 2019,
Vol. 28
2019,
Vol. 28

2. 杭州希澳环境科技有限公司, 浙江 杭州 310012
我国是世界上的捕捞大国。近年来,大规模的过度捕捞,导致我国近海大陆架的鱼类资源衰退,而虾蟹类作为海洋中营养级较低的物种,由于缺少天敌,生存环境适宜,因此获得了较好的生存空间,种群大量繁殖。虾蟹类的营养价值和鱼类相当,在东海近岸鱼类资源遭到严重破坏,鱼类资源越来越匮乏的情况下,虾蟹类群落中一些个体大、数量多、鱼汛期规律、经济价值高的种类成为了沿海渔民的捕捞对象,目前,虾蟹类资源在海洋捕捞中占有越来越重要的地位。早期对东海虾蟹类的研究主要包含在区系[1-3]、分类学[4-6]、渔业资源评估[7]和生物学特征[8-9]研究中,有关西北太平洋蟹类群落结构方面的研究国外只在一些报道[10-11]中提到,国内首篇东海蟹类群落结构文章见于俞存根等[12]的研究,之后渔业资源的群落结构研究的文章广泛出现。近十年有关东海虾蟹类的研究主要集中在种类组成和时空分布[13]和东海近岸分布渔场的虾蟹类群落结构[14-16]等方面,但是有关东海河口区及潮下带虾蟹类群落结构的研究相对较少。渔业资源的群落结构特征与调查海域的水文、气候条件关系密切,而瓯江口海域在气候上属于暖温带向亚热带气候的过渡区域,研究瓯江口海域虾蟹类群落结构特征和空间分布,对了解河口区及气候过度区域的虾蟹类分布特征和生态类群,具有一定的应用价值。因此,本研究基于2013年5月和2013年10月在瓯江口海域渔业资源的实地调查数据,对瓯江口海域不同季节的虾蟹类群落结构,进行群落结构和空间分布的研究,旨在帮助研究者更好地了解河口区海域虾蟹类的群落分布特征及生态特征,为其他河口区海域的虾蟹类研究提供参考依据。
1 材料与方法 1.1 数据采集所用数据来源于2013年5月(春)、10月(秋)在瓯江口海域开展渔业资源的定点拖网调查数据,调查船租用群众单拖网渔船,船名为浙乐休渔122号,渔船主机功率为184 kw。调查站位设计采用系统调查方法,共设置12个站位(图 1),其中春季调查由于5号站位网具破损未捕到生物。渔业资源调查按照《海洋渔业资源调查规范》(SC/T9403—2012)进行,调查网具为底拖网,网具类型为单拖,规格为800 mm×72 mm,网囊网目尺寸20 mm,拖速约为3.5 kn。每站渔获场均装入样品袋,编号记录,低温冰鲜保存,在实验室内进行样品分类鉴定,并对虾蟹类进行称重和计数,称重使用电子天平,精确度为0.01 g。

|
图 1 瓯江口海域调查站位分布示意图 Fig. 1 Sample station in the Oujiang River estuary survey area |
由于春秋季各站位实际调查过程中拖网时间不同,所以本文以1小时为标准对调查数据进行标准化处理。针对相同的捕获方式,不同的生物其运动能力不同,所以逃逸能力也就不同,将虾类的逃逸系数定为0.3,蟹的逃逸系数定为0.2,以对标准化数据进行修正[17]。
1.3 分析方法 1.3.1 优势种优势种的判断依据为相对重要性指数(IRI),将100≤IIR<1 000的种类定义为重要种,IIR≥1 000的种类定义为优势种[13]。计算公式为
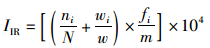 (1)
(1)
式中:ni、wi分别为第i种物种的个体数,个/km2和生物量,g/km2;N、W分别为物种的总个体数和总生物量;fi为第i种生物在m次取样中出现的次数;m为总取样次数。
1.3.2 群落结构相似性本研究采用等级聚类(非加权组的平均,即UPGAMA)的聚类分析(CLUSTER)和非度量多维标度排序(non-metric multidimensional scaling,NMDS)方法,分析瓯江口虾蟹类的群落结构特征[18]。创建“种类×站点”生物量矩阵,为了降低优势种的影响,所有数据均通过四次方根转换,然后得到Bray-Curtis相似性矩阵,在此基础上作聚类分析和NMDS二维图[19],并用胁迫系数(Stress)衡量NMDS二维点阵分布图效果,通常认为:Stress<0.05时,具有很好的代表性;0.05≤Stress<0.1时,结果基本可信;0.1≤Stress<0.2时,具有一定解释意义[20]。再通过ANOSIM显著性检验得出Global R,R=0,表示完全相同;0<R<0.5,表示差异不显著;0.5<R<0.75,表示虽有重叠,但可以分开;0.75<R<1,表示差异显著;R=1,表示完全不同;并且以上结果的显著性水平(significance)P<0.05才能接受[21]。应用SEPER(similarity percentages)方法分析各物种对样本组内相似性和组间相异性的平均贡献率,将贡献率≥4%的种类定义为主要特征种(此方法在一个群落内至少有两个站位时才能得出结果)。Bray-Curtis相似性指数计算公式[22]为
 (2)
(2)
式中:xij、xim分别为第i个种类在j站位和m站位单位时间的渔获量(经4次方根转换)。
1.3.3 ABC曲线数量生物量比较曲线(abundance biomass comparison curve,简称ABC曲线)方法是在同一坐标系中比较生物量优势度曲线和丰度优势度曲线,通过两条曲线的分布情况来分析群落不同干扰状况下的特征。若丰度优势度曲线高于生物量曲线,则群落中小个体占优势或群落处于严重干扰状态;若两曲线为相交的情况,则群落处在中等干扰状态;若生物量优势度曲线高于丰度优势度曲线,则群落中大个体占优势或群落处于未受干扰状态。在ABC曲线法中,通过W统计量展开计算,公式为
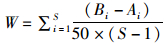 (3)
(3)
式中:Ai和Bi表示种类序号所对应的丰度和生物量的累积百分比,S为生物总种数。若生物量优势度曲线高于丰度优势度曲线,则W大于0,反之则小于0。
1.4 图表制作本研究所得的调查站位分布图、虾蟹类数量分布图均通过Sufer11软件绘制,聚类分析、NMDS、群落结构相似性贡献率计算和ABC曲线均通过软件PRIMER 5.0实现,相对重要性指数(IRI)则通过Excel表格加载宏完成。文中出现生物的拉丁名主要参照《中国海洋生物名录》[23]。
2 结果与分析 2.1 种类组成及物种数平面分布本次调查江口北部设1、2号站位,其余站位均分布在江口南部。采集的样品共鉴定出:虾蟹类25种,隶属于11科6属;秋季23种;春季18种。在科级(Family)分类单元上,春季和秋季均以长臂虾科和梭子蟹科占优势,两季中两科所占比例之和均超过40%(表 1)。春季瓯江口沿岸海域各站位种类组成中以幼体为主,秋季以成体为主。由图 2可知:春季各站位物种数在1~12种之间,平均每站约5.2种,江口北部海域站位物种数较少,平均每站2种,南部站位物种数较多,平均每站6种;秋季各站物种数在4~11种之间,平均每站8种,江口北部站位平均每站6.5种,少于平均值,南部平均每站物种数为8.8种,多于平均值。

|
表 1 瓯江口调查海域虾蟹类科、属、种的组成 Tab.1 Compositions of crab and shrimp at species, genus and family levels in Oujiang estuary survey area |

|
图 2 瓯江口调查海域春、秋季虾蟹类种类数分布 Fig. 2 Distribution of species of shrimps and crabs in spring and autumn in Oujiang estuary survey area |
由表 2可知:春季的优势种为哈氏仿对虾Parapenaeopsis hardwickii、中国毛虾Acetes chinensis、三疣梭子蟹Portunus trituberculatus;秋季的优势种为脊尾白虾Exopalaemon carinicauda、中华管鞭虾Solenocera crassicornis、三疣梭子蟹、日本蟳Charybdis japonica、锯缘青蟹Scylla serrata;春季优势种和重要种均属于广温广盐生态类群;秋季除脊尾白虾、巨指长臂虾(Palaemon macrodactylus)属于广温低盐生态类群和锈斑蟳(Charbdis feriatus)属于高温广盐生态类群外,其余种类均属于广温广盐生态类群。

|
表 2 调查海域春、秋季优势种和重要种组成特征 Tab.2 Component characteristics of dominant species and important species in spring and autumn in Oujiang estuary survey area |
对瓯江口海域春、秋季的生物量进行聚类分析和MDS排序,结果如图 3所示。综合分析,约在50%的相似性水平上,将春、秋季调查站位分为江口区、近岸区和岛礁区3个群落。春季MDS分析结果的胁迫系数Stress=0.05,秋季MDS分析结果的胁迫系数Stress=0.07,春、秋季的胁迫系数均在0.05~0.10,表明聚类结果较好。分别对春、秋季所划分的3个群落所包含的站位点进行ANOSIM分析,结果为R春季=0.769,P=0.004,R秋季=0.934,P=0.001,表明春秋季3个组群之间差异极显著,分组结果可信。春、秋季群落的站位空间分布图如图 4所示。

|
图 3 不同季节虾蟹类群落的Bray-Curtis聚类图和NMDS分析 Fig. 3 Bray-Curtis clustering and NMDS analysis of shrimp and crab communities in different seasons |

|
图 4 瓯江口调查海域春、秋季群落的空间分布 Fig. 4 Spatial distribution of community in spring and autumn in Oujiang estuary survey area |
春季:群落Ⅰ只有1个站位,所以未得到内部相似性贡献率≥4%的主要特征种;群落Ⅱ组内特征种有3种,平均相似性贡献率最高的种类是哈氏仿对虾,为76.32%,组内各站位间平均相似性为27.41%;群落Ⅲ组内特征种有2种,平均相似性贡献率最高的种类是中国毛虾,为88.59%,组内各站位间平均相似性为41.31%。春季群落间平均相异性均在75%以上,对组间相异性贡献较高的种类为哈氏仿对虾和中国毛虾。秋季, 各群落内部平均相似性在40%左右,各群落内部平均相似性贡献率≥4%的主要特征种均多于3种:群落Ⅰ和群落Ⅲ中最高的是脊尾白虾,分别为63.28%和33.09%;群落Ⅱ中最高的是日本蟳,为36.32%。秋季群落间平均相异性均在70%以上,对组间相异性贡献较高的种类为锯缘青蟹和三疣梭子蟹。春秋季各群落内主要特征种的相似性或相异性贡献率见表 3、表 4。

|
表 3 不同季节虾蟹类群落内部平均相似性贡献率≥4%的主要特征种 Tab.3 Main characteristic species with average contribution rate≥ 4%within the shrimp and crab community in different seasons |

|
表 4 不同季节虾蟹类群落间平均相异性贡献率≥4%的主要特征种 Tab.4 Main characteristic species with average contribution rate ≥ 4% within shrimp and crab communities in different seasons |
春、秋季各群落的ABC曲线趋势如图 5所示,春季瓯江口海域3个群落的丰度优势度曲线均位于生物量优势度曲线上方;秋季瓯江口海域3个群落中,除了群落Ⅰ的丰度优势度曲线位于生物量优势度曲线上方外,其余两个群落的丰度优势度曲线均位于生物量优势度曲线下方。从W统计量来看,春季群落状态受外界扰动较大,秋季群落状态除群落Ⅰ受外界扰动较大外,其余群落相对较稳定。

|
图 5 瓯江口调查海域春、秋季群落的ABC曲线及W统计值 Fig. 5 ABC Curves and W statistical values of spring and autumn communities in Oujiang estuary survey arer |
海区的生物区系特征是在海区内不同种群的相互联系和环境的共同作用下形成的生物适应环境的的具体表现[24]。因此,一个海区的种类组成情况和物种数分布情况,一方面与物种的生态习性有关,另一方面则是环境因子对生物的影响和制约。瓯江口海域东北方向有江浙沿岸流,东南方向有来自台湾暖流的高温高盐水,西侧有瓯江的冲淡水[25]。因此,瓯江口海域的种类组成和分布具有特有的特点。从虾蟹类的种类组成来看,瓯江口调查海域春季有18种,秋季有23种,总计25种,其中共有种为15种,春秋季种类以广温广盐种类和广温低盐种类为主,未见冷水种,这与徐兆礼[26]对该海域的研究结果一致。本研究发现:春季瓯江口沿岸海域各站位分布大量优势种类的幼体,如哈氏仿对虾(0.68 g/尾)、葛氏长臂虾(Palaemon gravieri)(0.45 g/尾)、三疣梭子蟹(7.23 g/尾)的幼体;秋季瓯江口海域分布较多的脊尾白虾(2.26 g/尾)、周氏新对虾(5.92 g/尾)、三疣梭子蟹(50.50 g/尾)的成体。有研究[27-28]表明河口和浅湾地区一般为海洋生物的产卵场和索饵场。由此可以推测,瓯江口海域春季种类组成以幼体或小型个体为主,秋季主要以成体为主,同时推测瓯江口春季海域可能存在哈氏仿对虾、三疣梭子蟹等具有洄游习性生物的产卵场和索饵场。
3.2 种类数分布及优势种生态类型分析由调查海域虾蟹类丰富度分布图(图 2)可以看出,本研究春、秋季各区域捕获的渔获物的种类数相差较大,春季物种丰富度较高的区域分布在近岸海域,较低的站位分布在江口堤坝北区和岛礁区,秋季江口堤坝南部物种数分布趋势与春季相反,越远离近岸丰富度越高。同时研究区域内各站位的基础数据发现,每个站位除了优势种的尾数较多外其他种类的尾数相对较少。造成这两种情况有以下3个原因:(1)可能是受调查时间的限制,本研究两次出海采样均在白天进行,而虾蟹类大都有昼伏夜出的生活习性[29];(2)可能是调查站位底质的差异,底栖生物对生活环境的底质具有选着性,例如蟹类喜欢沙质底及礁石较多的区域;(3)也可能是由于过度捕捞和环境退化造成资源衰退,各物种的种群变小,降低了被捕获的概率[30]。
虾蟹类的生态类群与海洋环境的关系十分密切,根据虾蟹类的分布水深,分布海域的水温、盐度性质,将东海海区大陆架的虾蟹类划分为广温低盐生态类群、广温广盐生态类群和高温广盐生态类群和高温高盐生态类群等四大类[31]。金显仕等[32]研究长江口渔业生物的群落特征结果显示,长江口海域生物以暖温性和暖水性种类为主,罕见冷水种,且大多数暖温性种类都具有洄游习性。瓯江口海域与长江口海域在地理位置上具有相同的经度,但纬度更低,结合长江口的研究成果,从表 2中春秋季优势种和重要种的组成可以看出,多数种类属于广温广盐生态类群,如哈氏仿对虾、三疣梭子蟹、日本
春、秋季虾蟹类的调查站位可分为3个群落组(见图 3,图 4):群落Ⅰ江口堤坝北部,主要典型种为安氏白虾(Exopalaemon annandalei)、脊尾白虾等;群落Ⅱ堤坝南部近岸区,主要典型种为哈氏仿对虾、中国毛虾、三疣梭子蟹等;群落Ⅲ岛礁区,主要典型种为哈氏仿对虾、中华管鞭虾、三疣梭子蟹等。上述典型种中,除了安氏白虾、脊尾白虾为河口、浅海区种类外,其余以海洋性种类为主,在春、夏季集中在沿岸近海索饵、产卵,秋、冬季随水温降低逐渐洄游到深海进行越冬。研究各群落内的生物组成发现,群落Ⅰ内物种的丰富度较低,且与其他群落物种组成差异较大,主要种类为安氏白虾和脊尾白虾,出现这种情况的原因是瓯江口昆灵岛以南建有浅堤坝,泥沙堆积导致瓯江径流的主要下泄途径为昆灵岛北部河道[33],大量的淡水团径流使得河口区盐度较低,适合广温低盐种类生活,从而导致群落Ⅰ的虾蟹类丰富度较低。
除了盐度对群落结构的影响外,水深也是影响群落结构的重要因素。瓯江口海域地势西北高东南低,从北到南水深逐步增大[26]。此次瓯江口海域群落聚类分析得出的春、秋季群落组的分布结果与水深有明显的关联性,群落分布明显呈西北向东南分布(图 3,图 4),这与瓯江口海域的地形走势相一致。同时分析各群落组内的种类组成发现由河口区到深海区的物种组成与水深环境也有一定适应性,例如:群落Ⅰ位于河口海域,水深较浅导致海水与河水的混合程度较高,其组内典型种为安氏白虾等广温低盐种;而群落Ⅲ位于中心海域,水深较深,温盐条件相对稳定,其组内典型种为三疣梭子蟹等。相关研究也得出,对海洋生物群落结构具有最大影响的因素即为水深,水深不断变化时水温和盐度会发生相应的变化[34]。
群落聚类结果结合虾蟹类群落内平均相似性贡献率和群落间平均相异性贡献率≥4%的主要特征种(表 3、表 4)分析发现,各群落组内的典型种在一定程度上又是群落组间的分歧种,如春季的哈氏仿对虾、中国毛虾等,秋季的中华管鞭虾、三疣梭子蟹等。进一步分析发现各群落组内的特征种均为群落的优势种或重要种,且多为小个体或幼体。群落中的优势种和重要种及群落组中的特征种主要以较低营养级生物为主,而较高营养级的生物在群落中不占优势或未出现,此结果与金显仕等[32]研究的长江口渔业生物群落特征中种类组成及优势种的结果一致。由于群落中较高营养级较少,无法通过下行控制机制对低营养级种类的数量进行有效控制[35],如中国毛虾。低营养级的种类具有种群内部波动机制,在适宜条件下种群增长迅速,反之衰退迅速。低营养级种类的种群内部的波动,也将成为影响群落结构稳定性的重要因素。
BLANCHARD等[36]认为鱼类群落中ABC曲线特征反映了群落中大型种类和小型种类相对数量的变化以及个体大小组成的变化,群落中优势种类的大小决定了生物量优势度曲线和数量优势度曲线的位置[37]。本研究中春季瓯江口海域3个群落的丰度优势度曲线均高于生物量优势度曲线,说明春季小型种类居多,这与春季各群落内虾蟹类的种类组成的优势种主要为小型种类或大型种类的幼体的结果相符,如哈氏仿对虾(平均质量为0.68 g)、中国毛虾(平均质量为0.31 g)、三疣梭子蟹(平均质量为7.23 g)均为幼体。秋季瓯江口海域3个群落中,除了群落Ⅰ的丰度优势度曲线高于生物量优势度曲线外,其余两个群落的生物量优势度曲线均高于丰度优势度曲线,说明秋季大型种类或成体居多,此结果同样与秋季虾蟹类群落内优势种的平均重量的出的结果相符,如脊尾白虾(平均质量为2.26 g)、三疣梭子蟹(平均质量为50.50 g)、锯缘青蟹(平均质量为80.90 g)均为成体。因此,ABC曲线的结果可以很好地反映本研究瓯江口调查海域春秋季虾蟹类的群落结构。但是,春秋季群落Ⅰ的W值均小于零,这可能是由于群落Ⅰ位于瓯江口北部河口区的主航道上,人类活动较为频繁,干扰较多。
| [1] |
刘瑞玉. 黄海及东海经济虾类区系的特点[J]. 海洋与湖沼, 1959, 2(1): 35-42. LIU R Y. Notes of the economic macrurous crustacean fauna of the Yellow Sea and the East China Sea[J]. Oceanologia et Limnologia Sinica, 1959, 2(1): 35-42. |
| [2] |
沈嘉瑞, 刘瑞玉. 中国海蟹类区系特点的初步研究[J]. 海洋与湖沼, 1963, 5(2): 139-153. SHEN J R, LIU R Y. Preliminary studies on the characteristics of the crab fauna of China Seas[J]. Oceanologia et Limnologia Sinica, 1963, 5(2): 139-153. |
| [3] |
刘瑞玉, 徐鳳山. 黃、东海底栖动物区系的特点[J]. 海洋与湖沼, 1963, 5(4): 206-321. LIU R Y, XU F S. Preliminary studies on the benthic fauna of the Yellow Sea and East China Sea[J]. Oceanologia et Limnologia Sinica, 1963, 5(4): 206-321. |
| [4] |
戴爱云, 杨思琼, 宋玉枝, 等. 中国海洋蟹类[M]. 北京: 海洋出版社, 1986: 1-641. DAI A Y, YANG S Q, SONG Y Z, et al. Chinese marine crab[M]. Beijing: Maritime Press, 1986: 1-641. |
| [5] |
魏崇德. 浙江动物志-甲壳类[M]. 杭州: 浙江科学技术出版社, 1991: 1-481. WEI C D. Animal zoology of Zhejiang province-crustacean[M]. Hangzhou: Zhejiang Science and Technology Press, 1991: 1-481. |
| [6] |
刘文亮, 何文珊. 长江河口大型底栖无脊椎动物[M]. 上海: 上海科学技术出版社, 2007: 1-203. LIU W L, HE W S. Benthic macro-invertebrates in the Yangtze estuary[M]. Shanghai: Shanghai Science and Technology Press, 2007: 1-203. |
| [7] |
俞存根, 宋海棠, 姚光展. 东海大陆架海域蟹类资源量的评估[J]. 水产学报, 2004, 28(1): 41-46. YU C G, SONG H T, YAO G Z. Assessment of the crab stock biomass in the continental shelf waters of the East China Sea[J]. Journal of Fisheries of China, 2004, 28(1): 41-46. |
| [8] |
朱成德. 虾蟹类的幼体发育[J]. 水产养殖, 1989(3): 26-27. ZHU C D. Larval development of shrimps and crabs[J]. Journal of Aquaculture, 1989(3): 26-27. |
| [9] |
俞存根, 宋海棠, 姚光展. 东海细点圆趾蟹的生物学特性[J]. 水产学报, 2004, 28(6): 657-662. YU C G, SONG H T, YAO G Z. Biological characteristics of Ovalipes punctatus in the East China Sea[J]. Journal of Fisheries of China, 2004, 28(6): 657-662. |
| [10] |
MUROGA K, SSZUIK K, ISHIMARU K, et al. Vibriosis of swimming crab Portunus trituberculatus in larviculture[J]. Journal of the World Aquaculture Society, 1994, 25(1): 50-54. DOI:10.1111/jwas.1994.25.issue-1 |
| [11] |
YANAGI T, TSUKAMOTO H, IGAWA S, et al. Recruitment strategy of swimming crab, Portunus trituberculatus, in Hiuchi-Nada, Japan[J]. Fisheries Oceanography, 1995, 4(3): 217-229. DOI:10.1111/fog.1995.4.issue-3 |
| [12] |
俞存根, 宋海棠, 姚光展. 东海蟹类群落结构特征的研究[J]. 海洋与湖沼, 2005, 36(3): 213-220. YU C G, SONG H T, YAO G Z. Crab community structure in the East China Sea[J]. Oceanologia et Limnologia Sinica, 2005, 36(3): 213-220. DOI:10.3321/j.issn:0029-814X.2005.03.004 |
| [13] |
郑献之, 俞存根, 陈小庆, 等. 舟山渔场及邻近海域蟹类种类组成和时空分布[J]. 海洋与湖沼, 2012, 43(1): 147-153. ZHENG X Z, YU C G, CHEN X Q, et al. Species composition and spatio-temporal distribution of crabs in Zhoushan fishing ground and its adjacent areas[J]. Oceanologia et Limnologia Sinica, 2012, 43(1): 147-153. |
| [14] |
陈小庆.东海中南部外海虾蟹类群落结构特征的研究[D].舟山: 浙江海洋学院, 2010. CHEN X Q. Community characteristics of shrimps and crabs in the offshore waters of the mid-southern East China Sea[D]. Zhoushan: Zhejiang Ocean University, 2010. http://cdmd.cnki.com.cn/article/cdmd-10340-2010107862.htm |
| [15] |
徐开达, 卢占晖, 薛利建, 等. 中街山列岛保护区蟹类群落结构特征的研究[J]. 上海海洋大学学报, 2012, 21(6): 1032-1037. XU K D, LU Z H, XUE L J, et al. Crab community structure in the protected Zhongjieshan Islands area[J]. Journal of Shanghai Ocean University, 2012, 21(6): 1032-1037. |
| [16] |
夏陆军, 俞存根, 蔡厚才, 等. 南麂列岛海洋自然保护区虾类群落结构及其多样性[J]. 海洋学报, 2016, 38(2): 73-83. XIA L J, YU C G, CAI H C, et al. Community structure and diversity of shrimp in Nanji Islands marine conservation area[J]. Journal of Oceanography, 2016, 38(2): 73-83. DOI:10.3969/j.issn.0253-4193.2016.02.007 |
| [17] |
吕振波. 山东近海经济生物资源调查与评价[M]. 北京: 海洋出版社, 2010. LV Z B. Investigation and evaluation of economic resources in Shandong offshore[M]. Beijing: China Ocean Press, 2010. |
| [18] |
周红, 张志南. 大型多元统计软件PRIMER的方法原理及其在底栖群落生态学中的应用[J]. 青岛海洋大学学报, 2003, 33(1): 58-64. ZHOU H, ZHANG Z N. Rationale of the multivariate statistical software PRIMER and its application in benthic community ecology[J]. Journal of Ocean University of Qingdao, 2003, 33(1): 58-64. DOI:10.3969/j.issn.1672-5174.2003.01.023 |
| [19] |
BRAZNER J C, BEALS E W. Patterns in fish assemblages from coastal wetland and beach habitats in Green Bay, Lake Michigan:a multivariate analysis of abiotic and biotic forcing factors[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1997, 54(8): 1743-1761. DOI:10.1139/f97-079 |
| [20] |
CLARKE K R. Non-parametric multivariate analyses of changes in community structure[J]. Austral Ecology, 1993, 18(1): 117-143. |
| [21] |
FIELD J G, CLARKE K R, WARWICK R M. A practical strategy for analyzing multispecies distribution patterns[J]. Marine Ecology-Progress Series, 1982, 8: 37-52. DOI:10.3354/meps008037 |
| [22] |
张焕君, 李凡, 丛日翔, 等. 黄河口海域无脊椎动物群落结构及其变化[J]. 中国水产科学, 2014, 21(4): 800-809. ZHANG H J, LI F, CONG R X, et al. Community structure of invertebrate and its change in Huanghe (Yellow River) Estuary[J]. Journal of Fishery Sciences of China, 2014, 21(4): 800-809. |
| [23] |
刘瑞玉. 中国海洋生物名录[M]. 北京: 科学出版社, 2008: 1-1267. LIU R Y. Checklist of marine biota of China seas[M]. Beijing: Science Press, 2008: 1-1267. |
| [24] |
胡成业, 徐衡, 水柏年, 等. 温州瓯飞滩邻近海域春季游泳动物群落结构及多样性[J]. 南方水产科学, 2015, 11(3): 7-15. HU C Y, XU H, SHUI B N, et al. Community structure and biodiversity of nekton near Oufei Beach of Wenzhou[J]. South China Fisheries Science, 2015, 11(3): 7-15. DOI:10.3969/j.issn.2095-0780.2015.03.002 |
| [25] |
姚炎明, 陈吉余, 陈永平, 等. 温州湾水流及其与地形的关系[J]. 华东师范大学学报(自然科学版), 1998(2): 61-68. YAO Y M, CHEN J Y, CHEN Y P, et al. The relationship between current and topography in Wenzhou Bay[J]. Journal of East China Normal University (Natural Science), 1998(2): 61-68. |
| [26] |
徐兆礼. 瓯江口海域夏秋季蟹类数量分布与环境的关系[J]. 水产学报, 2009, 33(2): 237-244. XU Z L. Relationship of crab density distribution with environment in the Oujiang Estuary during Summer and Autumn[J]. Journal of Fisheries of China, 2009, 33(2): 237-244. |
| [27] |
MCBRIDE R S, CONOVER D O. Recruitment of young-of-the-year bluefish Pomatomus saltatrix to the New York Bight:variation in abundance and growth of spring-and summer-spawned cohorts[J]. Marine Ecology Progress Series, 1991, 78: 205-216. DOI:10.3354/meps078205 |
| [28] |
BRADSTOCK M, GORDON D P. Coral-like bryozoan growths in Tasman Bay, and their protection to conserve commercial fish stocks[J]. New Zealand Journal of Marine and Freshwater Research, 1983, 17(2): 159-163. DOI:10.1080/00288330.1983.9515993 |
| [29] |
戴爱云, 冯钟琪, 宋玉枝, 等. 三疣梭子蟹渔业生物学的初步调查[J]. 动物学杂志, 1977, 12(2): 30-33. DAI A Y, FENG Z Q, SONG Y Z, et al. Preliminary survey of fishery biology in Portunus trituberculatus[J]. Chinese Journal of Zoology, 1977, 12(2): 30-33. |
| [30] |
吕振波, 李凡, 王波, 等. 黄海山东海域春、秋季鱼类群落结构[J]. 水产学报, 2011, 35(5): 692-699. LÜ Z B, LI F, WANG B, et al. Community structure of fish resources in spring and autumn in the Yellow Sea off Shandong[J]. Journal of Fisheries of China, 2011, 35(5): 692-699. |
| [31] |
宋海棠, 俞存根, 薛利建, 等. 东海经济虾蟹类[M]. 北京: 海洋出版社, 2006: 5-14. SONG H T, YU C G, XUE L J, et al. Donghai economic shrimp and crab[M]. Beijing: China Ocean Press, 2006: 5-14. |
| [32] |
金显仕, 单秀娟, 郭学武, 等. 长江口及其邻近海域渔业生物的群落结构特征[J]. 生态学报, 2009, 29(9): 4761-4772. JIN X S, SHAN X J, GUO X W, et al. Community structure of fishery biology in the Yangtze River estuary and its adjacent waters[J]. Acta Ecologica Sinica, 2009, 29(9): 4761-4772. DOI:10.3321/j.issn:1000-0933.2009.09.021 |
| [33] |
祝永康. 瓯江溺谷海湾充填成陆和河口湾的形成过程[J]. 地理学报, 1993, 48(3): 254-261. ZHU Y K. Formation and filling up of Oujiang river estuary in Zhejing Province[J]. Acta Geographica Sinica, 1993, 48(3): 254-261. DOI:10.3321/j.issn:0375-5444.1993.03.009 |
| [34] |
刘维达, 林昭进, 江艳娥, 等. 南海北部陆架区底层渔业资源的空间分布特征[J]. 热带海洋学报, 2011, 30(5): 95-103. LIU W D, LIN Z J, JIANG Y E, et al. Spatial distribution of demersal fishery resources in the continental shelf of the northern South China Sea[J]. Journal of Tropical Oceanography, 2011, 30(5): 95-103. DOI:10.3969/j.issn.1009-5470.2011.05.013 |
| [35] |
李凡, 张焕君, 吕振波, 等. 莱州湾游泳动物群落种类组成及多样性[J]. 生物多样性, 2013, 21(5): 537-546. LI F, ZHANG H J, LÜ Z B, et al. Species composition and community diversity of nekton in Laizhou Bay, China[J]. Biodiversity Science, 2013, 21(5): 537-546. |
| [36] |
BLANCHARD F, LELOC'H F, HILY C, et al. Fishing effects on diversity, size and community structure of the benthic invertebrate and fish megafauna on the Bay of Biscay coast of France[J]. Marine Ecology Progress Series, 2004, 280: 249-260. DOI:10.3354/meps280249 |
| [37] |
VERGNON R, BLANCHARD F. Evaluation of trawling dis-turbance on macrobenthic invertebrate communities in the Bay of Biscay, France:abundance biomass comparison (ABC method)[J]. Aquatic Living Resources, 2006, 19(3): 219-228. DOI:10.1051/alr:2006022 |
 ,
SONG Weihua2,
YU Cungen1
,
SONG Weihua2,
YU Cungen1 ,
ZHOU Qingsong2,
ZHENG Ji1,
LIU Hui1,
DENG Xiaoyan1,
ZHANG Ping1
,
ZHOU Qingsong2,
ZHENG Ji1,
LIU Hui1,
DENG Xiaoyan1,
ZHANG Ping1
2. Hangzhou Xiao Environmental Technology Co. LTD., Hangzhou 310012, Zhejiang, China