 2020,
Vol. 29
2020,
Vol. 29



2. 大洋渔业资源可持续开发教育部重点实验室, 上海 201306;
3. 国家远洋渔业工程技术研究中心, 上海 201306;
4. 农业农村部大洋渔业开发重点实验室, 上海 201306;
5. 农业农村部大洋渔业资源环境科学观测实验站, 上海 201306
剑尖枪乌贼(Uroteuthis edulis)为近海暖温性种类[1],广泛分布于太平洋西北部及非洲东部沿岸海域[2-4],其中以我国东海资源密度最为丰富[5-7]。我国于1995年开始剑尖枪乌贼的商业性捕捞,随着光诱鱿钓技术的应用与推广,生产效率和产量逐渐提高[5-6]。剑尖枪乌贼的生命周期约为1年[8],世代更新快[9],摄食强度在繁殖期间达到高峰,随后逐渐减弱[10],依据产卵特性,将其划分为春、夏、秋、冬等4个产卵种群[1, 11-12]。已有学者对其渔业生物学[11-14]、渔场及渔获量时空分布[15-17]、资源评估与管理[18-19]等方面进行了研究,而目前研究的海域主要集中在东海南部和日本海海域,对于其主要索饵场即东海北部海域研究较少。了解渔业生物学特征是研究渔业物种资源量的基础工作之一。为此,本研究根据东海北部外海海域采样获得的剑尖枪乌贼样本对其渔业生物学特征进行分析,通过不同年份所采集的样品,比较其生物学年间差异,为全面了解剑尖枪乌贼的渔业生物学特征以及资源持续开发利用提供基础信息和科学依据。
1 材料与方法 1.1 样本来源采样时间分别为2017和2018年的9月至翌年2月,两年采样海域均为123°E~127°E,28°N~31.5°N,委托“浙岭渔23860号”拖网渔船对研究海域随机采样,2017年共采样245尾,2018年为368尾。样本经冷冻保存运回实验室进行生物学测定。
1.2 研究方法 1.2.1 生物学测定测定样本胴长、体质量、性别及性腺成熟度。使用皮尺测量样本胴长,精确到1 mm,电子秤称量体质量,精确到0.1 g。根据样本性腺发育程度将其成熟度划分为Ⅰ、Ⅱ、Ⅲ、Ⅳ和Ⅴ期[20],并判定性腺成熟度达到Ⅲ期及以上为性成熟期。
1.2.2 数据处理与分析采用频度分析法分析样本胴长及体质量分布,组距分别为20 mm和40 g。
采用线性回归法,分析年间雌雄个体的体质量-胴长关系,同时利用协方差分析(ANCOVA)年间个体体质量-胴长关系的差异,计算公式如下:
 (1)
(1)
式中:W为样本体质量,g;L为样本胴长,mm; a, b为估算的参数。
采用方差分析法(ANOVA),比较分析不同胴长组下性腺成熟度组成的差异。
采用Logistic曲线,估算年间雌雄个体的初次性成熟胴长,计算公式[21]如下:
 (2)
(2)
式中:Pi为给定胴长组下性成熟个体的占比;Li为给定胴长组的胴长;c, d为估算的参数;初次性成熟胴长L50%=-c/d。
对年间雌雄个体的胴长和体质量进行生长速度分析。采用绝对生长速率(absolute growth rate, AGR)和瞬时相对生长速率(instantaneous relative growth rate, G)分析剑尖枪乌贼的生长变化,计算公式[22]如下:
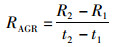 (3)
(3)
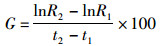 (4)
(4)
式中:R2为t2时刻样本的平均胴长或体质量;R1为t1时刻样本的平均胴长或体质量;t1、t2为采样时间;RAGR为绝对生长速率,单位为mm/d或g/d,G单位为%/d;
图表运用Microsoft Excel 2010软件绘制,统计分析运用SPSS Statistics 25软件完成。
2 结果与分析 2.1 胴长、体质量组成2017年渔汛期雄性个体胴长范围为69~234 mm,优势胴长组为100~180 mm,占整体样本的78.38%;2018年渔汛期分别为54~227 mm、80~160 mm和70.71%,两年渔汛期间雄性个体胴长差异极显著(P < 0.01)。2017年渔汛期雌性个体胴长范围为66~256 mm,优势胴长组为80~160 mm,占整体样本的72.39%;2018年渔汛期分别为61~223 mm、80~160 mm和79.41%,两年渔汛期间雌性个体胴长差异极显著(P < 0.01), 见图 1。两年中,雌雄个体间胴长均无显著性差异(P>0.05),并且2018年渔汛期雌雄个体的胴长组成均小于2017年。

|
图 1 剑尖枪乌贼胴长与体质量组成分布 Fig. 1 Distribution of mantle length and body mass of swordtip squid |
2017年渔汛期雄性个体体质量范围为17~356 g,优势体质量组为40~160 g,占整体样本的72.07%;2018年渔汛期分别为12~350 g、0~120 g和74.75%,两年渔汛期间雄性个体体质量差异极显著(P < 0.01)。2017年渔汛期雌性个体体质量范围为15~450 g,优势体质量组为40~160 g,占整体样本的67.16%;2018年渔汛期分别为13~354 g、0~120 g和75.29%。两年渔汛期间雌性个体体质量差异极显著(P < 0.01),见图 1。两年中,雌雄个体间体质量均无显著差异性(P>0.05),0~80 g个体数量逐渐增加,并且2018年渔汛期雌雄个体的体质量组成均小于2017年。综上可知,剑尖枪乌贼渔汛期雌雄个体的胴长和体质量年间存在差异。
2.2 体质量-胴长关系经协方差检验发现,两年渔汛期间雄性个体体质量-胴长关系差异显著(P < 0.05),雌性个体差异不显著(P>0.05),2017年渔汛期雌雄个体间体质量-胴长关系差异显著(P < 0.05),而2018年渔汛期雌雄个体间差异不显著(P>0.05)。2017和2018年渔汛期雌雄个体体质量-胴长关系拟合结果见图 2。

|
图 2 剑尖枪乌贼体质量与胴长关系 Fig. 2 Relationships between mantle length and body mass of swordtip squid |
雄性:
2017年:W=0.000 9L2.365 7 R2=0.94,P < 0.05,n=111;
2018年:W=0.001 1L2.322 8 R2=0.96,P < 0.05,n=198。
雌性:
2017年:W=0.001 0L2.346 0 R2=0.92,P < 0.05,n=134;
2018年:W=0.000 4L2.528 3 R2=0.95,P < 0.05,n=170。
2.3 性腺成熟度组成不同胴长组性腺成熟度比较(图 3)可知:在雄性个体中,胴长大于60 mm时出现性成熟的个体,2017年渔汛期性成熟个体的胴长主要集中在大于120 mm的个体,而2018年渔汛期则集中在大于100 mm的个体,并且在较小胴长组中高性腺成熟度的个体出现频率增加;在雌性个体中,胴长大于80 mm时出现性成熟的个体,2017年渔汛期性成熟个体的胴长主要集中在大于140 mm的个体,而2018年渔汛期则集中在大于100 mm的个体,并且在较小胴长组中高性腺成熟度的个体出现频率增加。因此,渔汛期雌雄个体的性腺成熟度组成和性成熟胴长年间均存在差异。

|
图 3 剑尖枪乌贼胴长与性腺成熟度关系 Fig. 3 Relationships between sexual maturity and mantle length of swordtip squid |
根据胴长与性腺成熟度关系,分别拟合2017和2018年渔汛期雌雄个体初次性成熟胴长的关系式(图 4):

|
图 4 剑尖枪乌贼初次性成熟胴长分布 Fig. 4 Distribution of mantle length at 50% maturity of swordtip squid |
2017年渔汛期雄性个体Pi= 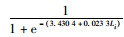
2017年渔汛期雌性个体Pi= 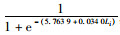
2018年渔汛期雄性个体Pi= 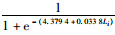
2018年渔汛期雌性个体Pi= 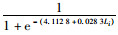
因此,初次性成熟胴长:2017年渔汛期雄性为147.19 mm,雌性为169.58 mm;2018年渔汛期雄性为129.58 mm,雌性为145.29 mm。
2.5 生长速率雄性个体胴长绝对生长率的变化范围为0.20~0.25 mm/d,相对生长率为0.16~0.17 %/d,雌性个体胴长生长率变化范围分别为0.34~0.40 mm/d和0.24~0.32 %/d。雄性个体体质量绝对生长率的变化范围为0.40~0.49 g/d,相对生长率为0.31~0.52 %/d,雌性个体体质量生长率变化范围分别为0.43~0.82 g/d和0.33~0.85 %/d,见表 1。因此,渔汛期雌性个体的胴长和体质量生长速率均快于雄性,并且雌雄个体的体质量生长速率年间差异较大,胴长生长速率年间相近。

|
表 1 剑尖枪乌贼胴长、体质量平均生长速率 Tab.1 Average growth rate of mantle length (ML) and body mass (BM) of swordtip squid |
2000年在东海南部海域通过灯光围网采样的研究表明,剑尖枪乌贼的优势胴长为81~120 mm,优势体质量为31~70 g[11, 14],2000—2002年在南海北部海域通过拖网采样的研究表明其优势胴长和体质量分别为40~110 mm、10~80 g[23],而本研究中雌雄个体的优势胴长和体质量均大于以上研究的结果。自东海海域实施伏季休渔制度[24]后,剑尖枪乌贼的渔汛主要集中在秋季[5],本研究采样时期为秋冬季,个体经过休渔后体型相对丰满,并且北部外海海域多为拖网渔业,而南部海域多为灯诱渔业,有研究[25]表明灯诱渔业渔获的个体相对较小,因此采样的季节和使用的渔具渔法可能是造成东海南北海域种群胴长与体质量差异的原因。此外,东海采样的剑尖枪乌贼优势胴长和体质量明显高于南海,可能是由于该海域内多条海流交汇,海底地形复杂多样[14, 26],为其提供丰富的营养物质和天然的庇护所,从而使得种群肥满度较高[8, 26-27]。
本研究表明,两年间雌性个体的平均胴长和体质量均大于雄性,然而雄性个体的胴长和体质量年间差异较大,这与2009—2010年在东海南部采样的剑尖枪乌贼研究结果[12]相一致。FORSYTHE[28]和TAFUR等[29]研究表明,茎柔鱼(Dosidicus gigas)的生长发育受海水温度影响明显,水温升高加快个体的成熟速度,使积累的能量更多投入到生殖过程,导致个体体型较小,剑尖枪乌贼同样也存在类似的情况[12, 29-30]。此外,产卵种群的洄游路线以及距离的长短也会影响种群大小结构的时空变化[31-32]。因此,剑尖枪乌贼雄性个体的索饵洄游距离可能较长,消耗了大量的能量,同时海域水温的季节变化以及自身的繁殖策略可能加快个体的发育,导致雄性个体年间差异较大。
剑尖枪乌贼年间、季节资源量以及种群组成波动较大[33-34],种群对栖息海域的海水温度、盐度以及海流等环境因素的变化敏感[35],并且不同季节产卵种群进入渔场的时期以及生长特性均存在差异[12],因此,雌雄个体的胴长和体质量年间差异可能与产卵种群的生长特征以及海域环境要素的变化有关。
3.2 体质量-胴长关系生长参数b的年间变化可以作为海洋生物对海域环境变化能力的指标[36]。本研究表明,两年中雌雄个体生长参数b的范围为2.32~2.53,并且2017年雌性个体的b值小于2018年,而雄性个体两年中b值相近,这表明研究种群为负异速生长,雌性个体对栖息环境的年间变化更加敏感[35],经协方差分析发现,2018年渔汛期雌雄个体体质量-胴长关系差异不显著(P>0.05),这可能由于两年栖息环境的变化引起雌雄个体的生长关系差异不明显。2003—2008年和2007年分别对东海南部和南海海域采样研究[35-38]表明,生长参数b的范围分别为1.97~2.10和2.14~2.32,并且两个海域b值的范围均小于东海北部海域,这可能与海域环境、种群组成结构、生活史特征及捕捞强度[25]等因素有关。相较于北部海域,南部海域的海流较弱,食物组成较为缺乏,因此剑尖枪乌贼的生长也会受到一定限制。另外,角质颚形态的不同特征也可能会影响到摄食,进而影响剑尖枪乌贼的生长[39-40]。
3.3 性腺成熟度组成本研究表明,两年中雄性个体的性成熟胴长小于雌性,并且雌雄个体在较小的胴长组下即可达到性成熟。PIERCE等[41]和GUERRA等[42]研究发现,在较小胴长下成熟的雄性个体,可能是与不同时期性成熟的雌性进行生殖交配,使其种群繁衍下去的一种繁殖策略,这与其他闭眼亚目种类的繁殖情况[41]相似,也与2003—2008年东海南部采样研究的结果[37]相一致。种群为了响应捕捞强度的变化,会调节自身生理机制,以更多种群数量、体型较小的个体来提高种群的繁殖能力[43],从而保证种群延续下去的目的,这种响应机制可能导致两年间性成熟胴长的降低。此外,栖息海域海水温度的变化也会影响枪乌贼个体性成熟的大小[12]。因此,不同胴长组下雌雄个体性腺成熟程度的差异可能与自身繁殖策略、捕捞强度以及海域水温的年间变化有关。
王凯毅[11]研究表明,春季产卵种群雌雄个体的初次性成熟胴长为分别为163 mm和139 mm,夏季产卵种群体型较小,数量较多[44],结合本研究胴长与体质量组成、初次性成熟胴长以及胴长绝对生长率分析判断,两年中采样的剑尖枪乌贼均为春季和夏季产卵种群,并且2018年个体中夏季产卵种群的幼体数量较多。
3.4 生长速率王凯毅[11]通过耳石日轮计数研究发现,个体生长200天后,雄性个体的胴长生长速率略快于雌性,而本研究表明,雌性个体的胴长生长速率快于雄性,并且雌雄个体胴长生长速率年间差异不大,而体质量生长速率差异较大,这可能与雌雄个体的生长阶段[12]、繁殖策略[37, 41]、摄食习性[8, 45]以及年间采样种群的组成有关。剑尖枪乌贼具有世代更新快、资源量补充和恢复性强的特征,并且个体生长发育和生活史不同阶段栖息的海域环境特征存在差异,导致个体的生长速率也随之发生改变[46]。
4 展望研究剑尖枪乌贼渔业生物学特性,对了解其种群生物特征以及年间变化具有一定科学意义,但缺少夏季采样时期的样本,导致已有样本并不能充分说明生物学特征的年间差异,因此,建议今后需通过剑尖枪乌贼耳石、角质颚等硬组织结构并结合微量元素等方法,探究其不同产卵群体的渔业生物学特征,并分析影响其差异的生物(摄食和繁殖等)和非生物因子(海水温度和盐度等),为揭示其生活史特征以及该资源的可持续开发利用提供科学依据和理论指导。
| [1] |
陈新军, 刘必林, 王尧耕. 世界头足类[M]. 北京: 海洋出版社, 2009. CHEN X J, LIU B L, WANG Y G. Cephalopods in the world[M]. Beijing: China Ocean Press, 2009. |
| [2] |
徐至正.利用平衡石结构及化学讯号判别台湾东北部海域剑尖枪锁管之孵化群差异[D].基隆: 台湾海洋大学, 2014. HSU Z Z. Hatching cohort discrimination of swordtip squid (Uroteuthis edulis) off northeastern Taiwan in statolith structure and chemical signatures[D]. Keelung: Taiwan Ocean University, 2014. |
| [3] |
JEREB P, ROPER C F E. Cephalopods of the world. An annotated and illustrated catalogue of cephalopod species known to date. Volume 2. Myopsid and Oegopsid squids. FAO Species Catalogue for Fishery Purposes, No.4 Vol 2[R]. Rome, Italy: FAO.
|
| [4] |
JEREB P, ROPER C F E. Cephalopods of the Indian Ocean. A review. Part Ⅰ. Inshore squids (Loliginidae) collected during the International Indian Ocean Expedition[J]. Proceedings of the Biological Society of Washington, 2006, 119(1): 91-136. DOI:10.2988/0006-324X(2006)119[91:COTIOA]2.0.CO;2 |
| [5] |
曾建豪.台湾东北部海域剑尖枪锁管族群丰度时空变动模式分析[D].基隆: 台湾海洋大学, 2012. TSENG C H. The spatio-temporal pattern of swordtip squid Uroteuthis (Photo Uroteuthis) edulis stock abundance in the northeastern waters of Taiwan[D]. Keelung: Taiwan Ocean University, 2012. |
| [6] |
凌建忠, 郑元甲. 东海黄海头足类资源量的评估[J]. 海洋渔业, 2000, 22(2): 60-62. LING J Z, ZHENG Y J. Stock assessment of cephalopoda in East China Sea and Yellow Sea[J]. Marine Fisheries, 2000, 22(2): 60-62. DOI:10.3969/j.issn.1004-2490.2000.02.004 |
| [7] |
凌建忠, 严利平, 李建生. 应用资源密度面积法估算东海区头足类资源量[J]. 海洋渔业, 2008, 30(1): 56-61. LING J Z, YAN L P, LI J S. Application of resources density/area method to estimate cephalopod stock of the East China Sea[J]. Marine Fisheries, 2008, 30(1): 56-61. DOI:10.3969/j.issn.1004-2490.2008.01.010 |
| [8] |
NATSUKARI Y, NAKANOSE T, ODA K. Age and growth of loliginid squid Photololigo edulis (Hoyle, 1885)[J]. Journal of Experimental Marine Biology and Ecology, 1988, 116(2): 177-190. DOI:10.1016/0022-0981(88)90054-8 |
| [9] |
刘宗祐.利用形态测量法探讨剑尖枪锁管与台湾锁管族群分布结构之研究[D].基隆: 台湾海洋大学, 2004. LIU T Y. Studies on the population structure of swordtip (Uroteuthis edulis) and mitre (Uroteuthis chinensis) squids by using morphometric method[D]. Keelung: Taiwan Ocean University, 2004. |
| [10] |
董正之. 世界大洋经济头足类生物学[M]. 济南: 山东科学技术出版社, 1991. DONG Z Z. Biology of the economic species of cephalopods in the world oceans[M]. Jinan: Shandong Science and Technology Press, 1991. |
| [11] |
王凯毅.台湾东北部陆棚海域剑尖枪锁管生活史之研究[D].基隆: 台湾海洋大学, 2009. WANG K Y. Life history of Uroteuthis (Photo Uroteuthis) edulis (Hoyle, 1885) in the Shelf Waters off Northeastern Taiwan[D]. Keelung: Taiwan Ocean University, 2009. |
| [12] |
WANG K Y, CHANG K Y, LIAO C H, et al. Growth strategies of the swordtip squid, Uroteuthis edulis, in response to environmental Changes in the Southern East China Sea:a cohort analysis[J]. Bulletin of Marine Science, 2013, 89(3): 677-698. |
| [13] |
王凯毅.台湾东北部海域剑尖枪锁管分布与洄游之研究[D].基隆: 台湾海洋大学, 2003. WANG K Y. Study on the distribution and migration of Uroteuthis (Photo Uroteuthis) edulis (Hoyle, 1885) in the northeastern waters of Taiwan[D]. Keelung: Taiwan Ocean University, 2003. |
| [14] |
王友喜. 东海南部剑尖枪乌贼渔业生物学特性[J]. 海洋渔业, 2002, 24(4): 169-172. WANG Y X. Fishery biological characteristics of swordtip squid Loligo edulisin the Southern Part of the East China Sea[J]. Marine Fisheries, 2002, 24(4): 169-172. DOI:10.3969/j.issn.1004-2490.2002.04.005 |
| [15] |
LIAO C H, LEE M A, LAN Y C. The temporal and spatial change in position of squid fishing ground in relation to oceanic features in the northeastern waters of Taiwan[J]. Journal of the Fisheries Society of Taiwan, 2006, 33(2): 99-113. |
| [16] |
何欣颖.海洋环境变动对台湾东北部海域棒受网渔业剑尖枪锁管渔获率与分布关系之研究[D].基隆: 台湾海洋大学, 2016. HO H Y. Ocean variations in relate to catch rates and distributions of swordtip squid (Uroteuhis eduilis) of stick-held dip net fishery in the northeastern waters of Taiwan[D]. Keelung: Taiwan Ocean University, 2016. |
| [17] |
郑元甲. 进一步开发和合理利用东海鱿鱼资源[J]. 海洋渔业, 1997(2): 51-53. ZHENG Y J. Further development and rational utilization of squid resources in the East China Sea[J]. Marine Fisheries, 1997(2): 51-53. |
| [18] |
YODA M. Stock assessment of Uroteuthis edulis in Japan Sea and East China Sea[J]. Bulletin of Seikai National Fisheries Research Institute, 2011, 77(2): 89-92. |
| [19] |
CHANG K Y, CHEN C S, CHIU T Y, et al. Argentine shortfin squid (Illex argentinus) stock assessment in the southwest Atlantic using geostatistical techniques[J]. Terrestrial, Atmospheric and Oceanic Sciences, 2016, 27(2): 281-292. DOI:10.3319/TAO.2015.11.05.01(Oc) |
| [20] |
LIPIN'SKI M R, UNDERHILL L G. Sexual maturation in squid:quantum or continuum?[J]. South African Journal of Marine Science, 1995, 15(1): 207-223. |
| [21] |
TAFUR R, VILLEGAS P, RABIÍ M, et al. Dynamics of maturation, seasonality of reproduction and spawning grounds of the jumbo squid Dosidicus gigas (Cephalopoda:Ommastrephidae) in Peruvian waters[J]. Fisheries Research, 2001, 54(1): 33-50. |
| [22] |
RICKER W E. Computation and Interpretation of Biological Statistics of Fish Populations[J]. Journal of Wildlife Management, 1977, 41(1). |
| [23] |
李建柱, 陈丕茂, 贾晓平, 等. 中国南海北部剑尖枪乌贼资源现状及其合理利用对策[J]. 中国水产科学, 2010, 17(6): 1309-1318. LI J Z, CHEN P M, JIA X P, et al. Resources status and conservation strategy of Loligo edulis hoyle in the northern South China Sea[J]. Journal of Fishery Sciences of China, 2010, 17(6): 1309-1318. |
| [24] |
卢昌彩, 赵景辉. 东海伏季休渔制度回顾与展望[J]. 渔业信息与战略, 2015, 30(3): 168-174. LU C C, ZHAO J H. The review and prospect on fish moratorium policy in the East China Sea[J]. Fishery Information & Strategy, 2015, 30(3): 168-174. |
| [25] |
陈姿莹.台湾东北部不同海域及不同渔法采样下剑尖枪锁管渔业生物学之差异[D].基隆: 台湾海洋大学, 2012. CHEN Z Y. The fisheries biology of Uroteuthis (Photo Uroteuthis) edulis caught by different sea areas and different fishing methods in the northeastern waters of Taiwan[D]. Keelung: Taiwan Ocean University, 2012. |
| [26] |
宋万先. 1984年6-7月东海黑潮及其邻近海域的水文特征[C]//黑潮调查研究论文集.北京: 海洋出版社, 1987. SONG W X. Hydrological characteristics of kuroshio and its adjacent sea areas in the East China Sea in June and July 1984[C]//. Beijing: China Ocean Press, 1987. |
| [27] |
WANG K Y, LIAO C H, LEE K T. Population and maturation dynamics of the swordtip squid (Photololigo edulis) in the southern East China Sea[J]. Fisheries Research, 2008, 90(1/3): 178-186. |
| [28] |
FORSYTHE J W. Accounting for the effect of temperature on squid growth in nature:from hypothesis to practice[J]. Marine and Freshwater Research, 2004, 55(4): 331-339. DOI:10.1071/MF03146 |
| [29] |
TAFUR R, KEYL F, ARGVELLES J. Reproductive biology of jumbo squid Dosidicus gigas in relation to environmental variability of the northern Humboldt Current System[J]. Marine Ecology Progress Series, 2010, 400: 127-141. DOI:10.3354/meps08386 |
| [30] |
PECL G T, JACKSON G D. The potential impacts of climate change on inshore squid:biology, ecology and fisheries[J]. Reviews in Fish Biology and Fisheries, 2008, 18(4): 373-385. DOI:10.1007/s11160-007-9077-3 |
| [31] |
IBÁÑEZ C M, CUBILLOS L A. Seasonal variation in the length structure and reproductive condition of the jumbo squid Dosidicus gigas (d'Orbigny, 1835) off central-south Chile[J]. Scientia Marina, 2007, 1(1): 123-128. |
| [32] |
WELLS M J, CLARKE A. Energetics:the costs of living and reproducing for an individual cephalopod[J]. Philosophical Transactions of the Royal Society B:Biological Sciences, 1996, 351(1343): 1083-1104. DOI:10.1098/rstb.1996.0095 |
| [33] |
王晓晴, 章俊. 一九九四年和一九九五年夏季浙江渔场头足类资源量评估[J]. 浙江水产学院学报, 1998, 17(2): 96-101. WANG X Q, ZHANG J. Evaluation on the cephalopod biomass in the Zhejiang fishing ground in the summers of 1994 and 1995[J]. Journal of Zhejiang College of Fisheries, 1998, 17(2): 96-101. |
| [34] |
朱文斌, 薛利建, 卢占晖, 等. 东海南部海域头足类群落结构特征及其与环境关系[J]. 海洋与湖沼, 2014, 45(2): 436-442. ZHU W B, XUE L J, LU Z H, et al. Cephalopod community structure and its relationship with environmental factors in the southern East China Sea[J]. Oceanologia et Limnologia Sinica, 2014, 45(2): 436-442. |
| [35] |
许语婕.东海南部剑尖枪锁管之资源评估[D].基隆: 台湾海洋大学, 2015. HSU Y J. Stock assessment of swordtip squid (Uroteuthis edulis) in the southern East China Sea[D]. Keelung: Taiwan Ocean University, 2015. |
| [36] |
朱立新, 侯刚, 梁振林. 基于贝叶斯方法的黄海北部鳀鱼体长与体重关系式参数估计[J]. 中国水产科学, 2015, 22(4): 757-769. ZHU L X, HOU G, LIANG Z L. Parameter estimation of the weight-length relationship of Japanese anchovy in the north Yellow Sea using Bayesian methods[J]. Journal of Fishery Sciences of China, 2015, 22(4): 757-769. |
| [37] |
王凯毅, 张可扬, 李国添, 等. 南东海剑尖枪锁管体型与生殖状况之时空变化[J]. 台湾水产研究, 2010, 18(1): 13-32. WANG K Y, ZHANG K Y, LEE K T, et al. Spatial and temporal variations in the length structure and reproductive condition of the Uroteuthis (Photo Uroteuthis) edulis in the southern East China Sea[J]. Journal of Taiwan Fisheries Research, 2010, 18(1): 13-32. |
| [38] |
孙典荣, 李渊, 王雪辉, 等. 北部湾剑尖枪乌贼生物学特征及资源状况变化的初步研究[J]. 南方水产科学, 2011, 7(2): 8-13. SUN D R, LI Y, WANG X H, et al. Biological characteristics and stock changes of Loligo edulis in Beibu Gulf, South China Sea[J]. South China Fisheries Science, 2011, 7(2): 8-13. DOI:10.3969/j.issn.2095-0780.2011.02.002 |
| [39] |
方舟, 陈新军, 金岳. 基于角质颚的东、黄海头足类科类判别[J]. 上海海洋大学学报, 2019, 28(3): 365-372. FANG Z, CHEN X J, JIN Y. Preliminary study of species identification of cephalopod in East China Sea and Yellow Sea based on beak morphology[J]. Journal of Shanghai Ocean University, 2019, 28(3): 365-372. |
| [40] |
马迪, 金岳, 陈芃, 等. 基于角质颚形态的东海2种常见乌贼类的种类判别[J]. 上海海洋大学学报, 2018, 27(4): 594-602. MA D, JIN Y, CHEN P, et al. Species identification of Sepioidea in the East China Sea based on beak morphology[J]. Journal of Shanghai Ocean University, 2018, 27(4): 594-602. |
| [41] |
PIERCE G J, BOYLE P R, HASTIE L C, et al. The life history of Loligo forbesi (Cephalopoda:Loliginidae) in Scottish waters[J]. Fisheries Research, 1994, 21(1/2): 17-41. |
| [42] |
GUERRA A, ROCHA F. The life history of Loligo vulgaris and Loligo forbesi (Cephalopoda:Loliginidae) in Galician waters (NW Spain)[J]. Fisheries Research, 1994, 21(1/2): 43-69. |
| [43] |
SHAN X J, LI X S, YANG T, et al. Biological responses of small yellow croaker (Larimichthys polyactis) to multiple stressors:a case study in the Yellow Sea, China[J]. Acta Oceanologica Sinica, 2017, 36(10): 39-47. DOI:10.1007/s13131-017-1091-2 |
| [44] |
丁天明, 宋海棠. 东海剑尖枪乌贼生物学特征[J]. 浙江海洋学院学报(自然科学版), 2000, 19(4): 371-374. DING T M, SONG H T. Biological characteristics of Loligo edulis Hoyle in the East China Sea[J]. Journal of Zhejiang Ocean University (Natural Science), 2000, 19(4): 371-374. DOI:10.3969/j.issn.1008-830X.2000.04.014 |
| [45] |
林静远, 刘必林, 桓梦瑶, 等. 我国几种常见头足类角质颚碳氮稳定同位素分析[J]. 上海海洋大学学报, 2019, 28(3): 394-401. LIN J Y, LIU B L, HUAN M Y, et al. Carbon and nitrogen stable isotopes analysis of the common cephalopods beaks in the offshore of China[J]. Journal of Shanghai Ocean University, 2019, 28(3): 394-401. |
| [46] |
CADRIN S X. Advances in morphometric identification of fishery stocks[J]. Reviews in Fish Biology and Fisheries, 2000, 10(1): 91-112. DOI:10.1023/A:1008939104413 |
2. Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources, Shanghai Ocean University, Ministry of Education, Shanghai 201306, China;
3. National Engineering Research Center for Oceanic Fisheries, Shanghai Ocean University, Shanghai 201306, China;
4. Key Laboratory of Oceanic Fisheries Exploration, Ministry of Agriculture and Rural Affairs, Shanghai 201306, China;
5. Scientific Observing and Experimental Station of Oceanic Fishery Resources, Ministry of Agriculture and Rural Affairs, Shanghai 201306, China