间范围为35°N~45°N和140°E~179°E,统计内容包括日期、经度、纬度、日产量和作业渔船,空间分辨率为1°×1°。分别计算各年和各月每个渔区的单位捕捞努力量渔获量(catch per unit effort,CPUE),其公式为
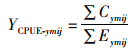 (1)
(1)
式中:YCPUE-ymij为某一渔区某年某月的单位捕捞努力量渔获量,t/(船·年)或t/(船·月); ∑Cymij为某一个经度或纬度所有渔区内某年某月的累计渔获量,t; ∑Eymij为某一个经度或纬度所有渔区内某年某月的累计捕捞努力量,船;y为年份;m为月份;i为经度;j为纬度。
1.2 环境数据反演的遥感环境数据包括海表面温度和叶绿素a质量浓度,来源于NOAA的Oceanwatch网站(http://oceanwatch.pifsc.noaa.gov/erddap/index.html),时间为1998—2017年,空间范围分为产卵场和育肥场2个海域,产卵场海域数据时间1—5月,范围为20°N~30°N和130°E~170°E[8];育肥场海域数据时间7—11月,范围为35°N~50°N和150°E~175°E[8]。时间分辨率为月,空间分辨率为1°×1°。同时计算产卵场与育肥场海域SST和Chl.a质量浓度的距平值(sea surface temperature anomaly,SSTA;chlorophyll a anomaly,CHLA),分析它们与CPUE的关系。
1.3 气候事件的定义根据美国NOAA气候预报中心(http://ggweather.com/enso/oni.htm)对厄尔尼诺和拉尼娜事件的定义,若Niño3.4区SSTA连续5个月滑动平均值超过+0.5 ℃认为发生1次厄尔尼诺事件;若连续5个月低于-0.5 ℃则认为发生一次拉尼娜事件。根据尼诺指数(Ocean Niño index, ONI)的大小可将厄尔尼诺和拉尼娜事件划分为不同强度(表 1),若ONI处于各临界值区间内连续超过3个月,则认为发生此类强度的异常气候事件。本研究根据此定义将1998—2017年发生的异常气候事件按照强度进行划分(表 2)。

|
表 1 厄尔尼诺和拉尼娜事件强度划分依据 Tab.1 Definition of El Niño and La Niña events intensity division |

|
表 2 1998—2017年不同强度厄尔尼诺和拉尼娜事件划分 Tab.2 Classification of the El Niño and La Niña events intensity from 1998 to 2017 |
利用灰色关联聚类模型对北太平洋柔鱼资源丰度进行聚类[18]。设有m个观测对象,每个对象观测n个特征数据,得到序列如下:
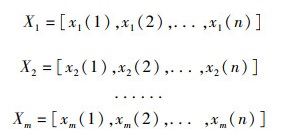 (2)
(2)
观测对象X为1998—2017年各个年份和5—12月各个月份的CPUE序列,特征数据x为某一经度或纬度范围内某年某月的CPUE平均值。
令所有i≤j,i,j=1、2、...、m,计算Xi与Xj的灰色绝对关联度εij[18],得到关联系数矩阵如下:
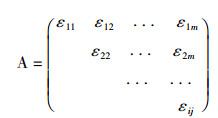 (3)
(3)
设临界值r(r>0.5),当εij≥r(i≠j)时,则视为Xj与Xi为同类特征。
2 结果 2.1 产量和CPUE变化1998—2017年产量和CPUE年间变化显著(图 1):20年的年平均产量为3.40万t,1998年产量最高为8.10万t,2012年产量最低为0.06万t;20年的年平均CPUE为2.26 t/(船·年),2007年CPUE最高为4.19 t/(船·年),2009年CPUE最低为1.32 t/(船·年),除2007年和2008年,年CPUE与年产量变化基本一致。

|
图 1 1998—2017年西北太平洋柔鱼年产量及CPUE变化 Fig. 1 Annual yield and CPUE of O. bartramii in the Northwest Pacific from 1998 to 2017 |
5—12月产量和CPUE变化呈现先增加后减少的正态分布趋势(图 2):8个月的月平均产量为8.20万t,8月产量最高为16.00万t,5月产量最低为0.40万t;8个月的月平均CPUE为1.31 t/(船·月),8月CPUE最高为2.06 t/(船·月),12月CPUE最低为0.68 t/(船·月)。

|
图 2 5—12月西北太平洋柔鱼月产量及CPUE变化 Fig. 2 Monthly yield and CPUE of O. bartramii in the Northwest Pacific from May to December |
从灰色关联聚类系数来看(表 3和表 4),年间与月间各经、纬度向CPUE关联度均大于0.51,年间CPUE各经、纬度向关联度平均值为0.64和0.67,月间CPUE各经、纬度向关联度平均值为0.61和0.65,表明年间与月间CPUE空间分布相似性都较大。由于灰色关联聚类中的临界值r可根据实际问题来确定,若r越接近于1,分类越细,每组变量越少,反之分类越粗,每组变量越多。所以取年间CPUE关联聚类矩阵临界值r=0.90,月间CPUE关联聚类矩阵临界值r=0.75作为分类依据。

|
表 3 1998—2017年各经、纬度向CPUE空间分布的灰色关联聚类系数 Tab.3 Grey relational clustering coefficients of CPUE spatial distribution at each longitude and latitude from 1998 to 2017 |

|
表 4 5—12月各经度向、纬度向CPUE空间分布的灰色关联聚类系数 Tab.4 Grey relational clustering coefficients of CPUE spatial distribution at each longitude and latitude from May to December |
根据灰色关联聚类系数矩阵得到各聚类类群(表 5)。从结果来看,年间纬度向类群分为明显的4类:类群1包含的对象数量最多,为8个年份,包括中强度、弱强度和正常年份气候事件;类群2包含的对象数量为2个年份,全部为强拉尼娜事件;类群3包含的对象数量为2个年份,为强拉尼娜和超强厄尔尼诺事件;类群4包含的对象数量为3个年份,全部为弱拉尼娜事件。其余年份自成一类,为弱强度和中强度气候事件,大多数为2种不同气候事件发生转换的年份。

|
表 5 1998—2017年各类群结果 Tab.5 Results of various groups from 1998 to 2017 |
年间经度向类群也分为明显的4类(表 5):类群1包含的对象数量最多,为6个年份,包括中、弱强度厄尔尼诺、弱强度拉尼娜和正常年份气候事件;类群2包含的对象数量为3个年份,为强拉尼娜和正常年份气候事件;类群3包含的对象数量为4个年份,为强、弱强度拉尼娜事件;类群4包含的对象数量为2个年份,为中强度厄尔尼诺和正常年份气候事件。其余年份自成一类,为中强度和超强度气候事件,大多数为2种不同气候事件发生转换的年份。
月间纬度向类群分为3类(表 6):类群1为5、6、7、9、10和11月;类群2为8月;类群3为12月。月间经度向类群分为4类,类群1为5、6、7和8月;类群2为9月;类群3为10月;类群4为11和12月。

|
表 6 5—12月各类群结果 Tab.6 Results of various groups from May to December |
对西北太平洋柔鱼产卵场和育肥场海域各年、月经度向和纬度向类群的SST与SSTA进行平均得到各类群CPUE与SST、SSTA的关系(图 3和图 4)。

|
图 3 1998—2017年各类群产卵场和育肥场平均海表温度距平的变化 Fig. 3 Variability of the averaged SSTA of various groups on the spawning ground and feeding ground from 1998 to 2017 |

|
图 4 5—12月各类群产卵场和育肥场平均海表温度的变化 Fig. 4 Variability of the averaged SST of various groups on the spawning ground and feeding ground from May to December |
由图 3可知,纬度向各类群的CPUE差异比经度向类群明显,各类群的产卵场海表面温度距平值(SGSSTA)和育肥场海表温度距平值(FGSSTA)变化基本上一致,且高CPUE类群对应高的SSTA,低CPUE类群对应低的SSTA,即年CPUE与SSTA呈正相关关系。类群2的CPUE最高,包含2个强拉尼娜年份,类群3的CPUE最低,包含1个强拉尼娜和1个超强厄尔尼诺年份,说明强拉尼娜事件会使海表温度降低,导致CPUE增高;超强厄尔尼诺事件会使海表温度升高,导致CPUE骤减。类群1和类群4均为中、弱强度气候事件和正常气候年份,CPUE接近年平均CPUE,海表面温度距平值为-0.10~0.10,说明弱拉尼娜事件和中强度气候事件对年CPUE的影响不大。经度向各类群CPUE均小于年平均CPUE,类群4的CPUE最低,包含1个中强度厄尔尼诺年份和1个正常气候年份。
由图 4可知,经度向类群和纬度向类群的CPUE均有明显差异。各类群的产卵场海表面温度(SGSST)和育肥场海表面温度(FGSST)变化一致,且SGSST大于FGSST,月CPUE与SST呈正相关关系。其中,纬度向类群2和经度向类群2的CPUE最高,分别为8月和9月,纬度向类群3和经度向类群4的CPUE最低,为11月和12月,相比于纬度向类群,经度向各类群季节变化特征更为明显。
2.4.2 产卵场与育肥场平均Chl.a和CHLA由图 5可知,纬度向各类群的CPUE差异比经度向类群明显,各类群的产卵场叶绿素a质量浓度距平值(SGCA)和育肥场叶绿素a质量浓度距平值(FGCA)变化基本上一致,且FGCA变化幅度比SGCA小。高CPUE类群对应低CHLA,而低CPUE类群对应高CHLA,即年CPUE与CHLA呈负相关关系,恰好与SSTA的关系相反。类群2的CPUE最高,包含2个强拉尼娜年份,类群3的CPUE最低,包含1个强拉尼娜和1个超强厄尔尼诺年份,说明强拉尼娜事件会使Chl.a质量浓度升高,导致CPUE增高;超强厄尔尼诺事件会使Chl.a质量浓度降低,导致CPUE骤减。经度向各类群CPUE均小于年平均CPUE,CHLA变化幅度小,SGCA与FGCA数值为-0.01~0.01。

|
图 5 1998—2017年各类群产卵场和育肥场平均叶绿素a质量浓度距平的变化 Fig. 5 Variability of the averaged Chl.a concentration anomaly of various groups on the spawning ground and feeding ground from 1998 to 2017 |
由图 6可知,经度向类群和纬度向类群的CPUE均有明显差异。各类群的产卵场叶绿素a质量浓度(SGC)和育肥场叶绿素a质量浓度(FGC)变化一致,FGC大于SGC且FGC波动幅度更大。CPUE最高的纬度向类群2和经度向类群2的Chl.a质量浓度均最低,纬度向类群3和经度向类群4的CPUE最低,SGC最高,纬度向类群1和经度向类群1的FGC最高,各类群有明显的季节性变化,即SGC和FGC先递减再递增,8月和9月Chl.a质量浓度最低。

|
图 6 5—12月各类群产卵场和育肥场平均叶绿素a质量浓度的变化 Fig. 6 Variability of the averaged Chl.a mass concentration of various groups on the spawning ground and feeding ground from May to December |
柔鱼作为短生命周期的经济头足类,生活史通常为一年,这就极大依赖于环境条件的变化[19]。不同强度的厄尔尼诺拉尼娜事件对柔鱼渔场环境的调控机制不同,导致对柔鱼资源丰度产生不同的影响。如YU等[20]认为:弱拉尼娜事件和正常气候年份,柔鱼渔场内水温最高,适宜栖息地面积骤增;中高强度拉尼娜事件时,柔鱼渔场内水温较高,但适宜栖息地面积骤减;厄尔尼诺事件时,柔鱼渔场内水温低,弱强度和超高强度厄尔尼诺事件时,适宜栖息地面积大于中强度厄尔尼诺事件。本研究选取1998—2017年5—12月的渔获数据,基于不同气候事件会影响柔鱼年、月平均资源丰度的假设,从而导致不同年份、月份资源丰度的空间分布存在差异,利用灰色关联聚类的方法计算它们之间的关联度,根据关联度大小进行分类,分析各类别CPUE与气候事件和环境因子的关系。研究结果(表 5)发现,年纬度向类群比年经度向类群的CPUE差异更明显,强拉尼娜和弱拉尼娜事件集中的类群显著,厄尔尼诺、中强度拉尼娜和正常年份分类较模糊。例如:包含2个强拉尼娜事件年份的纬度向类群2的CPUE最高,产卵场和育肥场的SSTA也最大,但CHLA最低;包含3个弱拉尼娜事件的纬度向类群4和包含弱、强拉尼娜事件的经度向类群3的CPUE均略低于年平均CPUE,其SSTA较大,CHLA接近于0(图 3和图 5)。这说明拉尼娜事件的强弱对柔鱼资源的调控机制不同,拉尼娜事件会使渔场温度升高,更有利于柔鱼的生长与发育,从而提高产量,但强拉尼娜事件会使渔场Chl.a质量浓度下降,柔鱼资源丰度骤增。纬度向类群3和经度向类群4的CPUE均较低,分别包含1个超强厄尔尼诺事件和1个中强度厄尔尼诺事件,纬度向类群3的产卵场和育肥场的SSTA最小,但CHLA最高,表明中强度以上的厄尔尼诺事件会使柔鱼资源丰度骤减,这一结果与余为等[21]的研究结果有所出入,可能与Chl.a质量浓度遥感数据的来源、空间分辨率和卫星模拟算法的不同有关。从5—12月各类群结果(表 6)来看:各月份的CPUE与环境因子呈现明显的季节性变化,且经度向类群的季节性变化更显著,即SST夏季高冬季低,SGSST和FGSST呈先增加后减小的趋势,8、9月最高,11、12月最低,与CPUE变化一致;Chl.a质量浓度则恰好相反,5—7月最高,8、9月最低(图 4和图 6)。这些与已有研究结果[22-23]一致。
灰色关联聚类的原理是通过比较序列在直角坐标轴中形成的曲线图形的相似度来衡量其变量之间的相互关联性程度,拟合程度越好,关联度越大。相比于传统的统计学方法,允许服从任意分布,样本数量少,且计算方法简单,效果明显[24]。本文选取1998—2017年5—12月各经纬度的CPUE作为分析序列,从聚类矩阵结果(表 3和表 4)来看,关联度差异较大,能够较明显地对观测对象进行聚类,特征数据即各年份、月份经纬度向的CPUE差异明显,可以作为聚类分析的指标之一。但本研究仍存在一些不足,如虽然在不同强度的异常气候事件下的得到较好的聚类结果,但并未解释其对不同年份、月份聚类类群CPUE在空间上经纬度变化的内在机理。此外,本研究只分析了SST和Chl.a质量浓度对不同类群CPUE的影响,建议在今后的研究中加入其他环境因子,如海表面高度(SSH)、海表面盐度(SSS)、海流等因素,建立具体的量化模型,更好地对西北太平洋柔鱼资源丰度进行评价。
| [1] |
陈新军. 世界头足类资源开发现状及我国远洋鱿钓渔业发展对策[J]. 上海海洋大学学报, 2019, 28(3): 321-330. CHEN X J. Development status of world cephalopod fisheries and suggestions for squid jigging fishery in China[J]. Journal of Shanghai Ocean University, 2019, 28(3): 321-330. |
| [2] |
陈新军, 刘必林, 王尧耕. 世界头足类[M]. 北京: 海洋出版社, 2009: 60-69. CHEN X J, LIU B L, WANG Y G. Cephalopods of the world[M]. Beijing: China Ocean Press, 2009: 60-69. |
| [3] |
NISHIKAWA H, TOYODA T, MASUDA S, et al. Wind-induced stock variation of the neon flying squid (Ommastrephes bartramii) winter-spring cohort in the subtropical North Pacific Ocean[J]. Fisheries Oceanography, 2015, 24(3): 229-241. DOI:10.1111/fog.12106 |
| [4] |
XU J, CHEN X J, CHEN Y, et al. The effect of sea surface temperature increase on the potential habitat of Ommastrephes bartramii in the Northwest Pacific Ocean[J]. Acta Oceanologica Sinica, 2016, 35(2): 109-116. DOI:10.1007/s13131-015-0782-9 |
| [5] |
唐峰华, 樊伟, 伍玉梅, 等. 北太平洋柔鱼渔场资源与海洋环境关系的季节性变化[J]. 农业资源与环境学报, 2015, 32(3): 242-249. TANG F H, FAN W, WU Y M, et al. Seasonal changes of relationship between marine environment and squid fishing resources in north Pacific Ocean[J]. Journal of Agricultural Resources and Environment, 2015, 32(3): 242-249. |
| [6] |
宋婷婷. 基于海面高度数据研究西北太平洋巴特柔鱼(Ommastrephes bartrami)渔场分布[D]. 上海: 上海海洋大学, 2014. SONG T T. Relationship between fishing grounds of Ommastrephes bartrami and satellite altimeter data in northwestern Pacific[D]. Shanghai: Shanghai Ocean University, 2014. |
| [7] |
余为, 陈新军, 易倩, 等. 西北太平洋柔鱼传统作业渔场资源丰度年间差异及其影响因子[J]. 海洋渔业, 2013, 35(4): 373-381. YU W, CHEN X J, YI Q, et al. Annual difference of abundance index and its influencing factors of Ommastrephes bartramii in traditional fishing grounds in the northwest Pacific[J]. Marine Fisheries, 2013, 35(4): 373-381. DOI:10.3969/j.issn.1004-2490.2013.04.001 |
| [8] |
CHEN X J, ZHAO X H, CHEN Y. Influence of El Niño/La Niña on the western winter-spring cohort of neon flying squid (Ommastrephes bartramii) in the northwestern Pacific Ocean[J]. ICES Journal of Marine Science, 2007, 64(6): 1152-1160. DOI:10.1093/icesjms/fsm103 |
| [9] |
WANG C Z, DESER C, YU J Y, et al. El Niño and southern Oscillation (ENSO): a review[M]//GLYNN P, MANZELLO D, ENOCHS I. Coral Reefs of the Eastern Tropical Pacific. Dordrecht: Springer, 2017: 85-106.
|
| [10] |
ANDERSON C I H, RODHOUSE P G. Life cycles, oceanography and variability: ommastrephid squid in variable oceanographic environments[J]. Fisheries Research, 2001, 54(1): 133-143. DOI:10.1016/S0165-7836(01)00378-2 |
| [11] |
温健, 陆鑫一, 陈新军, 等. 基于海表温度和光合有效辐射的西北太平洋柔鱼冬春生群体栖息地热点预测[J]. 上海海洋大学学报, 2019, 28(3): 456-463. WEN J, LU X Y, CHEN X J, et al. Predicting the habitat hot spots of winter-spring cohort of Ommastrephes bartramii in the northwest Pacific Ocean based on the sea surface temperature and photosynthetically active radiation[J]. Journal of Shanghai Ocean University, 2019, 28(3): 456-463. |
| [12] |
陈新军. 灰色系统理论在渔业科学中的应用[M]. 北京: 中国农业出版社, 2003: 1-32. CHEN X J. Grey system theory in fisheries science[M]. Beijing: China Agriculture Press, 2003: 1-32. |
| [13] |
高雪, 陈新军, 余为. 基于灰色系统的西北太平洋柔鱼冬春生群资源丰度预测模型[J]. 海洋学报, 2017, 39(6): 55-61. GAO X, CHEN X J, YU W. Forecasting model of the abundance index of winter-spring cohort of Neon flying squid (Ommastrephes bartramii) in the northwest Pacific Ocean based on grey system theory[J]. Acta Oceanologica Sinica, 2017, 39(6): 55-61. DOI:10.3969/j.issn.0253-4193.2017.06.006 |
| [14] |
解明阳, 陈新军, 汪金涛. 基于灰色系统的太平洋褶柔鱼冬春生群资源丰度变化研究[J]. 海洋渔业, 2018, 40(6): 641-648. XIE M Y, CHEN X J, WANG J T. On changes of abundance index of winter-spring cohort of Todarodes pacificus in the northwest Pacific based on grey system theory[J]. Marine Fisheries, 2018, 40(6): 641-648. DOI:10.3969/j.issn.1004-2490.2018.06.001 |
| [15] |
李媛洁, 陈新军, 汪金涛, 等. 东南太平洋智利竹䇲鱼资源渔场时空分布[J]. 上海海洋大学学报, 2019, 28(4): 616-625. LI Y J, CHEN X J, WANG J T, et al. Spatio-temporal distribution of mackerel in southeast Pacific Ocean[J]. Journal of Shanghai Ocean University, 2019, 28(4): 616-625. |
| [16] |
赵龙明, 卢琪, 王娜, 等. 改进的灰色关联分析法在渔业水质评价中的应用[J]. 华北理工大学学报(自然科学版), 2017, 39(2): 110-114, 119. ZHAO L M, LU Q, WANG N, et al. Application of improved grey correlation analysis method in fishery water quality assessment[J]. Journal of North China University of Science and Technology (Natural Science Edition), 2017, 39(2): 110-114, 119. DOI:10.3969/j.issn.1674-0262.2017.02.016 |
| [17] |
杨胜龙, 张禹, 戴阳, 等. 基于GM(1, 1)模型的贫信息渔业数据CPUE标准化研究[J]. 应用海洋学学报, 2019, 38(1): 141-147. YANG S L, ZHANG Y, DAI Y, et al. CPUE standardization for poor information fishery data based on GM(1, 1) model[J]. Journal of Applied Oceanography, 2019, 38(1): 141-147. DOI:10.3969/J.ISSN.2095-4972.2019.01.016 |
| [18] |
刘思峰, 杨英杰, 吴利丰, 等. 灰色系统理论及其应用[M]. 7版. 北京: 科学出版社, 2014: 113-116. LIU S F, YANG Y J, WU L F, et al. Grey system theory and its application[M]. 7th ed. Beijing: Science Press, 2014: 113-116. |
| [19] |
YU W, CHEN X J, YI Q, et al. A review of interaction between neon flying squid (Ommastrephes bartramii) and oceanographic variability in the north Pacific Ocean[J]. Journal of Ocean University of China, 2015, 14(4): 739-748. DOI:10.1007/s11802-015-2562-8 |
| [20] |
YU W, CHEN X J, YI Q, et al. Variability of suitable habitat of western winter-spring cohort for neon flying squid in the Northwest Pacific under anomalous environments[J]. PLoS One, 2015, 10(4): e0122997. DOI:10.1371/journal.pone.0122997 |
| [21] |
余为, 陈新军. 西北太平洋柔鱼冬春生群体栖息地的变化研究[J]. 海洋学报, 2018, 40(3): 86-94. YU W, CHEN X J. Variability in the potential habitat of winter-spring cohort of neon flying squid Ommastrephes bartramii in the northwest Pacific Ocean[J]. Acta Oceanologica Sinica, 2018, 40(3): 86-94. DOI:10.3969/j.issn.0253-4193.2018.03.008 |
| [22] |
毛志华, 朱乾坤, 龚芳. 卫星遥感北太平洋渔场叶绿素a浓度[J]. 水产学报, 2005, 29(2): 270-274. MAO Z H, ZHU Q K, GONG F. Satellite remote sensing of chlorophyll a concentration in the north Pacific fishery[J]. Journal of Fisheries of China, 2005, 29(2): 270-274. |
| [23] |
余为, 陈新军. 西北太平洋柔鱼栖息地环境因子分析及其对资源丰度的影响[J]. 生态学报, 2015, 35(15): 5032-5039. YU W, CHEN X J. Analysis of environmental conditions and their influence on the abundance of neon flying squid in the northwest Pacific Ocean[J]. Acta Ecologica Sinica, 2015, 35(15): 5032-5039. |
| [24] |
刘勇, 王冬冬, 周婷. 基于决策粗糙集的多属性灰色关联聚类方法[J]. 控制与决策, 2017, 32(11): 2034-2038. LIU Y, WANG D D, ZHOU T. Decision-theoretic rough set based multi-attribute grey incidence clustering method[J]. Control and Decision, 2017, 32(11): 2034-2038. |
2. Key Laboratory of Oceanic Fisheries Exploration, Ministry of Agriculture and Rural Affairs, Shanghai 201306, China;
3. National Engineering Research Center for Oceanic Fisheries, Shanghai Ocean University, Shanghai 201306, China;
4. Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources, Ministry of Education, Shanghai Ocean University, Shanghai 201306, China;
5. Scientific Observing and Experimental Station of Oceanic Fishery Resources, Ministry of Agriculture and Rural Affairs, Shanghai 201306, China