 2020,
Vol. 29
2020,
Vol. 29



2. 上海海洋大学 上海水产养殖工程技术研究中心, 上海 201306;
3. 上海海洋大学 农业农村部鱼类营养与环境生态研究中心, 上海 201306
底栖动物指生活史全部或大部分时间生活于水体底部的水生动物类群,在淡水食物网中行使着各种功能,包括有机碎屑分解、物质矿化、水质净化及生物排泄和扰动引起的营养盐释放及泥水界面的物质交换等[1],因此是水生态系统的重要组成部分,并在其物质循环和能量流动中扮演重要作用。另外,底栖动物生活周期较长,营固定生活或活动空间有限,对水质的敏感性存在差异,它们的生长、繁殖以及群落结构与干扰(如污染程度)紧密相关[2],因而底栖动物是反映水体污染程度的良好指示类群[3]。
崇明岛为世界最大河口冲积、中国第3大岛,自2005年提出现代综合生态岛总体定位后,崇明生态岛建设逐渐升级,2016年12月进一步明确世界级生态岛建设定位。明珠湖位于崇明岛西南部绿华镇境内,是岛上最大的天然内陆湖泊。山水林田湖草是生命共同体,因此明珠湖在崇明世界级生态岛建设中扮演重要角色。2006—2009年,有学者[4-9]对明珠湖水生生物开展了调查,如浮游生物和底栖动物群落结构、多样性及水质生物学评价等方面。为了解近10年来该湖底栖动物的演变及其水质变化,2017—2018年,对明珠湖底栖动物进行了调查,分析了底栖动物群落结构现状及其与环境因子的关系,以期为明珠湖生态修复和水质管理提供基础数据和科学依据。
1 材料与方法 1.1 研究区域概况明珠湖(31°43.7′—31°45.0′N,121°14.8′—121°15.5′E)地处崇明岛西部的绿华镇境内,原为长江南支的一条支流,是20世纪70年代经人工筑堤围造成的泄湖,现建有2个进水闸门,属于通江湖泊。该湖南北宽约2.7 km、东西较窄约0.36~0.73 km,水体总面积仅约1.33 km2,平均水深约为2.7 m(1.0~4.5 m)。明珠湖地处亚热带季风气候区,气候温和湿润,四季分明,全年平均水温约为17.5 ℃(4.8~31.0 ℃)。
1.2 野外采样方法根据明珠湖的形态功能特征设置5个采样点(图 1),于2017年4月至2018年12月对明珠湖底栖动物进行逐月采样。采用改良彼得森采泥器(开口面积为0.062 5 m2)在每个站点采集底泥2次,将所采底泥倒入底栖生物过滤网(网目450 μm),在水中洗净污泥后将网中剩余物倒入500 mL塑料瓶中,带回实验室挑拣底栖生物样本,用8%的甲醛溶液固定,贴好标签以作进一步的种类鉴别、分类计数和样品湿质量称量(精确到0.000 1 g),最后依据采样面积折算成密度(ind./m2)和生物量(g/m2)。在底栖动物采样的同时,测定水深(WD)、水温(WT)、透明度(SD)、溶氧(DO)、pH、总氮(TN)、总磷(TP)和高锰酸盐指数(CODMn)等理化因子(表 1)。

|
图 1 崇明明珠湖底栖动物采样站点分布 Fig. 1 Sampling site distribution in the Mingzhu Lake of Chongming Island, Shanghai |

|
表 1 2017—2018年明珠湖主要理化因子的年均值 Tab.1 Annual average of the main physicochemical factors in Mingzhu Lake from 2017 to 2018 |
采用Margalef丰富度指数(R)、Shannon-Weaver多样性指数(H′,下文简称Shannon多样性指数)、Pielou均匀度指数(J)和Simpson优势度指数(λ)计算物种多样性指数,根据重要值(IV)判断优势种。
计算公式分别为
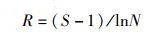 (1)
(1)
 (2)
(2)
 (3)
(3)
 (4)
(4)
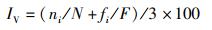 (5)
(5)
式中:S为物种数;fi为第i个物种的出现频次;F为所有物种出现频次的和;ni为第i个物种的个体数;N为所有物种的总个体数;IV为重要值。
1.4 水质生物学评价运用Wright指数、Carlander生物量指数、H′、J、Goodnight-Whitley指数(IGW)评价明珠湖水质,各指数计算公式及水质生物学评价标准如下。
Wright指数指每平方米颤蚓科的数量,评价标准:< 100为清洁;100~999为轻污染;1 000~5 000为中污染;>5 000为重污染[10]。
Carlander生物量的评价标准:底栖动物生物量处于0.2~1.7 g/m2为贫营养;2.50~6.25 g/m2为中营养;10.0~25.0 g/m2为富营养[11]。
利用两年调查数据计算每个站点的底栖动物物种数量组成,按1.3节中的公式计算Shannon多样性指数,并根据判断标准划分水质污染等级:H′为0~1是重污染;1~2是中污染;2~3是轻污染;H′>3为清洁[12]。
Pielou均匀度指数的水质评价标准:J < 0.3为重污染;0.3~0.5为中度污染;0.5~0.8为轻度污染;J >0.8为清洁状态[12]。
 (6)
(6)
式中:n为寡毛类的密度;N为各站点所有物种的密度和。水质评价标准:IGW < 30%为清洁;30%~60%为轻污染;60%~80%为中污染;IGW>80%为严重污染[13]。
1.5 统计分析数据处理及作图用Excel完成。在统计分析软件SPSS 19.0中,采用三因素方差分析(three-way ANOVA)探讨底栖动物密度、生物量和多样性的年际、季节和空间差异性。利用PRIMER 5.0软件进行聚类分析(CA)和非度量多维尺度分析(NMDS)以探讨明珠湖底栖动物物种组成的时空分布特征。分析之前,将相对密度低于1%的物种数据整合在一起,组成一组的变量(称作少见种)。将各季节和站点的生物密度换算为相对密度,然后对数据进行平方根反正弦函数转换,采用平均连接法和Bray-Curtis距离进行聚类分析。利用NMDS来验证结果是否准确,NMDS分析中采用Bray-Curtis距离。检验NMDS的拟合优度采用压力系数(stress),当stress≤0.1时,说明具有较好的拟合度;当stress<0.05时,说明该图形具有代表性。之后采用ANOSIM相似性分析检验不同群落之间差异的显著性,并应用SIMPER相似性百分比分析各物种在同一群落组内的平均相似性贡献率和不同群落组间的平均相异性贡献率[14]。
2 结果 2.1 底栖动物物种组成及其时空差异2017—2018年共在明珠湖采集到25种底栖动物,2017和2018年各有23和20种,均由线形动物门、节肢动物门、环节动物门、软体动物门组成。其中,节肢动物种类最多(12种),占总物种数的48.0%;其次为环节动物(7种、28.0%)和软体动物(5种,20.0%);线形动物只有1种(4.0%)。见表 2。

|
表 2 明珠湖大型底栖动物的重要值 Tab.2 Important values of different species in macrozoobenthic community in Mingzhu Lake |
整个调查期间,底栖动物优势种为霍甫水丝蚓(Limnodrilus hoffmeisteri)、中国长足摇蚊(Tanypus chinensis)和黄色羽摇蚊(Chironomus plumosus),分别占了群落总重要值的23.5%、21.6%和18.8%,前者在2年中均排在第1位,后2个物种均排在第2~3位。
聚类分析结果显示,明珠湖底栖动物在季节上无显著差异(ANOSIM,R=0.547,P>0.05),NMDS得出相同结果,其压力系数stress=0.12,说明群落划分无意义;在站点间也无显著差异(ANOSIM,R=0.588,P>0.05),NMDS得出相同结果,其压力系数stress=0.12,说明群落划分无意义。
2.2 底栖动物现存量及其时空变化明珠湖底栖动物的年均密度和生物量分别为1 116.8 ind./m2和3.53 g/m2。其中,节肢动物的现存量最大,密度和生物量分别为650.9 ind./m2和2.65 g/m2;密度环节动物门次之,软体动物(446.6 ind./m2)和线形动物(6.1 ind./m2)均很低;生物量软体动物(0.55 g/m2)和环节动物(0.32 g/m2)分列第2和第3位,线形动物最低(0.01 g/m2)。
密度较高的前3种底栖动物为霍甫水丝蚓(340.3 ind./m2,占年均总密度的30.5%)、中国长足摇蚊(303.7 ind./m2,27.2%)和黄色羽摇蚊(253.8 ind./m2,22.7%),3者密度占年均总密度的80.4%。生物量较高的前3种底栖动物为黄色羽摇蚊(1.91 g/m2,占年均总生物量的54.2%)、中国长足摇蚊(0.44 g/m2,12.6%)和霍甫水丝蚓(0.22 g/m2,6.1%),3者生物量占年均总生物量的72.8%。三因素方差分析表明,密度在年份、季节和站点间不存在显著差异,生物量仅存在显著的季节变化,不存在显著的年际和空间变化,这些因子之间对密度和生物量均不存在交互作用(表 3~6)。Duncan氏多重比较表明,秋季生物量显著高于冬季和夏季,春季显著高于夏季,其他季节两两间不存在显著差异(表 5)。

|
表 3 明珠湖底栖动物现存量及多样性指数三因素方差分析 Tab.3 Three-way analysis of variance of the standing crop and diversity indices of benthic animals in Mingzhu Lake |

|
表 4 明珠湖底栖动物现存量及多样性指数年际变化 Tab.4 Annual variation of the standing crop and diversity index of benthic animals in Mingzhu Lake |

|
表 5 明珠湖底栖动物现存量及多样性指数的季节变化 Tab.5 Seasonal variation of the standing crop and diversity indices of benthic animals in Mingzhu Lake |

|
表 6 明珠湖底栖动物现存量及多样性指数的空间变化 Tab.6 Spatial variation of the standing crop and diversity indices of benthic animals in Mingzhu Lake |
三因素方差分析表明,丰富度指数无显著的年际、季节和空间差异,Shannon多样性指数、均匀度指数和优势度指数存在显著的空间变化,无显著年际和季节差异,3因素对4种多样性指数均无交互作用(表 3~6)。
Duncan氏多重比较表明,S1站点的Shannon多样性指数和均匀度指数均显著低于其他4个站点,优势度指数正好相反,其他4个站点两两间无显著差异(表 6)。
2.4 现存量和多样性指数与环境因子的关系相关分析表明,影响现存量和多样性指数的主要环境因子为亚硝态氮、高锰酸盐指数和水深。密度和优势度指数与水深显著正相关,Shannon多样性指数、丰富度指数和均匀度指数与水深显著负相关。生物量和优势度指数与高锰酸盐指数显著正相关,Shannon多样性指数和丰富度指数与高锰酸盐指数显著负相关。生物量还与亚硝态氮显著正相关(表 7)。

|
表 7 现存量和多样性指数与环境因子的相关性分析 Tab.7 Correlation analysis between standing crop and diversity index and environmental factors |
采用Wright指数、Carlander生物量指数、Shannon多样性指数(H′)、Pielou均匀度指数(J)、Goodnight-Whitley指数5个指数对明珠湖进行水质评价。Wright指数评价结果显示S3、S5号站点为轻污染状态,S1、S2、S4号站点为中污染状态;Goodnight-Whitley指数评价的明珠湖S1、S2号站点为清洁状态,S3、S4、S5号站点为轻污染状态;从Carlander生物量指数评价的结果可以看出,5个站点均为中营养状态;通过Shannon多样性指数对水质进行评价,5个站点均为中污染状态;Pielou均匀度指数评价的结果表明,S1号站点为中污染状态,其他4个站点为轻污染状态(表 8)。

|
表 8 各指数对明珠湖水质的评价 Tab.8 Water quality assessment of Mingzhu Lake according to different indices |
大型底栖动物群落结构如物种组成、密度、生物量和多样性的时空分布主要与生境的异质性、底质类型、水深、水温以及营养水平有关[15-19]。2006—2008年明珠湖底栖动物物种组成、密度、生物量和各种多样性指数均无空间差异[9],与此一致,本研究结果显示该湖底栖动物密度、生物量和物种组成亦无空间差异,但除了丰富度指数之外的其他3种多样性指数均存在显著空间差异(表 6),这可能主要与站点水深存在较大差异有关。2006—2008年底栖动物调查期间,明珠湖旅游开发还处于起步阶段,仅建有简易码头。本研究之前,该湖旅游已经初具规模,建成了大型码头。为了大型游船停泊及通航的需要,对S1号站点附近水域进行疏浚,水深得以增加。本次调查发现,在5个站点中,S1号站点的水深(3.6 m)最大,显著大于S3(2.4 m)和S4(2.7 m)号站点,后两者又显著大于S2(1.5 m)和S5(1.6 m)号站点(双因素方差分析:季节,F=0.190,P=0.903>0.05;站点,F=48.003,P<0.001;季节×站点交互作用,F=1.148,P=0.337>0.05)。
水深会导致水体重要理化因子(如食物和溶解氧)发生变化而间接影响底栖动物,成为影响静水生态系统底栖动物分布的关键生态因子[17, 20]。深水湖库软体动物物种会随着水深的增加而减少甚至消失[20-21],有些浅水湖泊也有减少的趋势[22]。研究发现,水深最深的S1号站点的软体动物物种数只有1种,其他站点有2~4种。另外,水深的增加可能会导致耐受低氧等能力低的物种消失,霍甫水丝蚓等耐受能力强的物种的相对密度增加,从而增加该站点底栖动物群落的优势度、降低均匀度,从而使Shannon多样性指数下降。此推理与我们的研究结果一致,Shannon多样性指数和均匀度指数与水深显著负相关、优势度指数与水深显著正相关(表 7);S1号站点的Shannon多样性指数和均匀度指数均显著低于其他4个站点,但优势度指数相反(表 6)。另外,S1号站点水深的增加由疏浚导致,疏浚本身及其产生的扰动可能导致明珠湖S1号站点底栖动物多样性显著低于其他站点[23]。陈其羽等[24]发现,在水深不超过5 m的浅水湖泊中,水深每增加1 m,底栖动物密度减少330 ind./m2。本研究结果与此相反,明珠湖底栖动物密度与水深显著正相关,S1号站点水深最大,密度最高(表 6)。水深较大的地方可能有利于有机质沉降累积[8],明珠湖水体深的地方沉积物全部由淤泥组成,而比较浅的地方淤泥比例稍有下降,粗有机质含量有所增加。颤蚓亚科寡毛类和摇蚊幼虫更喜欢在有机质含量高的淤泥中生活[25]。
一些研究表明,底栖动物物种分布、现存量以及多样性等群落结构与化学需氧量(CODCr)或CODMn有关[26-29]。CODMn或CODCr是表征水体污染程度的重要综合指标之一,它们越高的地方污染越严重。随着营养盐(污染)梯度变化,底栖动物多样性和现存量呈单峰分布,在某一营养盐水平下达到最高值,低于或者高于这一阈值,多样性和现存量都会下降[30],东湖底栖动物多样性与营养水平呈负相关关系,而密度与之呈正相关关系[31]。明珠湖底栖动物Shannon多样性和丰富度指数与CODMn负相关,而生物量与之正相关(表 6),说明底栖动物现存量由升转降的营养盐阈值可能高于多样性转换的营养盐阈值。高剂量亚硝酸盐具有很大毒性,在明珠湖其含量很低(0.005 mg/L),远低于对水生生物产生损害的临界值0.1 mg/L[32]。亚硝酸盐能造成水体富营养化、可反映水质污染程度[32],明珠湖底栖动物生物量与亚硝酸盐显著正相关,说明亚硝酸盐表征的营养盐水平还未到达导致底栖动物现存量由升转降的阈值。磷含量是水体富营养化程度的一个重要指标,有学者[24, 33]指出总磷是影响底栖动物群落结构的主要环境因子,明珠湖2017—2018年总磷含量比2007—2008年增加了约50%,但总磷不是影响明珠湖底栖动物群落结构(如现存量空间分布)的环境因子,其他报道也存在此类现象[15, 26],可能其他一些环境因子起着更为重要的作用。
2006—2008年,红裸须摇蚊(Propsilocerus akamusi)为明珠湖摇蚊幼虫群落和底栖动物群落的优势种,且是造成群落物种组成季节差异的指示物种,冬季密度很高、而其他季节很低或为0[8-9]。红裸须摇蚊幼虫为冷水性种类,具季节性垂直迁移特性,在水温低于20 ℃时,将潜入底泥深处滞育,深度可达80~90 cm[34]。本研究发现,明珠湖底栖动物群落物种组成无显著季节差异,主要与本次调查红裸须摇蚊密度在冬季很低有关。两次调查水温差别不是很大,推测不是水温增加导致其往沉积物深处迁移而不能被采泥器采集到所致,是否与物种竞争有关或仅反映了该物种种群数量的年际变化,有待进一步研究。
3.2 基于底栖动物的水质生物学评价与2006—2008年的调查结果相比,明珠湖溶解氧变化很小,透明度(-16.4%)、高锰酸盐指数(-25.1%)和总氮(-45.1%)有较大程度的下降,总磷有较大幅度的增加(50%),见表 1。但根据单因子水质评价,基于溶解氧和总磷的水质等级无变化,分别为Ⅰ类和Ⅴ类水,基于总氮和高锰酸盐指数的水质分别提升了1个等级,总氮和高锰酸盐指数分别显示明珠湖水质从Ⅳ类提升到Ⅲ类和从Ⅲ类提升到Ⅱ类。因此从水化学水质评价来看,2017—2018年水质较2006—2008年稍有改善,但仍属于Ⅴ类水(表 1)。多种底栖动物多样性和生物指数被用来评价水质,但很难取得一致性的评价结果[35]。指数的选择对水质生物学评价结果与实际污染现状的符合程度具有很大的影响。HU等[9]曾比较了多种生物指数和多样性指数对明珠湖水质评价的结果,提出采用香农多样性指数能较准确地对该湖的水质进行评价。本文研究发现,该指数评价的水质等级与2006—2008年的相比无变化,各站点均处于中污染状态,与水化学方法评价结果反映的情况较为类似,说明Shannon多样性指数能较为真实地反映该湖水质的长期变化。
本文结果表明,明珠湖底栖动物优势种为霍甫水丝蚓、中国长足摇蚊和黄色羽摇蚊,它们的密度也均处在前3位。霍甫水丝蚓和中国长足摇蚊为耐污染物种,在富营养化水体中普遍分布[15-16, 31, 36],羽摇蚊群物种也是有机污染较为严重水体的指示类群[15-16, 36]。3种耐污种占优势指示明珠湖水体受到了较为严重的有机污染,说明指示物种法(indicator species method)与水化学评价及Shannon多样性评价结果较为一致。
| [1] |
COVICH A P, PALMER M A, CROWL T A. The role of benthic invertebrate species in freshwater ecosystems[J]. BioScience, 1999, 49(2): 119-127. DOI:10.2307/1313537 |
| [2] |
王备新, 杨莲芳. 大型底栖无脊椎动物水质快速生物评价的研究进展[J]. 南京农业大学学报, 2001, 24(4): 107-111. WANG B X, YANG L F. Advances in rapid bio-assessment of water quality using benthic macroinvertebrates[J]. Journal of Nanjing Agricultural University, 2001, 24(4): 107-111. |
| [3] |
程宏, 王金辉, 唐娅菲, 等. 春季闽北海域大型底栖动物群落特征及环境质量分析[J]. 上海海洋大学学报, 2018, 27(2): 238-249. CHENG H, WANG J H, TANG Y F, et al. Research on community characteristics of macrozoobenthos and environmental quality of offshore North Fujianin spring[J]. Journal of Shanghai Ocean University, 2018, 27(2): 238-249. |
| [4] |
陈立婧, 吴竹臣, 胡忠军, 等. 上海崇明岛明珠湖浮游植物群落结构[J]. 应用生态学报, 2011, 22(6): 1599-1605. CHEN L J, WU Z C, HU Z J, et al. Phytoplankton community structure in Mingzhu Lake of Chongming Island, Shanghai[J]. Chinese Journal of Applied Ecology, 2011, 22(6): 1599-1605. |
| [5] |
吴竹臣.上海崇明明珠湖浮游植物群落结构演替及水质评价[D].上海: 上海海洋大学, 2011. WU Z C. Phytoplankton community structure succession and water quality evaluation in Mingzhu Lake, Chongming, Shanghai[D]. Shanghai: Shanghai Ocean University, 2011. http://cdmd.cnki.com.cn/Article/CDMD-10264-1012003637.htm |
| [6] |
陈立婧, 顾静, 彭自然, 等. 上海崇明岛明珠湖轮虫群落结构[J]. 应用生态学报, 2009, 20(12): 3057-3062. CHEN L J, GU J, PENG Z R, et al. Community structure of rotifer in Mingzhu Lake of Chongming Island, Shanghai[J]. Chinese Journal of Applied Ecology, 2009, 20(12): 3057-3062. |
| [7] |
陈立婧, 顾静, 胡忠军, 等. 上海崇明明珠湖原生动物的群落结构[J]. 水产学报, 2010, 34(9): 1404-1413. CHEN L J, GU J, HU Z J, et al. The research of protozoa community structure in Mingzhu Lake of Chongming Island, Shanghai[J]. Journal of Fisheries of China, 2010, 34(9): 1404-1413. |
| [8] |
胡忠军, 刘其根, 陈立婧, 等. 上海崇明明珠湖摇蚊幼虫群落结构特征及其对水质的指示作用[J]. 应用生态学报, 2009, 20(4): 929-936. HU Z J, LIU Q G, CHEN L J, et al. Structural characteristics of chironomid community and their indicative significance in bioassessment of water quality in Mingzhu Lake of Chongming Island, Shanghai[J]. Chinese Journal of Applied Ecology, 2009, 20(4): 929-936. |
| [9] |
HU Z J, JIAX X, CHEN X H, et al. Spatial and seasonal pattern of macrozoobenthic assemblages and the congruence in water quality bioassessment using different taxa in artificial Mingzhu Lake in Shanghai[J]. Chinese Journal of Oceanology and Limnology, 2016, 34(5): 928-936. DOI:10.1007/s00343-016-4244-9 |
| [10] |
WRIGHT S, TIFFANY L H, TIDD W M. Limnological survey of western Lake Erie[R]. U. S. Fish and Wildlife Service, Special Scientific Report: Fisheries, 1955, No. 139. https://www.researchgate.net/publication/284885976_Limnological_survey_of_western_Lake_Erie
|
| [11] |
CARLANDER K D. Farm fish pond research in Iowa[J]. Journal of Wildlife Management, 1952, 16: 258-261. DOI:10.2307/3796634 |
| [12] |
陈萍萍, 张瑞雷, 赵风斌, 等. 上海市河道底栖动物群落结构及其水质评价[J]. 上海海洋大学学报, 2014, 23(4): 564-572. CHEN P P, ZHANG R L, ZHAO F B, et al. The zoobenthos community structure and bio-assessment of water quality in Shanghai urban rivers[J]. Journal of Shanghai Fisheries University, 2014, 23(4): 564-572. |
| [13] |
GOODNIGHT C J, WHITLEY L S. Oligochaetes as indicators of pollution[C]. Proceeding of 15th annual water conference, vol 160, Purdue University, Lafayette, 1960: 139-142. https://www.researchgate.net/publication/284313110_Oligochaetes_as_Indicators_of_Pollution
|
| [14] |
LI Y L, LIU Q G, CHEN L P, et al. A comparison between benthic gillnet and bottom trawl forassessing fish assemblages in a shallow eutrophic lake near the Changjiang River estuary[J]. Journal of Oceanology and Limnology, 2018, 36(2): 572-585. DOI:10.1007/s00343-018-6219-5 |
| [15] |
张超文, 张堂林, 朱挺兵, 等. 洪泽湖大型底栖动物群落结构及其与环境因子的关系[J]. 水生态学杂志, 2012, 33(3): 27-33. ZHANG C W, ZHANG T L, ZHU T B, et al. Community structure of macrozoobenthos and its relationship with environmental factors in Lake Hongze[J]. Journal of Hydroecology, 2012, 33(3): 27-33. |
| [16] |
章海鑫, 付辉云, 张燕萍, 等. 太泊湖底栖动物种群结构特征及水质评价[J]. 上海海洋大学学报, 2016, 25(3): 431-437. ZHANG H X, FU H Y, ZHANG Y P, et al. The community structure of benthic animals and water quality assessment in Taibo Lake[J]. Journal of Shanghai Fisheries University, 2016, 25(3): 431-437. |
| [17] |
PETRIDIS D, SINIS A. Benthos of Lake MikriPrespa (North Greece)[J]. Hydrobiologia, 1995, 304(3): 185-196. DOI:10.1007/BF02329313 |
| [18] |
BECHARA J A. The relative importance of water quality, sediment composition and floating vegetation in explaining the macrobenthic community structure of floodplain lakes (Parana River, Argentina)[J]. Hydrobiologia, 1996, 333(2): 95-109. DOI:10.1007/BF00017572 |
| [19] |
CHEN L P, ZHANG Y, LIU Q G, et al. Spatial variations of macrozoobenthos and sediment nutrients in Lake Yangcheng:Emphasis on effect of pen culture of Chinese mitten crab[J]. Journal of Environmental Sciences, 2015, 37(11): 118-129. |
| [20] |
胡忠军, 孙月娟, 刘其根, 等. 浙江千岛湖深水区大型底栖动物时空变化格局[J]. 湖泊科学, 2010, 22(2): 265-271. HU Z J, SUN Y J, LIU Q G, et al. Temporal and spatial distribution of profundal macrozoobenthic community in Lake Qiandao, Zhejiang, China[J]. Journal of Lake Science, 2010, 22(2): 265-271. |
| [21] |
CUI Y D, WANG H Z. Ecology of macrozoobenthic communities in two plateau lakes of Southwest China[J]. Chinese Journal of Oceanology and Limnology, 2008, 26(4): 345-352. DOI:10.1007/s00343-008-0345-4 |
| [22] |
孙月娟, 王武, 刘其根, 等. 江苏阳澄湖螺类群落的空间分布格局[J]. 长江流域资源与环境, 2011, 20(6): 711-716. SUN Y J, WANG W, LIU Q G, et al. Spatial Distribution Pattern of Snail Community in Yangcheng Lake, Jiangsu Province[J]. Resources and Environment in the Yangtza Basin, 2011, 20(6): 711-716. |
| [23] |
姜苹红, 周易勇, 纪磊, 等. 月湖底泥疏浚后底栖动物群落的恢复及其与环境的关系[J]. 生态环境学报, 2012, 21(6): 1088-1095. JIANG P H, ZHOU Y Y, JI L, et al. Zoobenthos recovering process and the relations with environment after sediment dredging in Lake Yuehu[J]. Ecology and Environmnet, 2012, 21(6): 1088-1095. |
| [24] |
陈其羽, 梁彦龄, 吴天惠. 武汉东湖底栖动物群落结构和动态的研究[J]. 水生生物学集刊, 1980, 7(1): 41-56. CHEN Q Y, LIANG Y L, WU T H. Study on the structure and dynamics of zoobenthos community in East Lake, Wuhan[J]. Acta Hydrobiologica Sinica, 1980, 7(1): 41-56. |
| [25] |
MATISOFF G, WANG X S, MCCALL P L. Biological redistribution of lake sediments by tubificid oligochaetes:Branchiuras owerbyi and Limnodrilus hoffmeisteri/Tubifex tubifex[J]. Journal of Great Lakes Research, 1999, 25(1): 205-219. DOI:10.1016/S0380-1330(99)70729-X |
| [26] |
高欣, 牛翠娟, 胡忠军. 太湖流域大型底栖动物群落结构及其与环境因子的关系[J]. 应用生态学报, 2011, 22(12): 3329-3336. GAO X, NIU C J, HU Z J. Macrobenthos community structure and its relations with environmental factors in Taihu River basin[J]. Chinese Journal of Applied Ecology, 2011, 22(12): 3329-3336. |
| [27] |
李佳娟, 于洪贤, 刘曼红, 等. 西泉眼水库水源地10月大型底栖动物群落特征及其与环境因子的关系[J]. 东北林业大学学报, 2011, 39(11): 79-83. LI J J, YU H X, LIU M H, et al. Characteristics of macrobenthos communities in Xiquanyan Reservoir in Autumn and their relationships with environmental factors[J]. Journal of Northeast Forestry University, 2011, 39(11): 79-83. |
| [28] |
刘祥, 陈凯, 陈求稳, 等. 淮河流域典型河流夏秋季底栖动物群落特征及其与环境因子的关系[J]. 环境科学学报, 2016, 36(6): 1928-1938. LIU X, CHEN K, CHEN Q W, et al. The community structure of macroinvertebrate and its relationship to the environmental factors in summer and autumn within typical reaches of Huai River Basin[J]. Acta Scientiae Circumstantiae, 2016, 36(6): 1928-1938. |
| [29] |
张蕾, 于海燕, 黄旻, 等. 浦阳江底栖动物群落特征与水质生物评价[J]. 应用与环境生物学报, 2018, 24(1): 155-161. ZHANG L, YU H Y, HUANG M, et al. Characteristics of zoobenthos community and biological evaluation of water quality in Puyang River[J]. Chinese Journal of Applied and Environmental Biology, 2018, 24(1): 155-161. |
| [30] |
FROUIN P. Effects of anthropogenic disturbances of tropical soft-bottom benthic communities[J]. Marine Ecology Progress, 2000, 194(1): 39-53. |
| [31] |
龚志军, 谢平, 唐汇涓, 等. 水体富营养化对大型底栖动物群落结构及多样性的影响[J]. 水生生物学报, 2001, 25(3): 210-216. GONG Z J, XIE P, TANG H J, et al. The influence of eutrophication upon community structure and biodiversity of macrozoobenthos[J]. Acta Hydrobiologica Sinica, 2001, 25(3): 210-216. |
| [32] |
周超, 高乃云, 楚文海, 等. 水体中亚硝酸盐生物毒性和去除的研究进展[J]. 给水排水, 2011, 37(5): 104-108. ZHOU C, GAO N Y, CHU W H, et al. Progress of study on biotoxicity and removal of nitrite in water[J]. Water & Wastewater Engineering, 2011, 37(5): 104-108. |
| [33] |
王小毛, 欧伏平, 王丑明, 等. 洞庭湖底栖动物长期演变特征及影响因素分析[J]. 农业环境科学学报, 2016, 35(2): 336-345. WANG X M, OU F P, WANG C M, et al. Long-term evolution and influencing factors of macrozoobenthos in Dongting Lake[J]. Journal of Agro-Environment Science, 2016, 35(2): 336-345. |
| [34] |
郭先武. 武汉南湖三种摇蚊幼虫生物学特性及其种群变动的研究[J]. 湖泊科学, 1995, 7(3): 249-255. GUO X W. Study on biological characteristics and population changes of three kinds of chironomidlarvae in Nanhu Lake, Wuhan[J]. Journal of Lake Science, 1995, 7(3): 249-255. |
| [35] |
HU Z J, WU H, LIU Q G. The ecology of zoobenthos in reservoirs of China: a mini-review[M]. Tropical and Sub-Tropical Reservoir Limnology in China, 2012: 155-164.
|
| [36] |
姜苹红, 崔永德, 王海军, 等. 汉阳湖群底栖动物群落及其对环境质量的指示[J]. 长江流域资源与环境, 2011, 20(5): 525-533. JIANG P H, CUI Y D, WANG H J, et al. Structural characteristics of macrozoobenthos community and their indicative significance in bioassessment of water quality in Hanyang Lake[J]. Resources and Environment in the Yangtza Basin, 2011, 20(5): 525-533. |
2. Shanghai Engineering Research Center of Aquaculture, Shanghai Ocean University, Shanghai 201306, China;
3. Centre for Research on Environmental Ecology and Fish Nutrition, Ministry of Agriculture and Rural Affairs, Shanghai Ocean University, Shanghai 201306, China