 2019,
Vol. 28
2019,
Vol. 28



河口区及其邻近地区因生态环境的敏感性和脆弱性,以及在生态系统中的重要性,一直都是研究的热点区域[1-3]。杭州湾营养物质丰富,饵料生物繁盛,是多种海洋生物产卵、索饵和育肥的场所[4]。杭州湾东北部面积较大且渔业资源丰富,为渔业资源的开发利用提供了有利条件。
近年来,关于定置张网渔业资源的研究较多集中在长江口常熟段及闽南海区和舟山沿岸。如张敏莹等[5]对常熟段定置张网渔业资源组成及多样性的调查分析,表明水利枢纽和泄洪排涝等对渔业资源有负面影响。刘勇等[6]对闽南海区定置张网渔业资源的研究及张龙等[7]对舟山沿岸定置张网渔业资源的研究,表明休渔期能够养护部分经济物种。目前对杭州湾东北部定置张网渔业资源的研究相对较少,仅王淼等[8]对杭州湾金山卫渔业资源种类和多样性进行了分析,因此对杭州湾东北部渔业资源群落结构的季节变化分析对种质资源保护、可持续利用和生态系统修复等具有重要意义。
1 材料与方法 1.1 数据来源本文所用数据来自2017年5月、8月、10月在杭州湾东北部所获得的定置张网渔业资源资料,每个月采集两次,每次采集1天(张网网衣总长22 m,网口扩张高度3.4 m,网口扩张长度13.6 m,网口周长34 m,网囊目13~15 mm)。由于杭州湾东北部渔业捕捞的现状和海况特点,因此将调查范围设置在30°00′~31°30′N、121°00′~122°24′E,张网站位设置见图 1。样品保存、分类、计数及称量均参照《海洋渔业资源调查规范》(SC/T 9403—2012)的要求操作。渔业资源的种类鉴定及生态类群划分参考《东海经济虾蟹类》[9]和《浙江动物志》(甲壳类) [10]及《世界鱼类》[11]。

|
图 1 杭州湾东北部采样站点分布示意图 Fig. 1 Sampling area in North-eastern Hangzhou Bay |
采用Jaccard相似性系数[12]、Shannon-Wiener多样性指数[13]、Margalef丰富度指数[14]、Pielou均匀度指数[15]、相对重要性指数IRI[16]来分析杭州湾定置张网渔业资源群落结构多样性、物种相似性及优势种的组成。
Jaccard相似性系数:
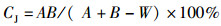 (1)
(1)
Shannon-Wiener多样性指数:
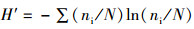 (2)
(2)
Margalef丰富度指数:
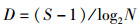 (3)
(3)
Pielou均匀度指数:
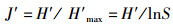 (4)
(4)
相对重要性指数:
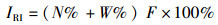 (5)
(5)
式中:ni为样品中第i种的个体数;N为所有种类的总个体数;S为种类总数;A、B分别为不同季度出现的物种数;W为两时期出现的共有物种数;N%和W%分别代表每个物种占总捕捞量的个体质量百分比和个体数量百分比;F%为某一种类渔业资源的出现频率的百分比。CJ为0~25%,为极不相似;25%~50%为中等不相似;50%~75%为中等相似;75%~100%为极相似[17]。0<D<1, 被认为受到重度影响;1<D<3,被认为受到中度影响。根据相对重要性指数(IRI) [16]分析研究渔业资源中的优势种及常见种成分,将相对重要性指数(IRI)大于1 000的定为优势种,100~1 000的定为常见种。通过SIMPER(species contributions to similarity)了解表征群落特征的物种(累积贡献率≥5%),用PRIMER软件计算得出[18]。
采用Primer 5.0软件,在Bray-Curtis相似性度量的基础上进行等级聚类(UPGMA)和非度量多维尺度(non-metric multidimensional scaling, MDS),分析杭州湾东北部渔业资源群落结构的相似性程度,MDS分析用胁迫系数(Stress)来衡量二维点图的优劣:当0.1<Stress<0.2时,具有一定的解释意义;当0.05<Stress<0.1时,排序效果基本可信;当Stress<0.05时,二维点图对群落结构排序具有很好的代表性[19-20]。并绘制丰度/生物量比较曲线(abundance /biomass curves,ABC曲线),根据ABC曲线中生物量和丰度的K-优势度曲线的波动,分析渔业资源群落在不同干扰状况下的特征[21]。
2 结果 2.1 种类组成及相似性春夏秋3个季度的定置张网渔业资源调查,共捕获渔业资源48种(表 1),其中,春季定置张网共捕获28种,夏季定置张网共捕获34种,秋季定置张网共捕获30种,春夏季共有种21种,夏秋季共有种21种,春秋季共有种19种,3季度共有种18种。物种相似性分析表明,春夏两季度的Jaccard相似性系数仅为23.22%,夏秋两季度的Jaccard相似性系数为23.72%,春秋两季度的Jaccard相似性系数为21.54%,均为极不相似水平。因此,不同季度杭州湾东北部定置张网的渔业资源物种相似性水平较低。

|
表 1 不同季度杭州湾东北部渔业资源的种类组成 Tab.1 Species composition of fishery resources in different seasons in North-eastern Hangzhou Bay |
春夏秋3个季度共捕获渔业资源48种(表 1),其中,鱼类30种,虾蟹类16种,水母类2种。
根据不同鱼类生长和繁殖的适温性及生态习性将鱼类主要分为以下5个生态类群。
(1) 河口暖水种:这种类型的鱼类包括河口定居性种类、降海洄游和溯河洄游的种类,大多数属于广温广盐性或广温低盐性种类。本次3个季度调查所获鱼类中共有8种,即刀鲚(Coilia nasua)、凤鲚(Coilia mystus)、大弹涂鱼(Boleophthalmus pectinirostris)、中国花鲈(Lateolabrax maculatus)、四指马鲅(Eleutheronema tetradactylum)、棱
(2) 近海暖水种:这种类型的鱼类多数栖息分布在水深30 m以上,因为具有较强的适温适盐能力,因此多数为广温广盐性种类。共有7种,即小带鱼(Eupleurogrammus muticus)、海鳗(Muraenesox cinereus)、
(3) 近海暖温种:共有7种,即黄姑鱼(Nibea albiflora)、黄鲫(Setipinna taty)、蓝点马鲛(Scombermorus niphonius)、黄鳍东方鲀(Takifugu xanthopterus)、小黄鱼(Pseudosciaena polyactis)、银鲳(Pampus argenteus)、黑鳃梅童鱼(Collichthy sniveatus)。
(4) 沿岸暖水种:这一类型的鱼类多数是在每年的春夏季度从近海或较深海区洄游至沿岸浅水区进行生殖产卵, 幼体在产卵场附近进行索饵、生长发育, 到了秋末冬初, 随着水温的下降而向较深海区进行越冬洄游, 这是一群广温广盐性种类。共有5种,即龙头鱼(Harpodon nehereus)、拉氏狼牙虾虎鱼(Odontamblyopus lacepedii)、皮氏叫姑鱼(Johnius belengerii)、孔虾虎鱼(Trypauchen vagina)、鲬鱼(Platycephalus indicus)。
(5) 沿岸暖温种:共有2种,即焦氏舌鳎(Cynoglossus joyneri)和安氏新银鱼(Neosalanx anderssoni)。
根据不同虾蟹类的分布水深,水温及盐度将虾蟹类主要分为以下2个生态类群。
(1) 广温低盐种:共有4种,即安氏白虾(Exopalaemon annandalei)、脊尾白虾(Exopalaemon carinicauda)、巨指长臂虾(Palaemon macrodactylus)、刺螯鼓虾(Alpheus hoplocheles)。
(2) 广温广盐种:共有11种,即葛氏长臂虾(Palaemon gravieri)、哈氏仿对虾(Parapenaeopsis hardwickii)、周氏新对虾(Metapenaeus joyneri)、太平洋长臂虾(Palaemon pacificus)、细指长臂虾(Palaemon tenuidactylus)、中国毛虾(Aceteschinensis)、口虾蛄(Oratosquilla oratoria)、三疣梭子蟹(Portunus trituberculatus)、日本
分析认为,凤鲚及安氏白虾为3个季度的共同优势种,其他优势种及常见种随着季度变化而发生改变。春季的优势种有3种:小黄鱼、凤鲚、安氏白虾;常见种有3种:狼牙鳗虾虎鱼、葛氏长臂虾、三疣梭子蟹;夏季的优势种有5种:凤鲚、安氏白虾、龙头鱼、棘头梅童鱼和葛氏长臂虾;常见种有2种:黄鳍东方鲀和灰鲳;秋季的优势种有3种:凤鲚、安氏白虾、龙头鱼;常见种有4种:棘头梅童鱼、狭颚绒螯蟹、三疣梭子蟹和葛氏长臂虾。

|
表 2 不同季度杭州湾东北部渔业资源的优势种 Tab.2 Dominant species of the fishery resources in different seasons in North-eastern Hangzhou Bay |
分析认为,凤鲚和安氏白虾为3个季度共有的特征种,春季渔业资源群落以小黄鱼、凤鲚和安氏白虾为特征种,夏季以凤鲚和安氏白虾、葛氏长臂虾为特征种,秋季以凤鲚、安氏白虾、葛氏长臂虾、龙头鱼为特征种(表 3)。

|
表 3 不同季度杭州湾东北部渔业资源的特征种 Tab.3 Characteristic species of fishery resources in different seasons in North-eastern Hangzhou Bay |
特征种共有6种(鱼类4种、虾蟹类2种),从贡献率表明:春季的杭州湾东北部渔业资源中鱼类的贡献率相对较高,表明鱼类在杭州湾东北部渔业资源群落中具有重要作用,能在一定程度上反映整个渔业资源群落结构的特点;夏秋季的杭州湾东北部渔业资源群落中虾蟹类具有绝对优势, 其代表了整个群落的特征。3个季度中,夏秋季度的特征种变化相对较小,而春夏两季的特征种变化相对比较明显,表明杭州湾东北部渔业资源群落在夏季和秋季较为稳定,而春夏季变化相对较大,反映出春夏季是杭州湾定置张网群落结构变动较大的季度,而夏秋季定置张网群落结构变动相对较小且较为稳定。
2.5 多样性指数分析分析认为,3个季度杭州湾东北部渔业资源群落Margalef丰富度指数均在1~3之间,因此受到中度影响。Shannon-Wiener多样性指数秋季最高,夏季最低,表现为秋季>春季>夏季;均匀度指数的最高值出现在秋季,最低值出现在夏季,表现为秋季>春季>夏季。总体而言,秋季相对其他季度能维持更高的多样性(表 4)。

|
表 4 不同季度杭州湾东北部渔业资源的多样性 Tab.4 Diversity of fishery resource sin different seasons in North-eastern Hangzhou Bay |
根据群落等级聚类结果,在相似性47.20%时,可将渔业资源群落结构聚为2组,第1组为春季的A1~A6号站位,第2组为夏季的B1~B6号站位和秋季的C1~C6号站位;根据MDS分析结果,可以将渔业资源群落分为2组,春季的A1~A6号站位为1组,夏季的B1~B6号站位和秋季的C1~C6号站位为第2组,Stress值为0.12在0.1~0.2之间,所以MDS结果具有一定的解释意义。因此,群落等级聚类和MDS排序结果均能说明由于季度变化,春季和夏秋季之间出现显著的群落结构差异,群落特点发生改变(图 2)。

|
图 2 不同季度杭州湾东北部渔业资源的聚类分析和MDS分析 Fig. 2 Dendrogram of cluster and MDS analysis of fishery resources in different seasons in North-eastern Hangzhou Bay |
春季的丰度曲线部分位于生物量曲线上方,且部分有所重叠,表明春季杭州湾东北部渔业资源群落结构受到了中等程度;夏、秋季的丰度/生物量曲线有重叠,生物量曲线部分位于丰度曲线上方且重叠程度低于春季,表明夏、秋季杭州湾东北部渔业群落结构受到了中等程度或重度的干扰,且受干扰程度高于春季,群落结构相对春季不稳定(图 3)。

|
图 3 不同季度杭州湾东北部渔业资源的ABC曲线及W统计值 Fig. 3 ABC curves and W value of fishery resources in different seasons in Northeastern Hangzhou Bay |
本次调查共捕获渔业资源48种(鱼类30种、虾蟹类16种)。3个季度渔业资源的种类数量和组成均不相同,共有鱼类6种、虾蟹类10种,与庞敏等[22]在2012—2013年对杭州湾海域虾蟹类的调查结果相比共有相同虾蟹类8种、不同虾蟹类8种,出现差异的原因可能其研究的海域受到钱塘江径流影响相对明显,且盐度明显低于杭州湾东北部[20];与谢旭等[23]在2012年对杭州湾鱼类调查的结果相比共有相同鱼类12种、不同鱼类18种,从生态习性上分析本次调查河口性种类和沿岸性种类共有15种占总数的一半,而谢旭等[23]的研究中河口性种类和沿岸性种类占绝对优势,造成差异的主要原因可能是谢旭等[23]的研究区域处于钱塘江口的典型的河口区,而本次调查的区域在杭州湾的东北部;与王淼等[24]在2016年对杭州湾东北部渔业资源调查结果相比共有相同渔业资源种类26种、不同渔业资源种类22种,出现差异的主要原因可能是王淼等[24]调查所用的渔具为拖网,最小囊网目20 mm,采样时间仅为1 h,而本调查采用定置张网,网目尺寸相对较小, 仅为13~15 mm且采样时间为1 d,对渔业资源的选择性差,因此捕获的渔业资源种类和数量相对较多。
3.2 杭州湾东北部定置张网优势种的季节变化春季,杭州湾东北部定置张网共捕获鱼类14种、虾蟹类13种,优势种中的鱼类共有2种,包括暖水性的凤鲚及暖温性的小黄鱼,其中凤鲚的相对重要性指数最高,为第一优势种,研究表明凤鲚初春时节游向近岸,在杭州湾及长江口附近进行生殖洄游[25-26],产卵后亲体游向深水区,幼鱼在江河口和近海索饵成长,春季出现大量的凤鲚与鱼类洄游存在一定的联系。虾蟹类优势种仅有1种为广温低盐的安氏白虾。
夏季,杭州湾东北部定置张网共捕获鱼类22种、虾蟹类12种,随着海水温度的升高,物种大量更替,暖水种大量出现,所占比例迅速上升,优势种中的鱼类包括3种,分别是暖水性的龙头鱼、棘头梅童鱼及凤鲚,优势种中的虾蟹类包括2种分别是广温低盐的安氏白虾及广温广盐的葛氏长臂虾,安氏白虾为第一优势种,且尾数和质量均远远高于其他物种,这可能是因为安氏白虾在杭州湾东北部有广泛分布,生长快,世代更新迅速,对环境适应能力强[27]且本文调查数据显示在夏季定置张网中安氏白虾的抱卵比例高。秋季,杭州湾东北部定置张网共捕获鱼类16种、虾蟹类12种,优势种中的鱼类包括2种,分别为暖水性的凤鲚和龙头鱼,优势种中的虾蟹类仅有1种,为广温低盐的安氏白虾,安氏白虾为第一优势种,但秋季出现的安氏白虾幼体较多,且生物量/丰度的比例均低于春夏季,造成这样的结果可能是因为安氏白虾4月中旬进入繁殖期,以抱卵亲虾为主,而9月份则以当年生的安氏白虾占绝对优势,其亲本基本消失[22]。
3.3 群落结构受扰动状况分析杭州湾定置张网渔业资源的ABC曲线显示,3个季度的丰度优势度曲线均在生物量优势度曲线之上,且丰度优势度曲线的起点均明显高于生物量优势度曲线的起点,这说明杭州湾东北部定置张网渔业资源的优势种主要由个体小、数量大的种类组成,群落结构处于中度及以上的干扰状态。从W统计值看,秋季W绝对值最高,其次为夏季,说明夏季和秋季杭州湾东北部定置张网渔业资源的群落受扰动的程度相对较高,这与群落结构等级聚类分析和MDS分析的结果相一致。部分原因可能是季节之间的环境变化如海水温度提高、盐度改变[29]等。另外近年来的大型海洋工程建设如东海大桥海上风电场、洋山深水港等[29]导致杭州湾海洋生态环境受到破坏,对环境造成一定影响的同时加大了海洋环境压力,特别是海洋工程的建设直接影响了底栖动物的生存环境[29],降低了鱼类和虾蟹类的多样性和数量密度[30],使渔业资源的群落结构向小型及低龄化过度,生态失衡已严重影响了海洋生态系统的健康和生态服务功能的正常发挥,进而影响渔业资源群落的受扰动程度。杭州湾东北部为传统定置张网作业区,该渔业资源群落结构受到人为扰动的影响较大。张网作业对渔业资源的选择性差,使得捕捞小型经济鱼、虾蟹类的同时兼捕到大量的经济鱼类幼鱼及虾蟹类幼苗。夏秋季中安氏白虾栖息密度占据绝对优势的原因可能是其广泛分布,世代更替快[22]。建议今后加强对杭州湾东北部的监测和生态调查,从而掌握其水文环境及生物资源的动态变化和影响机制,为杭州湾东北部修复生态环境和生物资源的可持续利用提供科学依据。
| [1] |
李宝泉, 李新正, 王洪法, 等. 长江口附近海域大型底栖动物群落特征[J]. 动物学报, 2007, 53(1): 76-82. LI B Q, LI X Z, WANG H F, et al. Characters of a macrobenthic community off the Changjiang River Estuary[J]. Acta Zoologica Sinica, 2007, 53(1): 76-82. |
| [2] |
RITTER C, MONTAGNA P A, APPLEBAUM S. Short-term succession dynamics of macrobenthos in a salinity-stressed estuary[J]. Journal of Experimental Marine Biology and Ecology, 2005, 323(1): 57-69. DOI:10.1016/j.jembe.2005.02.018 |
| [3] |
DESROY N, JANSON A L, DENIS L, et al. The intra-annual variability of soft-bottom macrobenthos abundance patterns in the North Channel of the Seine Estuary[J]. Hydrobiologia, 2007, 588(1): 173-188. DOI:10.1007/s10750-007-0661-2 |
| [4] |
罗民波, 陆健健, 沈新强, 等. 大型海洋工程对洋山岛周围海域大型底栖动物生态分布的影响[J]. 农业环境科学学报, 2007, 26(1): 97-102. LUO M B, LUJ J, SHEN X Q, et al. Impact of huge ocean engineering on distribution characteristics of macrobenthos in waters around Yangshan islands in Hangzhou Bay[J]. Journal of Agro-Environment Science, 2007, 26(1): 97-102. DOI:10.3321/j.issn:1672-2043.2007.01.018 |
| [5] |
张敏莹, 刘凯, 段金荣, 等. 长江常熟段定置张网渔获物组成及多样性季节变化分析[J]. 中国农学通报, 2011, 27(26): 92-96. ZHANG M Y, LIU K, DUAN J R, et al. Study on catch composition and diversity seasonal change of set net in Changshu section of the Yangtze River[J]. Chinese Agricultural Science Bulletin, 2011, 27(26): 92-96. |
| [6] |
刘勇, 洪明进, 叶泉土. 闽南海区休渔前后张网作业渔获物组成比较分析[J]. 福建水产, 2008(4): 60-63. LIU Y, HONGM J, YE Q T. Comparison of swing net catch before and after closed fishing seasons in Minnan sea waters[J]. Journal of Fujian Fisheries, 2008(4): 60-63. DOI:10.3969/j.issn.1006-5601.2008.04.013 |
| [7] |
张龙, 徐汉祥, 王甲刚, 等. 舟山沿岸定置张网作业休渔前后鱼类组成分析[J]. 浙江海洋学院学报(自然科学版), 2011, 30(1): 1-8. ZHANG L, XU H X, WANG J G, et al. Composition of set-net catch before and after fishing off season in Zhoushan coast areas[J]. Journal of Zhejiang Ocean University(Natural Science), 2011, 30(1): 1-8. DOI:10.3969/j.issn.1008-830X.2011.01.001 |
| [8] |
王淼, 张丹, 张玉平, 等. 杭州湾北部张网渔业资源种类组成及群落多样性初步分析[J]. 水产科技情报, 2013, 40(4): 188-192, 198. WANG M, ZHANG D, ZHANG Y P, et al. Preliminary study on Species composition and community diversity of fishery resource by set-net in north Hangzhou Bay[J]. Fisheries Science & Technology Information, 2013, 40(4): 188-192, 198. DOI:10.3969/j.issn.1001-1994.2013.04.005 |
| [9] |
宋海棠, 俞存根, 薛利建, 等. 东海经济虾蟹类[M]. 北京: 海洋出版社, 2006. SONG H T, YU C G, XUE L J, et al. Economic shrimps and crabs in East China Sea[M]. Beijing: China Ocean Press, 2006. |
| [10] |
魏崇德, 陈永寿. 浙江动物志(甲壳类)[M]. 杭州: 浙江科学技术出版社, 1991. WEI C D, CHEN Y S. Fauna in Zhejiang(Crustacean)[M]. Hangzhou: Zhejiang Science and Technology Press, 1991. |
| [11] |
郑元甲, 陈雪忠, 程家骅, 等. 东海大陆架生物资源与环境[M]. 上海: 上海科学技术出版社, 2003. ZHENG Y J, CHEN X Z, CHENG J Y, et al. Biological resources and environment of continental shelf in East China Sea[M]. Shanghai: Shanghai Science and Technology Press, 2003. |
| [12] |
原居林, 练青平, 王凯伟, 等. 瓯江干流丽水段渔业资源群落结构的季节变化[J]. 生态学杂志, 2010, 29(8): 1585-1590. YUAN J L, LIAN Q P, WANG K W, et al. Seasonal variations of fish community structure in Lishui section of Oujiang River main stream[J]. Chinese Journal of Ecology, 2010, 29(8): 1585-1590. |
| [13] |
袁梦, 陈作志, 张俊, 等. 南海北部陆坡海域中层渔业生物群落结构特征[J]. 南方水产科学, 2018, 14(1): 85-91. YUAN M, CHEN Z Z, ZHANG J, et al. Community structure of mesopelagic fish species in northern slope of South China Sea[J]. South China Fisheries Science, 2018, 14(1): 85-91. |
| [14] |
刘毅, 林小涛, 孙军, 等. 东江下游惠州河段鱼类群落组成变化特征[J]. 动物学杂志, 2011, 46(2): 1-11. LIU Y, LIN X T, SUN J, et al. Fish community changes in Huizhou segment of Dongjiang River[J]. Chinese Journal of Zoology, 2011, 46(2): 1-11. DOI:10.3969/j.issn.1671.7856.2011.02.01 |
| [15] |
汪雷, 徐东坡. 长江下游狼山沙和新开沙水域渔业资源群落结构特征分析[J]. 大连海洋大学学报, 2018, 33(1): 86-94. WANG L, XU D P. Characteristics of community structure of fishery resources in waters of Langshansha and Xinkaisha in lower reaches of the Yangtze River[J]. Journal of Dalian Ocean University, 2018, 33(1): 86-94. |
| [16] |
徐勇, 线薇微, 李文龙. 长江口及其邻近海域春季无脊椎动物群落时空变化[J]. 生物多样性, 2014, 22(3): 311-319. XU Y, XIAN W W, LI W L. Spatial and temporal variations of invertebrate community in the Yangtze River Estuary and its adjacent waters[J]. Biodiversity Science, 2014, 22(3): 311-319. |
| [17] |
马克平, 刘灿然, 刘玉明. 生物群落多样性的测度方法Ⅱβ多样性的测度方法[J]. 生物多样性, 1995, 3(1): 38-43. MA K P, LIU C R, LIU Y M. Measurement of biotic community diversity:Ⅱ β diversity[J]. Biodiversity Science, 1995, 3(1): 38-43. DOI:10.3321/j.issn:1005-0094.1995.01.007 |
| [18] |
杜飞雁, 王雪辉, 贾晓平, 等. 大亚湾海域大型底栖生物种类组成及特征种[J]. 中国水产科学, 2011, 18(4): 877-892. DU F Y, WANG X H, JIA X P, et al. Species composition and characteristics of macrobenthic fauna in Daya Bay, South China Sea[J]. Journal of Fishery Sciences of China, 2011, 18(4): 877-892. |
| [19] |
CLARKEKR, AINSWORTHM. A method of linking multivariate community structure to environmental variables[J]. Marine Ecology Progress Series, 1993, 92(3): 205-219. |
| [20] |
CLARKEK R, WARWICKR M. Change in marine communities:an approach to statistical analysis and interpretation[M]. 2nd ed. PRIMER-E: Plymout, 2001.
|
| [21] |
晏磊, 谭永光, 杨吝, 等. 南海珠江口沿岸张网渔业资源群落结构分析[J]. 生物学杂志, 2015, 32(5): 52-57. YAN L, TAN Y G, YANG L, et al. The resources community structure of stow-net fishery in the Pearl River Estuary coastal waters of the South China Sea[J]. Journal of Biology, 2015, 32(5): 52-57. DOI:10.3969/j.issn.2095-1736.2015.05.052 |
| [22] |
庞敏, 周青松, 俞存根, 等. 杭州湾海域春秋季虾蟹类群落结构及生物多样性分析[J]. 浙江海洋学院学报(自然科学版), 2015, 34(1): 1-8. PANG M, ZHOU Q S, YU C G, et al. Community structure and diversity of crustacea in Hangzhou Bay during spring and autumn[J]. Journal of Zhejiang Ocean University(Natural Science), 2015, 34(1): 1-8. DOI:10.3969/j.issn.1008-830X.2015.01.001 |
| [23] |
谢旭, 俞存根, 周青松, 等. 杭州湾海域春秋季鱼类种类组成和数量分布[J]. 海洋与湖沼, 2013, 44(3): 656-663. XIE X, YU C G, ZHOU Q S, et al. Species composition and quantitative distribution of fishes in the Hangzhou Bay during Spring and Autumn[J]. Oceanologiaet Limnologia Sinica, 2013, 44(3): 656-663. |
| [24] |
王淼, 洪波, 安传光, 等. 丰、枯水期杭州湾东北部渔业资源多样性及数量分布[J]. 上海海洋大学学报, 2016, 25(2): 246-254. WANG M, HONG B, AN C G, et al. Diversity and density distribution of fishery resources in Northeast of Hangzhou Bay during flood and drought seasons[J]. Journal of Shanghai Ocean University, 2016, 25(2): 246-254. |
| [25] |
倪勇. 长江口区凤鲚的渔业及其资源保护[J]. 中国水产科学, 1999, 6(5): 75-77. NI Y. Fishery resources conservation for Coilia mystus in the Changjiang Estuary[J]. Journal of Fishery Sciences of China, 1999, 6(5): 75-77. |
| [26] |
周永东, 薛利建, 徐开达. 舟山近海凤鲚Coilia mystus(Linnaeus)的生物学特性研究[J]. 现代渔业信息, 2004, 19(8): 19-21. ZHOU Y D, XUE L J, XU K D. A study on biological characteristics of Coilia mystus(Linnaeus) offshore of Zhoushan[J]. Modern Fisheries Information, 2004, 19(8): 19-21. DOI:10.3969/j.issn.1004-8340.2004.08.007 |
| [27] |
吴常文, 王伟洪. 杭州湾海域安氏白虾生物学与生态分布的研究[J]. 浙江水产学院学报, 1993, 12(1): 21-31. WU C W, WANG W H. On the biology and ecological distribution of shrimp, Palaemon(exopalaemon) annandalei, at Hangzhou Bay[J]. Journal of Zhejiang College of Fisheries, 1993, 12(1): 21-31. |
| [28] |
王淼, 周轩, 洪波. 杭州湾北部大型无脊椎动物群落结构[J]. 中国水产科学, 2018, 25(3): 663-673. WANG M, ZHOU X, HONG B. Community structure of macroinvertebrates in northern Hangzhou Bay[J]. Journal of Fishery Sciences of China, 2018, 25(3): 663-673. |
| [29] |
罗民波, 沈新强, 王云龙, 等. 运用ABC曲线评价工程条件下杭州湾周围海域环境变迁[J]. 农业环境科学学报, 2007, 26(s): 710-714. LUO M B, SHEN X Q, WANG Y L, et al. Environment condition evaluation of the waters around Yangshan Island in Hangzhou Bay under the huge ocean engineering by the ABC curve method[J]. Journal of Agro-Environment Science, 2007, 26(s): 710-714. |
| [30] |
唐峰华, 沈新强, 史赟荣, 等. 洋山工程海域虾蟹类资源的初步分析[J]. 上海海洋大学学报, 2010, 19(3): 372-377. TANG F H, SHEN X Q, SHI Y R, et al. Preliminary study on crab and shrimp stock biomass in waters around Yangshan Deep water Port[J]. Journal of Shanghai Ocean University, 2010, 19(3): 372-377. |