 2019,
Vol. 28
2019,
Vol. 28

2. 上海海洋大学 农业部鱼类营养与环境生态研究中心, 上海 201306;
3. 上海海洋大学 水产动物遗传育种中心上海市协同创新中心, 上海 201306;
4. 安徽省衡安学校, 安徽 合肥 231137;
5. 杭州千岛湖发展集团有限公司, 浙江 杭州 311700
浮游植物是湖泊生态系统的初级生产者,其种类和数量的变化直接影响到水域生态系统的结构和功能,对维护水域生态系统的健康与稳定起着重要的作用[1]。浮游植物的物种组成及演替与环境因子关系密切[2],从非生物环境来看,光照和温度是影响浮游植物分布的重要因素[3]。水体中的氮磷含量也决定了浮游植物的分布,氮磷等营养物质的过度输入促进了水体富营养化[4]。从生物环境来说,浮游动物的群落结构及滤食性鱼类的放养,也会对浮游植物群落产生影响[5-8]。浮游植物群落结构的改变可以表征水环境的变化[9-10],而对浮游植物群落结构特征的阐述仍不够详尽[11-14],因此,研究浮游植物的变化规律可以为水环境的保护提供重要的依据。
千岛湖,原名新安江水库,位于浙江省西部山区与安徽省交界处,地处亚热带中部东南沿海季风区,气候温暖,四季分明,雨量充沛,年平均气温约17 ℃[14]。水位为108 m时其水域面积达573 km2,库容量达178.4亿m3,其水资源量约占钱塘江流域水资源量的30%,是钱塘江的重要水源地。千岛湖一直达到国家Ⅰ类和Ⅱ类地表水标准,自2014年开始建设千岛湖引水工程,并作为杭州市第二饮用水源地,因此其水域生态环境的健康状况直接影响到饮用水的水质安全。
传统浮游植物鉴定是在分类水平上进行的,根据物种同源性特点将其归入不同的类群,很少体现环境生态特征。鉴于浮游植物群落高度多样化而且大多数物种的发生难以预测。依据浮游植物生理、生态、形态特点,REYNOLDS[10]提出了相对完整的浮游植物功能群理论,其主要内容就是属于同一功能群的浮游植物通常生长于相同的生境类型或相似的环境条件下[15],已被广泛应用于湖泊浮游植物的研究。本研究通过探究浮游植物群落结构及功能群的划分,为了解千岛湖水环境变动规律及水环境的保护与管理提供依据。
1 材料与方法 1.1 采样站点设置和采样频率2015年每月中旬在千岛湖进行浮游植物样品采集。根据千岛湖的水流方向和动力学特点,在千岛湖上中下游共设置5个采样点(图 1),即S1(宅上,上游河流区),S3(温馨岛,中游过渡区),S4(猴岛,中游过渡区),S8(姥山,下游湖泊区)和S9(密山,下游湖泊区)。其中S1、S4和S9 3个采样点划分为6个水层,即0.5、4、8、12、16、20 m,对每个水层采集分层水样和浮游植物样品,数据处理时,取各站点所有水层平均值进行统计分析;S3和S8采样点采集混合水样和浮游植物样品。千岛湖常年四季分明,春季4—6月为丰水期,冬季的12月、1—2月为枯水期。

|
图 1 千岛湖采样站点图 Fig. 1 Map of sampling sites in Qiandao Lake |
采用YSI-58型溶氧仪现场测定水温(WT)和溶解氧(DO),使用pH-4型便携式pH计测定水体酸碱度(pH),用塞氏盘法测定透明度(SD)。将采集的水样带回实验室置于4 ℃冰箱,并在12 h之内测定总氮(TN,GB11894—89)、总磷(TP,GB11893—89)和高锰酸盐指数(CODMn,GB11892-89)。
1.3 浮游植物样品处理与实验方法采用5 L柱状采水器对所设置的采样点采集样品,得到不同水层处水样或混合水样各1 L,样品采集完后立即加入Lugol试剂(15%)和3 mL甲醛(4 %),带回实验室置于分液漏斗中静置沉淀48 h,收集后在光学显微镜下进行种类鉴定和细胞计数,浮游植物的鉴定主要参考《中国淡水藻志》[16]以及《中国淡水藻类:系统、分类及生态》[17]。浮游植物功能群的分类参考REYNOLDS[10]及PASISÁK等[9]增补的浮游植物功能群分类方法进行分类。
1.4 数据处理和分析运用Shannon-Wiener多样性指数(H')、Pielou均匀度指数(J)、Margalef丰富度指数(D)以及优势度指数(Y)公式,来计算和分析浮游植物群落的多样性及判断优势种[18-19],计算公式如下:
 (1)
(1)
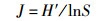 (2)
(2)
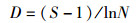 (3)
(3)
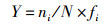 (4)
(4)
式中:ni为第i种浮游植物的个数;N为所有浮游植物总个数;S为样品中浮游植物种类数;fi为第i种浮游植物在各站点出现的频率,将Y > 0.02的浮游植物定为优势种, 采用Origin 8.5软件进行作图,使用IBM SPSS 19.0软件对各月份间浮游植物生物密度和生物量差异性分析,以P < 0.05为显著性差异,使用CANOCO for Windows 5.0软件对功能群与环境因子进行CCA典范对应分析。
2 结果与分析 2.1 千岛湖的理化因子从表 1中可以看出:透明度、温度、溶解氧和pH在各采样站点之间无显著性差异(P > 0.05)。TN、TP及高锰酸盐指数出现一定的差异性,其中S1(河流区)总氮显著高于其他站点(P < 0.05),S9(湖泊区)最低,为(1.08 ± 0.14) mg/L;河流区(S1)总磷显著高于湖泊区(S8,S9,P < 0.05),S9最低,为(0.009 ± 0.005) mg/L;S8站点的高锰酸盐指数显著低于S1站点(P < 0.05)。营养盐呈现出由河流区经过渡区至湖泊区降低的趋势。

|
表 1 千岛湖各站点主要理化因子 Tab.1 Annual average of main physicochemical factors in Qiandao Lake |
2015年共鉴定出浮游植物8门201种,隶属绿藻门(Chlorophyta)、硅藻门(Bacillariophyta)、蓝藻门(Cyanophyta)、隐藻门(Cryptophyta)、裸藻门(Euglennophyta)、黄藻门(Xanthophyta)、甲藻门(Pyrrophyta)和金藻门(Chrysophyta)。其中:蓝藻门(43种,21.39%),主要优势种为水华鱼腥藻(Anabaena aquae)、假鱼腥藻(Pseudanabaena sp.)、不定微囊藻(Microcystisincerta)、湖泊鞘丝藻(Lyngbyalimnetica)、两栖颤藻(Oscillatoriaamphibia)和小席藻(Phorimidiumtenus);绿藻门81种,占40.30%, 无优势藻种;硅藻门57种,占28.36%,优势种为小环藻(Cyclotella sp.)、克罗钝脆杆藻(F.crotomensis)和颗粒直链藻(M. granulata),这三大门类物种数最多。此外,甲藻门(8种,3.98%)、金藻门(1种,0.50%)、隐藻门(3种,1.49%)、黄藻门(3种,1.49%)以及裸藻门(5种,2.49%)物种数较少(图 2,表 2)。

|
图 2 千岛湖浮游植物各门类物种数 Fig. 2 Species of phytoplankton in Qiandao Lake |

|
表 2 千岛湖浮游植物优势种组成及其优势度 Tab.2 Dominant phytoplankton species and their dominance in Qiandao Lake |
从季节变化上看:浮游植物的种类数以秋季最多,为147种,其余季节为冬季(128种) > 春季(127种) > 夏季(110种),见图 3;绿藻门种类数全年占据优势,其次是蓝藻门和硅藻门;蓝藻门和绿藻门均以春季最少、秋季最多,硅藻门则是春季最多、秋季最少;除蓝藻门、绿藻门和硅藻门外其他门类浮游植物物种数均较少。

|
图 3 千岛湖浮游植物各季节门类组成 Fig. 3 Composition of species of phytoplankton at Qiandao Lake in each season |
浮游植物生物密度和生物量的组成不尽一致,千岛湖浮游植物生物密度全年平均值为(1 285.41±448.43)×104个/L,生物密度在各季节均以蓝藻门为绝对优势门类,秋季最高占88.35%,冬季最低,月份间生物密度差异极显著(F=45.503,P < 0.001)。而蓝藻生物量的最大值仅出现在秋季,最大值为9月份的9.44 mg/L,其余季节为硅藻门占优势,月份间生物量差异极显著(F=34.164,P < 0.001)。见图 4。

|
图 4 千岛湖浮游植物生物量与生物密度 Fig. 4 Phytoplankton biomass and biological density in Qiandao Lake |
从不同的采样点来看:2015年浮游植物Shannon多样性指数为2.055~2.371,最高值出现在1号点,最低值出现在8号点;Pielou均匀度指数为0.625~0.704,与Shannon多样性指数略有不同的是,最高值出现在1号点,最低值则出现在9号点;Margalef丰富度指数为3.918~4.644,与Shannon多样性指数相同,最大值出现在1号点,最小值出现在8号点。见表 3。

|
表 3 千岛湖各站点浮游植物多样性指数分布 Tab.3 Distribution of phytoplankton diversity at each sampling station in Qiandao Lake |
由图 5可以看出,在调查期间,各门类种类数在各采样点间存在一定的差异,但均以绿藻门物种数为多,且3、8号点的种类数较少,1、4和9号点种类数较多,1号点种类数最多,有147种。统计分析显示,1、4和9号点之间均无显著性差异(Friedman检验,X2=1.900,df=2,P=0.387),3号点和8号点之间差异也不显著(Wilcoxon符号秩检验,Z=-0.577,P=0.564)。

|
图 5 千岛湖各采样点浮游植物种数 Fig. 5 Phytoplankton species at each sampling station of Qiandao Lake |
按照水流方向来看(图 6),水库河流区浮游植物生物密度总体来说最低,多个季节为最低值。全年生物密度分布由高至低为湖泊区(15.395 0×106个/L)、过渡区(12.152 4×106个/L)和河流区(9.671 6×106个/L),方差分析显示,三者之间无显著性差异(P > 0.05)。

|
图 6 千岛湖浮游植物生物密度的空间分布 Fig. 6 Spatial distribution of phytoplankton density in Qiandao Lake |
依据REYNOLDS[10]及PASISÁK等[9]增补的浮游植物功能群分类方法对千岛湖浮游植物组成进行划分。本研究采集的浮游植物样品经镜检鉴定,可划分为24个功能类群(表 5)。

|
表 5 千岛湖主要浮游植物功能类群组成 Tab.5 Main composition of phytoplankton functional groups in Qiandao Lake |
浮游植物功能类群组成存在显著的时空差异。春季的浮游植物功能类群主要有B、S1、J、K、H1(图 7)。在S1(河流区):功能群B占绝对优势,相对丰度为37.0%;功能群S1次之,占14.7%左右;J功能群相对丰度为12.4%。S3过渡区S1功能群占绝对优势,相对丰度为19.7%;功能群B次之,占16.4%;以隐球藻为主的功能群K相对丰度占15.1%左右。S4(过渡区)的主要功能群是B,相对丰度为30.6%左右,与前几个站点不同:功能群H1在此站点相对丰度明显上升(15.5%);占比较高的是J功能群和S1功能群,分别为12.8%和10.7%。在8号点(湖泊区),相对丰度最高的是功能群X1,功能群B(12.5%)和以鱼腥藻为主的功能群H1(10.2%)也占有一定的优势。9号点(湖泊区)占比最高的是功能群H1,其次是B(16.0%)和S1(15.6%)。

|
图 7 春季千岛湖各样点浮游植物功能群组成 Fig. 7 Composition of phytoplankton functional groups in Qiandao Lake in spring |
夏季的功能类群主要有TC、J、S1、M、LO、H1和P(图 8)。S1(河流区)主要功能群有6类,相对丰度由高到低分别为功能群TC(占比为15.91%)、功能群J(14.0%)、功能群S1(13.9%)、功能群M(13.1%)、功能群LO(11.5%)和功能群H1(10.2%)。S3(过渡区)在夏季占绝对优势的是功能群P,占比高达61.2%左右,其次占优势的是功能群TC和功能群S1。S4(过渡区)占主要优势的是功能群P和功能群M,占比分别为25.6%和24.2%。其次为功能群TC和功能群S1,占比分别为14.4%和12.2%。8号点(湖泊区)占主要优势的是功能群S1,其次为功能群M、功能群H1和功能群TC。9号点(湖泊区)的绝对优势功能群是功能群M,相对丰度达52.2%,功能群P(13.5%)和功能群S1(10.5%)。

|
图 8 夏季千岛湖各样点浮游植物功能群组成 Fig. 8 Composition of phytoplankton functional groups in Qiandao Lake in summer |
秋季的功能类群主要有M、S1、TC、K、H1、J、P和LO(图 9),1号点(河流区)秋季的主要功能群有3类:主要优势功能群M,相对丰度为32.1%;其次为功能群S1,相对丰度为28.6%,功能群TC,相对丰度为15.4%。3号点(过渡区)的主要功能群与1号点存在明显的不同。其中相对丰度占比最大的功能群为TC(27.9%),其次为功能群P(19.9%)。4号点(湖泊区)的优势功能群类有5类,分别为功能群LO(以甲藻、色球藻为主)、K、M、TC和S1。8号点(湖泊区)占最大优势的是功能群S1,相对丰度为18.7%左右,功能群K、H1、J也具有一定的优势,相对丰度分别为14.7%、14.4%和14.2%。9号点(湖泊区)优势功能群较少,仅2种,分别是功能群S1(11.8%)和功能群TC(10.5%)。

|
图 9 秋季千岛湖各样点浮游植物功能群组成 Fig. 9 Composition of phytoplankton functional groups in Qiandao Lake in autumn |
冬季的功能类群主要有Y、P、B、LO、S1、H1、TC和D(图 10)。1号点(河流区)占主要优势的为功能群Y,相对丰度为27%左右;其次功能群P和B,相对丰度分别为19.4%和18.7%。3号点(过渡区):功能群P在冬季占主要优势,为32.1%;功能群TC次之,占16.1%;功能群Y和功能群D也占一定比例。4号点(过渡区):主要优势功能群为功能群S1,相对丰度占18.2%;功能群B、Y、P也占有一定优势。8号点(湖泊区):功能群Y占24.2%左右;其次为功能群B,相对丰度为19.5%左右;功能群S1也占有一定的比例。9号点(湖泊区):主要功能群是Y,相对丰度占20%左右;其次是H1,相对丰度为14.5%;功能群B和LO也占一定的比例。

|
图 10 冬季千岛湖各样点浮游植物功能群组成 Fig. 10 Composition of phytoplankton functional groups in Qiandao Lake in winter |
从以上可以看出,S1和H1浮游植物功能类群在四个季节均具有比较高的丰度比例,B浮游植物功能类群在春季和冬季具有比较高的丰度比例,J浮游植物功能类群在春季、夏季和秋季占比较高,浮游植物功能类群K在春季和秋季占比较高,TC浮游植物功能类群在夏季、秋季和冬季占比较高,M、LO、P浮游植物功能类群在夏季和秋季具有比较高的丰度比例,Y功能群和D功能群仅在冬季占比较高。
S1站点(河口区):春季主要功能群为B、S1和J;夏季主要功能群为TC、J、S1和M;秋季主要功能群为M、S1和TC;冬季主要功能群为Y、P和B。1号点的以湖泊假鱼腥藻为主的S1功能群在春季、夏季和秋季都处于优势地位。4号点(过渡区):春季主要功能群为B、J和S1;夏季主要功能群为P、M、TC和S1;秋季主要功能群为S1、K、H1和J;冬季主要功能群为H1、B、Y和P。9号点(湖泊区):春季主要功能群为B和S1;夏季主要功能群为M、P和S1;秋季主要功能群为S1和TC;冬季主要功能群为Y、H1、B和LO。
2.6 浮游植物功能群与环境因子的关系环境因子对浮游植物功能群的组成及变化有重要的影响,即水体中的营养盐、温度和光照等环境因子都会对浮游植物功能群产生重要的作用[20]。陆欣鑫等[21]研究表明,水体温度会影响浮游植物功能群的分组。
对功能群与环境因子的CCA分析显示:春季,前两个排序轴的特征值为0.253和0.109,水温与K和J浮游植物功能群呈正相关关系(P < 0.05);夏季,前两个排序轴的特征值为0.474和0.299,水温与S1、H1和M浮游植物功能群显著正相关(P < 0.05);与P功能群呈显著负相关关系(P < 0.05);秋季,水温与K、H1和J呈正相关关系;冬季,水温与D和TC功能群呈负相关关系。由此可以得出千岛湖水温的变化可以影响浮游植物功能群的组成[22-23]。
浮游植物的生长代谢速率与总磷和总氮等营养盐有密切的关系,营养盐是浮游植物生存的必要条件[24]。许多研究[25-26]也表明,营养盐的浓度是影响浮游植物群落结构演替的重要因素。本研究得出结论:春季,总氮与B浮游植物功能群呈显著正相关关系(P < 0.05),总磷与J功能群显著正相关(P < 0.05);总氮和总磷在秋季与M功能群显著正相关(P < 0.05);冬季,总氮与D和TC功能群显著正相关(P < 0.05),总磷与H1和LO呈现负相关关系(P < 0.05)。

|
图 11 千岛湖四季浮游植物功能群与环境因子的CCA分析 Fig. 11 CCA analysis of phytoplankton functional groups and environmental factors in four seasons of Qiandao Lake |
本研究鉴定出的浮游植物主要由绿藻、硅藻和隐藻组成,与历史研究一致[28],共鉴定出10种优势种,主要优势种为小环藻、克罗钝脆杆藻、颗粒直链藻、尖尾蓝隐藻等常见的温带小型浮游植物。在长江中下游硅藻数据库中,这些浮游种类均有很高的总磷适宜值,在欧洲一些富营养浅水湖泊中也很常见[29],并被作为富营养水体的主要指示种[30],其次是蓝藻门和绿藻门。与2013—2014年的浮游植物组成相比,水华鱼腥藻、假鱼腥藻、克罗钝脆杆藻、颗粒直链藻和尖尾蓝隐藻等在所研究年份均构成优势种[28],优势种在年度变化上既有延续又有更替。自从千岛湖实施非经典生物操纵以来,至2004年,生物密度出现明显的减少趋势,且下降到最低值[31-32]。2007—2010年浮游植物生物密度增加,但浮游植物生物量却没有升高,这表明千岛湖小型浮游植物的比例有所上升,这可能与鲢鳙的滤食作用及营养盐的年度变化有关[33]。
3.2 千岛湖浮游植物的时空分布从物种数的季节变化上看,千岛湖秋季最多(147种),春季最少(110种)。春季群落以硅藻类(如梅尼小环藻、短小曲壳藻等)为主,同时也会有尖尾蓝隐藻与水华束丝藻等其他藻类。但尖尾蓝隐藻主要出现在下游的S9站点,可能与S9站点位于水库下游的湖泊区,水流相对较小、且其营养盐含量较低有关;初春季节水温较低,也尚未形成温跃层,因此硅藻常成为优势种类;春末夏初,蓝藻生物量也逐渐升高,这既是浮游植物对水温升高的响应,同时也是对春季梅雨季节带入的大量外源性营养物的响应[28]。夏秋两季随着水温的进一步升高,营养盐丰富,蓝藻逐渐占据优势,主要优势种有水华鱼腥藻和湖泊鞘丝藻等,但硅藻中的某些种类如小环藻等仍占据着一定的优势[28]。
由于水流速度差异,浮游植物生物量和生物密度存在着明显的空间差异。在S1站点,硅藻门的生物量比其他站点高,主要是因为河流区水流湍急,悬浮颗粒多等特点[34-35],更适宜于硅藻的生长[10],因此S1站点的硅藻较其他站点高。浮游植物种类数从河流区经过渡区至湖泊区呈降低的趋势(即S1>S4>S9),S1采样点的物种数在两年的研究期间均较高,主要由于:(1)S1站点位于千岛湖的上游,为湖泊的进水口,受河流上游生活、农田污水的影响较大,因此S1站点营养盐水平比较高[36],浮游植物组成丰富;(2)由于S1站点为河口区,水动力较强,流速相对较快,不利于同一种浮游植物的积累[10],不会有优势度高的种类存在。3号点和8号点的生物量高于1、4和9号点,主要是由于蓝藻门浮游植物的生物量显著高于其他站点(P < 0.05)。3号点处于汇水区,蓝藻生物量相对较高,而4号点2015年的生物量却处在最低水平。总的来说,就生物量而言,1号点主要以硅藻为主,3、4号点主要是以硅藻、蓝藻和绿藻为主。8号点和9号点浮游植物群落结构相似。
千岛湖浮游植物生物量和生物密度大致会随水层降低而波动减少,可能受温度和光照的影响,水体的深度越深,温度越低,光照强度越弱,水体中的浮游植物就会越少[37]。从浮游植物种数来看,绿藻门在各样点各月份的种类数均很高。从浮游植物生物密度和生物量来看,夏季以蓝藻为主,春季以硅藻为主。
3.3 浮游植物功能群对环境因子的响应S1功能群主要以假鱼腥藻为主,蓝藻门生长繁殖最适温度在25~35 ℃,因夏季温度较高,使得蓝藻生长速度加快,更具有竞争优势;在夏季,多数水库以S1、H1和M浮游植物功能群为主,这几个功能群均适宜生活在富营养水体中,S1适宜生活在浑浊的、环境稳定的水体中;H1功能类群适宜生活在含氮量较低的、富营养的和分层的水体环境中[9]。这3种浮游植物功能群均以蓝藻为主,在国内外许多水库(或湖泊)中均有出现,且在水华易爆发时期为主要浮游植物功能群组合[38-41],例如巴西水库的浮游植物优势功能群包括M和H1[42],贵州三板溪水库的优势浮游植物功能群有H1功能群[43]。也有研究[44]表明,当S1、H1和M浮游植物功能群出现较少时,说明该水体浮游植物功能群的多样性在降低。浮游植物功能类群的组成及其变化受理化因子和生物因子的综合影响[45],而营养盐的作用经常比其他因子更为重要[46],因此研究浮游植物功能群的组成及演替规律,对千岛湖以及其他水域的生态特征及水质评价具有重要意义。
| [1] |
李大命, 于洋, 张彤晴, 等. 太湖春季和秋季浮游植物的光合作用活性[J]. 环境科学研究, 2014, 27(8): 848-856. LI D M, YU Y, ZHANG T Q, et al. Photochemical activity of phytoplankton in Taihu Lake in spring and autumn[J]. Research of Environmental Sciences, 2014, 27(8): 848-856. |
| [2] |
李秋华, 何伟添, 陈椽. 澳门湿地浮游植物群落特征[J]. 植物生态学报, 2009, 33(4): 689-697. LI Q H, HE W T, CHEN C. Characteristics of the phytoplankton community in wet-lands of Macao[J]. Chinese Journal of Plant Ecology, 2009, 33(4): 689-697. DOI:10.3773/j.issn.1005-264x.2009.04.007 |
| [3] |
KALFF J.湖沼学: 内陆水生态系统[M].古滨河, 刘正文, 李宽意, 等, 译.北京: 高等教育出版社, 2011. KALFF J. Limnology: inland water ecosystems[M]. GU B H, LIU Z W, LI K Y, et al, trans. Beijing: Higher Education Press, 2011. |
| [4] |
程曦, 李小平. 淀山湖氮磷营养物20年变化及其藻类增长响应[J]. 湖泊科学, 2008, 20(4): 409-419. CHENG X, LI X P. 20-year variations of nutrients (N and P) and their impacts on algal growth in Lake Dianshan, China[J]. Journal of Lake Sciences, 2008, 20(4): 409-419. DOI:10.3321/j.issn:1003-5427.2008.04.002 |
| [5] |
刘建康. 高级水生生物学[M]. 北京: 科学出版社, 1999. LIU J K. Higher aquatic biology[M]. Beijing: Science Press, 1999. |
| [6] |
韩希福, 王荣. 海洋浮游动物对浮游植物水华的摄食与调控作用[J]. 海洋科学, 2001, 25(10): 31-33. HAN X F, WANG R. The grazing impact and regulation effects of zooplankton on phytoplankton bloom[J]. Marine Sciences, 2001, 25(10): 31-33. DOI:10.3969/j.issn.1000-3096.2001.10.011 |
| [7] |
陈济丁, 任久长, 蔡晓明. 利用大型浮游动物控制浮游植物过量生长的研究[J]. 北京大学学报(自然科学版), 1995, 31(3): 373-382. CHEN J D, REN J C, CAI X M. Studies on control of excessive phytoplankton growth by macrozooplankton[J]. Acta Scientiarum Naturalium Universitatis Pekinensis, 1995, 31(3): 373-382. DOI:10.3321/j.issn:0479-8023.1995.03.001 |
| [8] |
WALTERS C J, KRAUSE E, NEILL W E, et al. Equilibrium models for seasonal dynamics of plankton biomass in Four Oligotrophy Lakes[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1987, 44(5): 1002-1017. DOI:10.1139/f87-118 |
| [9] |
PADISÁK J, CROSSETTI L O, NASELLI-FLORES L. Use and misuse in the application of the phytoplankton functional classification:a critical review with updates[J]. Hydrobiologia, 2009, 621(1): 1-19. DOI:10.1007/s10750-008-9645-0 |
| [10] |
REYNOLDS C S. Ecology of phytoplankton[M]. Cambridge: Cambridge University Press, 2006: 38-90.
|
| [11] |
张雅燕, 吴志旭, 朱溆君. 千岛湖藻类及相关环境因子多元线性回归和鱼腥藻预测模型的建立[J]. 中国环境监测, 2002, 18(3): 37-41. ZHANG Y Y, WU Z X, ZHU X J. Model prediction of anabaena bloom using backward multiple regression between algae & related environmental factors in Thousand-island Lake[J]. Environmental Monitoring in China, 2002, 18(3): 37-41. DOI:10.3969/j.issn.1002-6002.2002.03.015 |
| [12] |
吴志旭, 刘明亮, 兰佳, 等. 新安江水库(千岛湖)湖泊区夏季热分层期间垂向理化及浮游植物特征[J]. 湖泊科学, 2012, 24(3): 460-465. WU Z X, LIU M L, LAN J, et al. Vertical distribution of phytoplankton and physico-chemical characteristics in the lacustrine zone of Xin'anjiang reservoir (Lake Qiandao) in subtropic China during summer stratification[J]. Journal of Lake Sciences, 2012, 24(3): 460-465. DOI:10.3969/j.issn.1003-5427.2012.03.019 |
| [13] |
兰佳, 吴志旭, 李俊, 等. 新安江水库浮游植物群落特征及影响因素分析[J]. 中国环境监测, 2014, 30(2): 40-46. LAN J, WU Z X, LI J, et al. Analysis of phytoplankton community and its preliminary influenced factors in Xin'an Jiang Reservoir[J]. Environmental Monitoring in China, 2014, 30(2): 40-46. DOI:10.3969/j.issn.1002-6002.2014.02.008 |
| [14] |
胡忠军, 孙月娟, 刘其根, 等. 浙江千岛湖深水区大型底栖动物时空变化格局[J]. 湖泊科学, 2010, 22(2): 265-271. HU Z J, SUN Y J, LIU Q G, et al. Temporal and spatial distribution of profundal macrozoobenthic community in Lake Qiandao, Zhejiang, China[J]. Journal of Lake Sciences, 2010, 22(2): 265-271. |
| [15] |
董静, 李艳晖, 李根保, 等. 东江水系浮游植物功能群季节动态特征及影响因子[J]. 水生生物学报, 2013, 37(5): 836-843. DONG J, LI Y H, LI G B, et al. Seasonal dynamics characteristics and affecting physical factors of phytoplankton functional groups in Dongjiang River[J]. Acta Hydrobiologica Sinica, 2013, 37(5): 836-843. |
| [16] |
毕列爵, 胡征宇. 中国淡水藻志(第八卷)-绿藻门:绿球藻目(上)[M]. 北京: 科学出版社, 2004. BI L J, HU Z Y. Flora algarum sinicarum aquae dulcis[M]. Beijing: Science Press, 2004. |
| [17] |
朱浩然. 中国淡水藻志(第二卷):色球藻纲[M]. 北京: 科学出版社, 1991. ZHU H R. Flora algarum sinicarum aquae dulcis[M]. Beijing: Science Press, 1991. |
| [18] |
SALMASO N, PADISÁK J. Morpho-functional groups and phytoplankton development in two deep lakes (Lake Garda, Italy and Lake Stechlin, Germany)[J]. Hydrobiologia, 2007, 578(1): 97-112. DOI:10.1007/s10750-006-0437-0 |
| [19] |
TOLOTTI M, THIES H, NICKUS H, et al. Temperature modulated effects of nutrients on phytoplankton changes in a mountain lake[J]. Hydrobiologia, 2012, 698(1): 61-75. DOI:10.1007/s10750-012-1146-5 |
| [20] |
TILMAN D, KILHAM S S. Phosphate and silicate growth and uptake kinetics of the diatoms Asterionella formosa and Cyclotella meneghiniana in batch and semicontinuous culture[J]. Journal of Phycology, 1976, 12(4): 375-383. |
| [21] |
陆欣鑫, 刘妍, 范亚文. 呼兰河湿地夏秋两季浮游植物功能分组演替及其驱动因子[J]. 生态学报, 2014, 34(5): 1264-1273. LU X X, LIU Y, FAN Y W. Relationships between environmental variables and seasonal succession in phytoplankton functional groups in the Hulan River Wetland[J]. Acta Ecologica Sinica, 2014, 34(5): 1264-1273. |
| [22] |
严文逸, 王绍祥, 顾静, 等. 青草沙水库浮游藻类群落组成及其与环境因子的关系[J]. 上海海洋大学学报, 2017, 26(1): 75-84. YAN W Y, WANG S X, GU J, et al. Composition of planktonic algae community and its relationship with environmental factors in Qingcaosha Reservoir[J]. Journal of Shanghai Ocean University, 2017, 26(1): 75-84. |
| [23] |
邓建明, 蔡永久, 陈宇炜, 等. 洪湖浮游植物群落结构及其与环境因子的关系[J]. 湖泊科学, 2010, 22(1): 70-78. DENG J M, CAI Y J, CHEN Y W, et al. Structure of phytoplankton community and its relationship with environment factors in Lake Honghu[J]. Journal of Lake Sciences, 2010, 22(1): 70-78. |
| [24] |
LELAND H V. Distribution of phytobenthos in the Yakima River basin, Washington, in relation to geology, land use and other environmental factors[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1995, 52(5): 1108-1129. DOI:10.1139/f95-108 |
| [25] |
YUSOFF F M, ZUBAIDAH M S, MATIAS H B, et al. Phytoplankton succession in intensive marine shrimp culture ponds treated with a commercial bacterial product[J]. Aquaculture Research, 2002, 33(4): 269-278. DOI:10.1046/j.1355-557x.2002.00671.x |
| [26] |
NUCCIO C, MELILLO C, MASSI L, et al. Phytoplankton abundance, community structure and diversity in the eutrophicated Orbetello lagoon (Tuscany) from 1995 to 2001[J]. Oceanologica Acta, 2003, 26(1): 15-25. DOI:10.1016/S0399-1784(02)00007-5 |
| [27] |
DOMINGUES R B, SOBRINO C, GALVÃO H. Impact of reservoir filling on phytoplankton succession and cyanobacteria blooms in a temperate estuary[J]. Estuarine, Coastal and Shelf Science, 2007, 74(1/2): 31-43. |
| [28] |
胡忠军, 莫丹玫, 周小玉, 等. 千岛湖浮游植物群落结构时空分布及其与环境因子的关系[J]. 水生态学杂志, 2017, 38(5): 46-54. HU Z J, MO D M, ZHOU X Y, et al. Spatial and seasonal distribution of phytoplankton in Qiandao Lake and relationship with environmental factors[J]. Journal of Hydroecology, 2017, 38(5): 46-54. |
| [29] |
BENNION H. A diatom-phosphorus transfer function for shallow, eutrophic ponds in southeast England[J]. Hydrobiologia, 1994, 275-276(1): 391-410. DOI:10.1007/BF00026729 |
| [30] |
董旭辉, 羊向东, 王荣. 长江中下游地区湖泊富营养化的硅藻指示性属种[J]. 中国环境科学, 2006, 26(5): 570-574. DONG X H, YANG X D, WANG R. Diatom indicative species of eutrophication of the lakes in the middle and lower reach regions of Yangtze River[J]. China Environmental Science, 2006, 26(5): 570-574. DOI:10.3321/j.issn:1000-6923.2006.05.014 |
| [31] |
刘其根.千岛湖保水渔业及其对湖泊生态系统的影响[D].上海: 华东师范大学, 2005. LIU Q G. Aquatic environmental protection oriented fishery in Lake Qiandaohu and its influences on lake ecosystem[D]. Shanghai: East China Normal University, 2005. |
| [32] |
韩伟明, 胡水景, 金卫, 等. 千岛湖水环境质量调查与保护对策[J]. 湖泊科学, 1996, 8(4): 337-344. HAN W M, HU S J, JIN W, et al. Investigation and protection countermeasures of water environmental quality in Qiandao Lake[J]. Journal of Lake Sciences, 1996, 8(4): 337-344. |
| [33] |
VÖRÖS L, OLDAL I, PRÉSING M, et al. Size-selective filtration and taxon-specific digestion of plankton algae by silver carp (Hypophthalmichthys molitrix Val.)[J]. Hydrobiologia, 1997, 342/343: 223-228. DOI:10.1023/A:1017039423485 |
| [34] |
杨亮杰, 余鹏飞, 竺俊全, 等. 浙江横山水库浮游植物群落结构特征及其影响因子[J]. 中国应用生态学报, 2014, 25(2): 569-576. YANG L J, YU P F, ZHU J Q, et al. Community structure characteristics of phytoplankton and related affecting factors in Hengshan Reservoir, Zhejiang, China[J]. Chinese Journal of Applied Ecology, 2014, 25(2): 569-576. |
| [35] |
盛海燕, 虞左明, 刘明亮, 等. 千岛湖冬季浮游植物光合活性[J]. 生态学杂志, 2014, 33(2): 365-372. SHENG H Y, YU Z M, LIU M L, et al. The photosynthetic activity of phytoplankton in Qiandao Lake in winter[J]. Chinese Journal of Ecology, 2014, 33(2): 365-372. |
| [36] |
吕唤春, 王飞儿, 陈英旭, 等. 千岛湖水体叶绿素a与相关环境因子的多元分析[J]. 应用生态学报, 2003, 14(8): 1347-1350. LV H C, WANG F E, CHEN Y X, et al. Multianalysis between chlorophyll-a and environmental factors in Qiandao Lake water[J]. Chinese Journal of Applied Ecology, 2003, 14(8): 1347-1350. DOI:10.3321/j.issn:1001-9332.2003.08.030 |
| [37] |
潘继征, 熊飞, 李文朝, 等. 抚仙湖浮游植物群落结构、分布及其影响因子[J]. 生态学报, 2009, 29(10): 5376-5385. PAN J Z, XIONG F, LI W C, et al. Structure, distribution and its impact factors of phytoplankton community in Fuxian Lake[J]. Acta Ecologica Sinica, 2009, 29(10): 5376-5385. DOI:10.3321/j.issn:1000-0933.2009.10.024 |
| [38] |
PADISÁK J, BARBOSA F, KOSCHEL R, et al. Deep layer cyanoprokaryota maxima in temperate and tropical lakes[M]//KOSCHEL R, ADAMS D. Lake Stechlin. An Approach to Understanding an Oligotrophic Lowland Lake. Archiv für Hydrobiologoe/Advances in Limology, 2003, 58: 175-199.
|
| [39] |
张怡, 胡韧, 肖利娟, 等. 南亚热带两座不同水文动态的水库浮游植物的功能类群演替比较[J]. 生态环境学报, 2012, 21(1): 107-117. ZHANG Y, HU R, XIAO L J, et al. Comparative analysis of succession of the phytoplankton functional groups in two reservoirs with different hydrodynamics in southern China[J]. Ecology and Environmental Sciences, 2012, 21(1): 107-117. DOI:10.3969/j.issn.1674-5906.2012.01.020 |
| [40] |
黄享辉, 胡韧, 雷腊梅, 等. 南亚热带典型中小型水库浮游植物功能类群季节演替特征[J]. 生态环境学报, 2013, 22(2): 311-318. HUANG X H, HU R, LEI L M, et al. Seasonal succession of phytoplankton functional groups in typical small and medium-sized reservoirs in southern China[J]. Ecology and Environmental Sciences, 2013, 22(2): 311-318. DOI:10.3969/j.issn.1674-5906.2013.02.023 |
| [41] |
STEVIĆ F, MIHALJEVIĆ M, ŠPOLJARIĆ D. Changes of phytoplankton functional groups in a floodplain lake associated with hydrological perturbations[J]. Hydrobiologia, 2013, 709(1): 143-158. DOI:10.1007/s10750-013-1444-6 |
| [42] |
GEMELGO M C, MUCCI J L, NAVASPEREIRA D. Population dynamics:seasonal variation of phytoplankton functional groups in Brazilian reservoirs (Billings and Guarapiranga, Sao Paulo)[J]. Brazilian Journal of Biology, 2009, 69(4): 1001-1013. DOI:10.1590/S1519-69842009000500004 |
| [43] |
黄国佳, 李秋华, 陈椽, 等. 贵州高原三板溪水库浮游植物功能群时空分布特征[J]. 环境科学学报, 2015, 35(2): 418-428. HUANG G J, LI Q H, CHEN C, et al. Phytoplankton functional groups and their spatial and temporal distribution characteristics in Sanbanxi Reservoir, Guizhou Province[J]. Acta Scientiae Circumstantiae, 2015, 35(2): 418-428. |
| [44] |
BORICS G, TÍTHMÉRÉSZ B, LUKÁCS B A, et al. Functional groups of phytoplankton shaping diversity of shallow lake ecosystems[J]. Hydrobiologia, 2012, 698(1): 251-262. DOI:10.1007/s10750-012-1129-6 |
| [45] |
刘歆璞, 张玮, 王丽卿, 等. 青草沙水库浮游植物群落结构及其与环境因子的关系[J]. 上海海洋大学学报, 2015, 24(4): 532-543. LIU X P, ZHANG W, WANG L Q, et al. Structure of phytoplankton community and its relationship with environmental factors in Qingcaosha Reservoir[J]. Journal of Shanghai Ocean University, 2015, 24(4): 532-543. |
| [46] |
秦雪, 徐宾铎, 杨晓改, 等. 黄河口及其邻近水域夏季浮游植物群落结构及其与环境因子的关系[J]. 水产学报, 2016, 40(5): 711-720. QIN X, XU B D, YANG X G, et al. Phytoplankton community structure and its relationship with environmental variables in summer in the Yellow River Estuary and its adjacent waters[J]. Journal of Fisheries of China, 2016, 40(5): 711-720. |
 ,
HU Zhongjun1,2,3,
SHI Xianhe4,
REN Liping5,
PAN Jiayong5,
CUI Liang1,2,3,
CHEN Laisheng5,
HE Guangxi5,
LIU Qigen1,2,3
,
HU Zhongjun1,2,3,
SHI Xianhe4,
REN Liping5,
PAN Jiayong5,
CUI Liang1,2,3,
CHEN Laisheng5,
HE Guangxi5,
LIU Qigen1,2,3
2. Centre for Research on Environmental Ecology and Fish Nutrition(CREEFN) of the Ministry of Agriculture, Shanghai Ocean University, Shanghai 201306, China;
3. Shanghai Collaborative Innovation Center for Aquatic Animal Genetics and Breeding, Shanghai Ocean University, Shanghai 201306, China;
4. Heng'an School, Hefei 231137, Anhui, China;
5. Hangzhou Qiandao Lake Group Co. Ltd, Hangzhou 311700, Zhejiang, China