 2020,
Vol. 29
2020,
Vol. 29


2. 国家远洋渔业工程技术研究中心, 上海 201306;
3. 大洋渔业资源可持续开发教育部重点实验室, 上海 201306;
4. 远洋渔业协同创新中心, 上海 201306
日本鲭(Scomber japonicus)为近海中上层鱼类, 在0~300 m的水层栖息, 在太平洋和大西洋温带水域以及大陆架及其邻近海域广泛分布, 灯光围网是主要捕捞方式之一[1-3]。毛里塔尼亚海域地处几内亚湾暖流和加纳利寒流交汇区, 伴随较强的涌升流, 初级生产力较高, 是西北非重要渔场之一[4]。我国渔船从2006年开始在毛里塔尼亚海域捕捞日本鲭等中上层鱼类, 毛里塔尼亚2015—2019年渔业规划中, 第一条就是要更好地保护海洋渔业资源, 更大力度地开发中上层渔业资源。因此, 研究毛里塔尼亚海域以日本鲭为例的小型中上层渔业资源分布十分必要。
目前国外学者对日本鲭的研究主要集中在生物学, 如ALLAYA等[5]和MACHADO等[6]分别研究了大西洋日本鲭的形态变异和肝脏转录组的重装和标记。国内学者主要利用栖息地适宜性指数(HSI)模型和广义加性(GAM)模型对中国近海的日本鲭进行研究, 如:YU等[7]利用2006—2013年东海日本鲭的数据建立栖息地适宜性指数模型, 并用2014—2015年的数据验证, 得出最佳HSI模型能够有效地预测日本鲭的生境适宜性; LI等[8]利用HSI模型确定东海日本鲭每个月的热点; 王从军等[9]通过GAM模型分析海洋环境要素对东、黄海日本鲭的资源丰度的影响; 李显森等[10]利用渔获量重心法、GAM模型等方法研究了1—8月中东大西洋中部海域鲭鱼渔场的时空变化。随着GIS技术的发展, HSI模型在渔业领域中的应用日渐成熟且有其他生境模型不具备的优越性[11], 但在生境资料获取的客观性和全面性、SI曲线的可靠性、样本反映总体数据的代表性以及模型结构等方面尚存在一定局限性[12], 且其假设前提中存在弊端[13], 根据实测数据对模型不断修正才能逐步提高预测的准确性; GAM模型能够较好评估环境因子对资源的影响程度, 但无法准确地反映出测量因素与生物量之间的关系[14]。分位数回归模型最早由KOENKER等[15]提出, 已在研究鱼类的栖息地综合指数方面得到了应用[16-17], 相对HSI、GAM模型, 分位数回归模型能够克服传统的相关和回归统计中的不足, 并考虑各个环境因子对日本鲭分布的影响权重的不同条件及其相互作用。本文根据2017年在毛里塔尼亚海域测定的海洋环境数据以及日本鲭的渔获率统计资料, 用分位数回归和GAM模型探究日本鲭时空分布与海洋环境的关系, 旨在进一步了解日本鲭在毛里塔尼亚海域的时空分布特征, 准确预报渔情, 为提高我国渔船在此海域的作业效率, 为毛里塔尼亚养护和管理日本鲭等小型中上层渔业资源提供参考。
1 材料与方法 1.1 调查方法 1.1.1 调查船及基本信息“福远渔097”灯光围网船总长49.95 m, 型宽8.60 m, 型深4.00 m。设计吃水3.30 m, 设计排水量896.90 t, 主机功率为828 kW, 航速为6~11 kn, 鱼舱容量190 m3, 油舱容积120 m3, 淡水舱容积47 m3。航区为国际Ⅰ区。
1.1.2 调查时间、海域及站点分布调查时间为2017年9月20日至12月31日。调查海域为16°28'W~17°59'W和17°50'N~20°58'N。作业时, 当天的下网位置记录为对应的站点位置, 共112个调查站点。随机选择78%的站点数据用作建模, 剩下22%的站点数据用以模型验证, 站点分布见图 1。部分建模站点和验证站点出现重合, 原因是这些站点的下网位置相同。

|
图 1 毛里塔尼亚海域调查站点 Fig. 1 Modeling sites and verification sites in the waters near Mauritania |
“福远渔097”所用围网网具主体由网衣、包铅绳、浮子、上纲和下纲组成, 网口网衣拉直周长约1 643.4 m, 网口至囊端拉直长度约116 m, 上纲长度约809.84 m, 下纲长度约761.44 m。
1.1.4 调查仪器、方法及内容利用渔船上的全球定位仪记录每天放网开始时的经纬度, 同时记录渔获质量。调查站点预先确定, 但实际调查位置与计划站点存在一定偏差。每日放网后或起网前, 使用加拿大型号为XR-620的多功能水质仪, 测量调查站点海表面的叶绿素a浓度、海表面温度以及盐度数据。
1.2 数据处理方法 1.2.1 CPUE的计算方法单位捕捞努力量渔获量(catch per unit effort, CPUE)通常用来表示鱼类资源丰度的相对指数, 是对某个渔业资源进行评估的基本内容之一[18]。船上所得到的直接数据为渔获量, 建立关系模型前, 需要计算日本鲭的CPUE值, 记为R。计算方法如下:
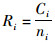 (1)
(1)
式中:Ri为i站点的当天CPUE值, t/网; Ci为在i站点当天的渔获量, t; ni为i站点当天的网次数量, 网。对CPUE数据进行了非参数检验, CPUE符合正偏态分布。
1.2.2 海洋环境因子值的计算方法海洋环境因子数据为各站点10 m以内所测得的各海洋环境因子数据的算术平均值[17]。
1.3 分位数回归建模 1.3.1 对变量多重共线性分析为排除海表面叶绿素a浓度、海表面温度以及海表面盐度间可能存在的高相关性, 建模前对环境因子间进行多重共线性分析, 删除具有多重共线性的环境变量。根据方差扩大因子VIF判断变量之间是否存在共线性(VIF<10)[19-20]。多重共线性分析在SPSS 23.0中实现。
1.3.2 建立日本鲭CPUE预测模型分位数回归建模的具体方法参照文献[21]。分位数回归分析采用由美国地理调查局中陆生态科学研究中心开发的统计学软件Blossom[17]完成。
通过对环境变量多重共线性分析, 确定进行建模的环境变量, 根据建模站点的单位捕捞努力量渔获量(Ri)、叶绿素a浓度、海表面温度、海表面盐度及其交互作用项建立日本鲭CPUE与海洋环境因子的关系模型, 其一般形式为
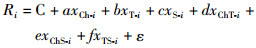 (2)
(2)
式中:C为常数项; a~f为自变量的系数; xChT-i表示叶绿素a浓度和海表面温度的交互作用项; xChS-i表示叶绿素a浓度和海表面盐度的交互作用项; xTS-i表示海表面温度和海表面盐度的交互作用项; ε为误差项, 通常假设其均值为0, 方差为δ2; i表示站点号数, 如xCh-i表示在i站点的叶绿素a浓度。
1.4 GAM评价环境因子对CPUE的影响分位数回归完成建模后, 利用GAM模型对环境因子与CPUE的关系进行分析, 评价各环境因子对CPUE的影响程度。GAM的表达式:
 (3)
(3)
式中:R为CPUE值; s为自然立方样条平滑函数; ε表示残差, E(ε)=0, ε=δ2符合正态分布。
根据赤池信息准则(akaike information criterion, AIC)选择AIC最小值的信息统计量, 逐步去除或添加变量以得到最佳模型[22]。利用AIC准则检验逐步调整模型的拟合程度, AIC越小, 拟合效果越好。利用F检验评价环境因子对CPUE影响的显著性[23]。有关GAM的操作通过R 3.6.1软件实现。
1.5 栖息地综合指数(IHI)计算方法HSI是对特定种类的潜在栖息地进行量化, 并使用0~1之间的数值来描述最佳结构的适合性[24-25], IHI是基于“鱼类栖息地”和“栖息地适宜性指数”的概念而提出的, 不同的种类、种群和调查数据可得出不同的“栖息地适宜性指数”[25]。根据关系模型, 得出建模站点和验证站点的预测CPUE, 按照下列公式计算得出日本鲭在不同站点的IHI值(记为I):
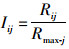 (4)
(4)
式中:Rij是指j组(j=1为建模站点; j=2为验证站点)i站点的CPUE预测值; Rmax-j是指j组中CPUE预测值中的最大值。
1.6 IHI模型预测能力的验证和评价把验证站点的环境数据逐一代入关系模型中, 使用SPSS 23.0软件, 通过计算日本鲭的CPUE实际值与预测值之间的Spearman相关系数(秩相关系数)[26], 结合双尾检验对IHI模型的预测能力进行评价。
用Arcmap 10.3作图得出建模站点和验证站点的IHI等值线与实测CPUE的分布图。将验证站点的实测CPUE值除以其中的最大值得出IHI'(记为I'), 用Excel作出IHI'和IHI的变化趋势图, 定性评价模型的预测能力。
 (5)
(5)
式中:Ri-a是指i站点的实测CPUE; Rmax-a是指实测CPUE中的最大值。
2 结果 2.1 多重共线性诊断结果利用SPSS对海表面温度(SST)、叶绿素a浓度、海表面盐度进行共线性诊断分析, 结果如表 1所示。VIF值都低于10, 说明环境因子间无共线性, 可用这些环境变量进行建模。

|
表 1 多重共线性诊断结果 Tab.1 Multicollinearity diagnosis results |
拟合得出的最佳预测模型如下:
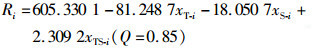 (6)
(6)
由式(6)可知, 影响日本鲭CPUE的海洋环境因素包括海表面温度、海表面盐度和温盐交互项, P依次为0.005、0.032和0.024。
2.2 分位数回归得出的预测模型利用Blossom建立CPUE与海洋环境的关系模型, 将每次拟合得出的模型系数和对应的P记录下来, 取分位数(用Q表示)最高的模型为最佳模型。见表 2。

|
表 2 最佳CPUE预测模型的参数估计 Tab.2 Estimation of parameters of the optimal CPUE prediction model |
GAM对环境因子的分析表明, 逐步回归的最终模型解释能力为64.1%, 在所有因子当中, 海表面温度、海表面盐度、温盐交互项所组成的模型为最优模型。GAM模型的统计结果如表 3所示。F检验表明, 海表面温度对日本鲭CPUE的影响最显著, 其次是温盐的交互作用和海表面盐度。海洋环境因子与CPUE关系如图 2所示, 可知日本鲭偏好的海表面温度为19~22 ℃, 日本鲭偏好的海表面盐度为36.3左右。

|
表 3 环境因子对CPUE影响的GAM模型统计结果 Tab.3 Statistical results of GAM about the influence of environmental variables to CPUE |

|
图 2 海表面温度、海表面盐度与CPUE关系的GAM分析图 Fig. 2 Relationship between SST and SSS, and CPUE based on the generalized additive model |
对建模站点和验证站点的CPUE实测值与预测值进行Wilcoxon(符号秩)检验, 结果见表 4, 两者的P均高于显著水平0.05, 说明建模站点和验证站点的CPUE实测值与预测值之间均无显著性差异。

|
表 4 建模站点和验证站点的实测CPUE与预测CPUE之间Wilcoxon检验结果 Tab.4 The result of Wilcoxon test between the observed and predicted CPUEs of the modeling sites and validation sites |
对验证站点的实测CPUE和预测CPUE的相关系数和显著性进行双尾检验, 得出实测CPUE与预测CPUE的Spearman相关系数达到0.803, 相伴概率小于0.01, 这说明显著性水平为0.01时, 实测CPUE和预测CPUE之间高度相关, 模型可靠。
2.5 IHI模型的定性评价由图 3可知, CPUE和IHI具有相关性:IHI在低于0.24时, CPUE普遍较低; IHI在0.26~0.38时, CPUE增大, 虽偶有较高值出现但平均值不高; IHI大于0.40时, CPUE较高值出现的频率显著增加, 且CPUE大于10 t/网集中出现; 当IHI大于0.42时, CPUE达到最大。

|
图 3 建模站点IHI等值线和实测CPUE分布 Fig. 3 IHI contour and CPUE distribution for modeling sites |
观察图 3可得, CPUE较低值主要集中在以下两个区域:16°20'W~16°50'W和17°30'N~19°00'N, 对应的IHI在0~0.24, 在这个范围内, CPUE不高, 平均在1~5 t/网; 17°20'W~17°30'W和19°40'N~20°00'N范围内, CPUE相对较高, 在5~10 t/网; CPUE较高值集中在16°50'W~17°10'W和19°10'N~19°40'N两个区域, 对应的IHI在0.24~0.28, CPUE值大于15 t/网的出现了数次; 在17°25'W~17°40'W和20°20'N~20°45'N范围内, 对应的IHI大于0.40。
由图 4可得出, CPUE和IHI具有明显的相关性:IHI为0~0.20时, 对应的海域为16°30'W~17°15'W和17°45'N~19°45'N, 此时的CPUE是验证站点中最低的, 普遍低于5 t/网; IHI在0.22~0.32时, 即17°30'W~17°40'W和19°50'N~20°20'N, CPUE较高值出现的频率随着IHI的增加而增加; IHI>0.32时, 即在17°30'W~17°45'W和20°20'N~20°45'N区域, CPUE的较高值集中出现, 分布密集。这说明IHI模型预测效果良好。

|
图 4 验证站点IHI等值线和实测CPUE分布 Fig. 4 IHI contour and CPUE distribution for the verification sites |
由图 5可知, IHI的变化趋势与IHI'高度一致。由此可见, 建立的模型能较好地预测CPUE。综上, IHI较高的海域对应较高的实测CPUE, 因此基于分位数回归拟合得出的关系模型能较好地反映CPUE与海洋环境因子的关系, 预测能力良好。

|
图 5 验证站点IHI'和IHI的变化趋势 Fig. 5 Trends of IHI' and IHI for the verification sites |
分位数回归已经发展成一套成熟的统计模型, 其方法有系统的统计学理论支撑[27-29]。建立分位数回归模型依据的概念是最小绝对偏差[30-31], 因此分位数回归基本可以克服传统模型易受极值影响的缺点。又因本文所使用的样本数据在时间跨度上只有4个月, 总数也只有112个, 用作建模的站点数量并不多, 这些数据没有呈现出正态分布的特点, 而是呈正偏态分布。分位数回归的一个优点就是对数据的分布不需要满足正态分布, 分位数回归模型能更好地反映因变量的平均水平[32]。
3.2 IHI分布高低的原因结合最佳关系模型和GAM模型可得, 海洋环境因子对日本鲭CPUE的影响程度为海表面温度、温盐交互作用以及海表面盐度, 叶绿素a浓度对CPUE没有显著影响。GAM模型得出日本鲭适宜的温度和盐度范围分别为19~22 ℃和36.3左右。李显森等[10]利用渔获量重心法、GAM模型等方法在研究1—8月中东大西洋中部海域鲭鱼渔场的时空变化时得出, 鲭鱼渔场重心的海表温度在20~21 ℃居多; 逄志伟等[33]利用GAM模型等方法研究中东大西洋海域中上层鱼类CPUE的月变化、时空分布与海表面温度等环境因子的关系时, 得出渔场的最适SST为20~22 ℃, 本文得出的结论与其范围相当; 郑波等[34]利用GLM和GAM模型研究东、黄海日本鲭渔场与环境因子的关系时得出, 适宜的海表盐度在34左右, 本文的结论与其有所不同, 原因在于毛里塔尼亚海域与中国东、黄海的近海水文条件不同。YU等[7]通过选择SSTA(sea surface temperature anomaly)、SSHA(sea surface height anomaly)和NPPA(net primary production anomaly)作为环境预测因子拟合HSI模型, 发现海表面温度的升高会对夏季东海日本鲭的生境适宜性产生积极的影响; 王从军等[11]通过GAM模型分析1999—2011年海洋环境因素对东、黄海日本鲭的资源丰度的影响时, 得出海表面温度对日本鲭的资源丰度影响最大; 李显森等[10]在研究1—8月中东大西洋中部海域日本鲭渔场的时空变化时得出, 海洋环境要素的显著性强弱顺序依次为海表面温度、经度和叶绿素a浓度。本文同样得出海表面温度对日本鲭的资源丰度影响最大, 这也可从实际情况得到解释:毛里塔尼亚海域位于几内亚湾暖流和加纳利寒流的交汇地带, 有利于渔场形成, 寒暖水团对应着温度的变化, 因此海表面温度对日本鲭的影响至关重要。YU等[7]认为东海日本鲭的渔场与叶绿素a浓度成负相关关系; 李纲等[35]利用渔获产量研究海洋环境因子对东海日本鲭渔场分布的影响, 得出叶绿素a浓度不是形成日本鲭渔场的主要因素。本文认为9—12月叶绿素a浓度对日本鲭的CPUE无显著影响, 原因可能是形成渔场的关键, 主要取决于浮游生物的量和规模[36], 而除了叶绿素a浓度外, 营养盐和海流等其他环境因子同样影响着浮游生物的量和规模; 加上本次调查覆盖的范围不够大, 叶绿素a浓度的变化不大, 海表面初级生产力差异小, 因此其影响不明显。
日本鲭的分布和洄游与几内亚湾暖流和加纳利寒流的交汇场变化有关。9—12月, IHI分布较高的区域在17°25'W~17°45'W和20°15'N~20°45'N, 对应在图 2和图 3中的偏北方向。9—10月东北信风带向北移动, 几内亚湾暖流增强, 加纳利寒流变弱, 此后大陆架上盛行向北的海流, 因此日本鲭向北聚集, 资源丰度较高。10—12月, 北半球入冬, 温度降低, 日本鲭南迁, 进行越冬洄游[10], 但总体还是在偏北方向, 相应的IHI也较高。
IHI分布较低的区域在16°02'W~16°28'W和17°30'N~18°50'N, 为东南方向。原因在于9—10月北部有更好的寒暖流交汇场, 日本鲭北游, 因此东南区域日本鲭资源丰度较低, IHI值也较低。
3.3 建议IHI模型能较准确地预测日本鲭的CPUE。日本鲭的IHI分布较高和资源丰度较高的区域在17°25'W~17°45'W和20°15'N~20°45'N, 因此建议我国灯光围网船在毛里塔尼亚海域9—12月作业时, 尽量使下网区域集中在上述区域, 以增加产量, 提高经济效益。日本鲭不仅是毛里塔尼亚海域小型中上层渔业的重要组成部分, 也是金枪鱼类等的主要食物, 因此对其进行养护与管理十分重要。IHI分布较低的区域是16°02'W~16°28'W和17°30'N~18°50'N, 日本鲭主要摄食浮游生物, 因此在9—12月, 可以在此海域适当增加氮等营养盐, 也可以投放少量流木为浮游生物的繁殖创造条件。
3.4 不足与展望在研究渔业资源的时空分布时,深度也是重要的考虑变量之一,因为鱼类被捕获时所处的深度是由该海域内所有的环境因素共同作用而决定的[37]。本文缺乏日本鲭栖息深度的数据,因此空间上未能说明日本鲭的垂直分布。此外,日本鲭的时空变动还与溶解氧、上升流和营养盐分布等因素有关。因此,要想更全面地探索日本鲭时空分布与海洋环境的关系,还要考虑上述影响因子。本次调查有效数据较少,因此之后的研究要尽可能增加站点数量,延长调查时间,扩大调查范围。另外,如有可能,可以尽量采集各水层的浮游生物量和日本鲭的胃含物等数据,为更深入地研究日本鲭的运动模式提供支撑。
| [1] |
唐启升. 中国专属经济区海洋生物资源与栖息环境[M]. 北京: 科学出版社, 2006: 599-607. TANG Q S. Marine biological resources and habitat environment in China's exclusive economic zone[M]. Beijing: Science Press, 2006: 599-607. |
| [2] |
EASTWOOD P D, MEADEN G J, CARPENTIER A, et al. Estimating limits to the spatial extent and suitability of sole (Solea solea) nursery grounds in the Dover Strait[J]. Journal of Sea Research, 2003, 50(2/3): 151-165. |
| [3] |
邓景耀, 赵传纟因. 海洋渔业生物学[M]. 北京: 中国农业出版社, 1991: 413-452. DENG J Y, ZHAO C Y. Marine fishery biology[M]. Beijing: China Agricultural Press, 1991: 413-452. |
| [4] |
韩保平, 方海, 阮雯. 毛里塔尼亚海洋渔业概况[J]. 现代渔业信息, 2011, 26(4): 20-23. HAN B P, FANG H, RUAN W. Outline of marine fisheries in Mauritania[J]. Modern Fisheries Information, 2011, 26(4): 20-23. |
| [5] |
ALLAYA H, BEN FALEH A, REBAYA M, et al. Identification of Atlantic Chub mackerel Scomber colias population through the analysis of body shape in Tunisian waters[J]. Cahiers de Biologie Marine, 2016, 57(3): 195-207. |
| [6] |
MACHADO A M, FELÍCIO M, FONSECA E, et al. A resource for sustainable management:De novo assembly and annotation of the liver transcriptome of the Atlantic chub mackerel, Scomber colias[J]. Data in Brief, 2018, 18: 276-284. DOI:10.1016/j.dib.2018.03.013 |
| [7] |
YU W, GUO A, ZHANG Y, et al. Climate-induced habitat suitability variations of chub mackerel Scomber japonicus in the East China Sea[J]. Fisheries Research, 2018, 207: 63-73. DOI:10.1016/j.fishres.2018.06.007 |
| [8] |
LI G, CHEN X J, LEI L, et al. Distribution of hotspots of chub mackerel based on remote-sensing data in coastal waters of China[J]. International Journal of Remote Sensing, 2014, 35(11/12): 4399-4421. |
| [9] |
王从军, 邹莉瑾, 李纲, 等. 1999-2011年东、黄海鲐资源丰度年间变化分析[J]. 水产学报, 2014, 38(1): 56-64. WANG C J, ZOU L J, LI G, et al. Analysis of the inter-annual variation of chub mackerel abundance in the East China Sea and Yellow Sea during 1999-2011[J]. Journal of Fisheries of China, 2014, 38(1): 56-64. |
| [10] |
李显森, 逄志伟, 朱建成, 等. 中东大西洋中部海域鲐鱼渔场的时空变化初步研究[J]. 水产科学, 2018, 37(1): 31-37. LI X S, PANG Z W, ZHU J C, et al. Spatial-temporal patterns in fishing ground of Scomber japonicus in central Eastern Central Atlantic Ocean[J]. Fisheries Science, 2018, 37(1): 31-37. |
| [11] |
金龙如, 孙克萍, 贺红士, 等. 生境适宜度指数模型研究进展[J]. 生态学杂志, 2008, 27(5): 841-846. JIN L R, SUN K P, HE H S, et al. Research advances in habitat suitability index model[J]. Chinese Journal of Ecology, 2008, 27(5): 841-846. |
| [12] |
龚彩霞, 陈新军, 高峰, 等. 栖息地适宜性指数在渔业科学中的应用进展[J]. 上海海洋大学学报, 2011, 20(2): 260-269. GONG C X, CHEN X J, GAO F, et al. Review on habitat suitability index in fishery science[J]. Journal of Shanghai Ocean University, 2011, 20(2): 260-269. |
| [13] |
范江涛, 张俊, 冯雪, 等. 南沙海域鸢乌贼渔场与海洋环境因子的关系[J]. 上海海洋大学学报, 2019, 28(3): 419-426. FAN J T, ZHANG J, FENG X, et al. Relationship between Sthenoteuthis oualaniensis fishing ground and marine environmental factors in Nansha area[J]. Journal of Shanghai Ocean University, 2019, 28(3): 419-426. |
| [14] |
THOMSON J D, WEIBLEN G, THOMSON B A, et al. Untangling multiple factors in spatial distributions:lilies, gophers, and rocks[J]. Ecology, 1996, 77(6): 1698-1715. DOI:10.2307/2265776 |
| [15] |
KOENKER R, BASSETT JR G W. Regression Quantiles[J]. Econometrica, 1978, 46(1): 33-50. |
| [16] |
冯波, 田思泉, 陈新军. 基于分位数回归的西南太平洋阿根廷滑柔鱼栖息地模型研究[J]. 海洋湖沼通报, 2010(1): 15-22. FENG B, TIAN S Q, CHEN X J. The habitat suitability index of Illex Argentinus by using quantile regression method in the southwest Atlantic[J]. Transactions of Oceanology and Limnology, 2010(1): 15-22. |
| [17] |
宋利明, 高攀峰, 周应祺, 等. 基于分位数回归的大西洋中部公海大眼金枪鱼栖息环境综合指数[J]. 水产学报, 2007, 31(6): 798-804. SONG L M, GAO P F, ZHOU Y Q, et al. Habitat environment integration index of Thunnus obesus in the high seas of the Central Atlantic Ocean based on the quantile regression[J]. Journal of Fisheries of China, 2007, 31(6): 798-804. |
| [18] |
ANN H B. Studies on age and growth of the pacific mackerel Scomber ponicus Houttuyn[J]. Bulletin of Fisheries Research and Development Agency, 1971, 7: 7-24. |
| [19] |
何晓群, 刘文卿. 应用回归分析[M]. 2版. 北京: 中国人民大学出版社, 2007: 171-184. HE X Q, LIU W Q. Applied regression analysis[M]. 2nd ed. Beijing: China Renmin University Press, 2007: 171-184. |
| [20] |
ZHANG T J, SONG L M, YUAN H C, et al. A comparative study on CPUE standardization of bigeye tuna in the Indian Ocean using multi-scale fisheries data and environment data[C]//Proceedings of the 10th Working Party on Methods. Donostia-San Sebastian, Spain, 2019.
|
| [21] |
SONG L M, ZHOU Y Q. Developing an integrated habitat index for bigeye tuna (Thunnus obesus) in the Indian Ocean based on longline fisheries data[J]. Fisheries Research, 2010, 105(2): 63-74. |
| [22] |
薛毅, 陈立萍. R统计建模与R软件[M]. 北京: 清华大学出版社, 2007. XUE Y, CHEN L P. R Statistical modeling and R software[M]. Beijing: Tsinghua University Press, 2007. |
| [23] |
KIM H Y, LEE C W, SHIN J K, et al. Dynamic simulation of the behavior of purse seine gear and sea-trial verification[J]. Fisheries Research, 2007, 88(1/3): 109-119. |
| [24] |
COLE C A, SMITH R L. Habitat suitability indices for monitoring wildlife populations-an evaluation[J]. Trans. No. Amer. Wildl. Nat. Resour. Conf, 1983, 48: 367-375. |
| [25] |
BROOKS R P. Improving habitat suitability index models[J]. Wildlife Society Bulletin, 1977, 25(1): 163-167. |
| [26] |
OKAMOTO H. Standardized Japanese longline CPUE for bigeye tuna in the Atlantic Ocean from 1961 to 2005[J]. Collective Volume of Scientific Papers, International Commission for the Conservation of Atlantic Tunas, 2006, 60(1): 143-154. |
| [27] |
YU K, LU Z, STANDER J. Quantile regression:applications and current research areas[J]. The Statistician, 2003, 52: 331-350. |
| [28] |
李育安. 分位数回归及应用简介[J]. 统计与信息论坛, 2006, 21(3): 35-38, 44. LI Y A. An introduction to quantile regression and it's application[J]. Statistics & Information Forum, 2006, 21(3): 35-38, 44. |
| [29] |
KOENKER R. Quantile regression. Econometric society monographs[M]. London: Cambridge University Press, 2005.
|
| [30] |
CADE B S, NOON B R. A gentle introduction to quantile regression for ecologists[J]. Frontiers in Ecology and the Environment, 2003, 1(8): 412-420. DOI:10.1890/1540-9295(2003)001[0412:AGITQR]2.0.CO;2 |
| [31] |
李群峰. 基于分位数回归的面板数据模型估计方法[J]. 统计与决策, 2011(17): 24-26. LI Q F. Estimation method of panel data model based on Quantile regression[J]. Statistics & Decision, 2011(17): 24-26. |
| [32] |
焦璨, 王宣承, 张敏强, 等. 分位数回归:心理统计方法的重要补充[J]. 中国考试, 2009(1): 9-15. JIAO C, WANG X C, ZHANG M Q, et al. Quantile regression:an important supplement to the psychological statistics[J]. China Examinations, 2009(1): 9-15. |
| [33] |
逄志伟, 李显森, 朱建成, 等. 中东大西洋中部海域中上层鱼类中心渔场的时空变化[J]. 生态学杂志, 2016, 35(11): 3072-3079. PANG Z W, LI X S, ZHU J C, et al. Spatiotemporal patterns of central fishing ground of pelagic fishes in the sea area of central Eastern Central Atlantic[J]. Chinese Journal of Ecology, 2016, 35(11): 3072-3079. |
| [34] |
郑波, 陈新军, 李纲. GLM和GAM模型研究东黄海鲐资源渔场与环境因子的关系[J]. 水产学报, 2008, 32(3): 379-386. ZHENG B, CHEN X J, LI G. Relationship between the resource and fishing ground of mackerel and environmental factors based on GAM and GLM models in the East China Sea and Yellow Sea[J]. Journal of Fisheries of China, 2008, 32(3): 379-386. |
| [35] |
李纲, 陈新军. 夏季东海渔场鲐鱼产量与海洋环境因子的关系[J]. 海洋学研究, 2009, 27(1): 1-8. LI G, CHEN X J. Study on the relationship between catch of mackerel and environmental factors in the East China Sea in summer[J]. Journal of Marine Sciences, 2009, 27(1): 1-8. |
| [36] |
许永久, 张敏. 东南太平洋智利竹䇲鱼渔场浮游生物数量分布及与渔场关系[J]. 海洋渔业, 2007, 29(4): 289-295. XU Y J, ZHANG M. Distribution of plankton on Trachurus murphyi fishing grounds of the Southeast Pacific and its relationship with fishing grounds[J]. Marine Fisheries, 2007, 29(4): 289-295. |
| [37] |
HUSTON M A. Introductory Essay: critical issues for improving predictions[M]//SCOTT J M, HEGLUND P J, MORRISON M L. Predicting Species Occurrences: Issues of Accuracy and Scale. Covelo, CA: Island Press, 2002: 7-21.
|
2. National Engineering Research Center for Oceanic Fisheries, Shanghai 201306, China;
3. Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources, Ministry of Education, Shanghai 201306, China;
4. Collaborative Innovation Center for Distant-Water Fisheries, Shanghai 201306, China