 2018,
Vol. 27
2018,
Vol. 27



2. 大洋渔业资源可持续开发省部共建教育部重点实验室, 上海 201306;
3. 国家远洋渔业工程技术研究中心, 上海 201306;
4. 农业部大洋渔业开发重点实验室, 上海 201306
柔鱼(Ommastrephes bartramii)作为一种大洋性头足类,广泛分布于三大洋,目前柔鱼资源规模性开发海域主要集中在北太平洋[1]。该资源最早由日本于1974年进行开发,而后其他国家和地区开始对其进行大规模的商业性开发利用[2]。我国自1993年开发和利用柔鱼资源以来,主要以西北太平洋柔鱼冬春生群体作为传统的捕捞对象;作业时间为每年6至12月,主要汛期为8至11月;捕捞海域分布较广,主要集中于38°N~45°N,140°W~165°W海域[3]。国内外学者对北太平洋柔鱼种群结构、年龄生长[4]、索饵洄游[5]、产卵洄游[6-8]及其渔场时空分布与海洋环境之间的关系[6, 9-11]等方面做了大量的研究。对于一年生的柔鱼,本文假设其因遗传而具有固定的洄游路径。柔鱼对于生存的海洋环境变化极为敏感[12-15],年间的洄游路径极易受到海洋环境的影响而发生改变[16]。因此,准确探究柔鱼洄游路径及其路径变动与海洋环境之间的关系对于实际的渔业生产具有重大意义。
柔鱼洄游路径通常可以使用渔场重心来表示[17-18]。渔场重心是表征渔场位置的重要参数,与环境要素和生物特征等因素有关,常用于描述渔场空间位置的变动,作为渔业资源和鱼类生态学研究的基础,亦可以用来确定中心渔场[19-20]和表征鱼群在渔场分布特征和变化规律[21-23]。以往的研究中,渔场重心计算采用的都是固定物体重心计算方法[20, 22-23],然而柔鱼具有洄游特性,并不等同于固定物体。因此,固定物体重心法对于实际渔业生产上的指导意义有失精度,使渔场重心无法准确表示鱼群分布特征[24]。反之,如果将整个渔场按照“经度×纬度”的方式划分成多个相连海域,分别计算渔场重心,当划分海域较多时,由于柔鱼作为生态机会主义者,渔场分布极易受环境因素影响而发生变化,故年间渔场重心的预报精度又会大幅下降。且实际生产更需要精确地指导,对应的时间、对应的地点和对应的鱼群资源丰度等,预报渔场重心过多,同样缺乏实际的指导意义。
鱿钓渔业是根据柔鱼集群和趋光性等特性所建立起来的高效灯光诱捕作业方式。实际生产中,因为柔鱼的集群而相应地出现鱿钓渔船之间的聚集,往往1°~2°海域范围内的渔船数量多达上百艘,占中国鱿钓船队的30%~80%。通过提升渔场重心在时间和空间上精度并分析其变化规律,可以为柔鱼渔业提供准确的生产信息服务。
1 材料与方法 1.1 材料(1) 渔业数据来自上海海洋大学鱿钓技术组,作业范围为35°N~45°N,140°E~165°E,即为我国鱿钓船队在西北太平洋海域的传统作业渔场,捕捞对象主要是柔鱼冬春生群体。时间跨度为2004—2015年,为尽可能排除渔获物中其他群体的影响,选取西北太平洋冬春生群体的主要渔汛期:8—11月。空间分辨率为0.25°×0.25°。数据包括日期、经度、纬度、日产量,作业船数。
(2) 环境数据包括海表面温度(Sea Surface Temperature, SST)、海表面盐度(Sea Surface Salinity, SSS)和叶绿素浓度(chlorophyll concentration, Chl-a),时间范围为2004—2015年8—11月,数据来源于Ocean-watch网站(http://oceanwatch.pifsc.noaa.gov/las/servlets/dataset),数据范围为35°N~45°N,140°E~165°E。另外本研究中用到的尼诺转化指数(Trans-Niño index, TNI)来源于esrl网站(http://www.esrl.noaa.gov/psd/data/climateindices/List)。尼诺转化指数用于指代Nino 1+2和Nino 4海区的温差,当差值较大时,则定义为中太平洋厄尔尼诺。
1.2 方法 1.2.1 柔鱼洄游路径分析根据柔鱼的集群性、鱿钓渔业所采用的灯光诱鱼技术以及在实际生产中渔船的聚集性,可以用高产海域的(柔鱼洄游聚集海域)的渔场重心来指代柔鱼的洄游分布。虽然使用物理重心指代整个渔场的渔场重心可以表征柔鱼种群分布整体的变动趋势,但对于实际的渔业生产指导却有失准确性;根据物理重心计算公式,当所计算海域足够小时且相对于整个渔场可以看做一个点时,渔场重心基本可以表示该海域内鱼群的具体分布。因此,如何选定局部海域的大小,以提升在时间和空间上的精度是本研究需要考虑的因素。局部海域过小,该海域内可能的渔获量也会减少;局部海域过大,渔场重心计算结果精度降低;故首先定义经度、纬度0.25°×0.25°为一个渔区,统计每个渔区内的产量,单位为t/d。逐月统计每个渔区的产量,筛选出各月份所有渔区中产量最高的渔区,结合柔鱼的洄游能力以及鱿钓渔船的航行能力,以该渔区的中心为圆心,1°为半径,将所有包含在内的渔区统一纳入局部海域渔场,并将该海域的渔场重心定义为柔鱼的洄游重心G1,并依次选出G2和G3,则G1、G2和G3分别代表该月份柔鱼聚群洄游的重心,该重心1°海域内为柔鱼的洄游范围,并同时计算每个重心所包含渔区的产量总和。依据公式(1)和(2)来计算G1、G2、和G3。同时计算整个西北太平洋柔鱼渔场每月的渔场重心。
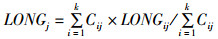 (1)
(1)
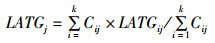 (2)
(2)
式中:LONGj重心经度位置;LATGj为重心纬度位置;Cij为i渔区,j月的产量;LONGij为渔区i中心点的经度;LATGij为渔区i中心点的纬度;k为渔区的总个数。
1.2.2 洄游重心的产量占比和离散度按上述方法筛选出的三个洄游重心,分别计算其产量占比:即3个海域内的产量的总和占该月份所有渔获产量的比值,以此来判断洄游重心这一概念的提出是否具有实际意义,即是否可以用来指代柔鱼种群的洄游位置,若三个洄游重心海域内的产量总和较低,则该洄游重心不足以代表柔鱼种群整体的洄游路径。因此,通过筛选出的洄游重心的产量占比来判断洄游重心的选择是否具有合理性。
离散度:以该月份整个西北太平洋海域柔鱼渔场的渔场重心作为参考点,按照公式1~3分别计算每月份柔鱼洄游重心G1、G2和G3与渔场重心之间的距离,利用该距离与各洄游重心海域内产量占比的乘积作为洄游重心离聚的判定指标。依据公式(3)计算出每月份柔鱼洄游重心的离散度,进而分析柔鱼洄游重心的年间变化与海洋气候环境之间的关系。
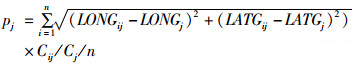 (3)
(3)
式中:pj为j月的离散度;LONGij为j月Gi的经度,LONGj为j月渔场重心的经度;LATGij为j月Gi纬度,LATGj为j月渔场重心的纬度;Cij为j月Gi洄游重心海域内所有渔区产量总和,Cj为j月渔场产量总和,n为洄游重心的数量,本研究中n取值为3。
1.2.3 洄游重心的系统聚类分析(Hierarchical cluster analysis)依据上述方法,可以根据2004—2015年12年产量数据,计算每月的洄游重心G1, G2和G3及其产量占比,按照月份和洄游重心种类进行分别聚类分析。本研究使用卡方距离,可变类平均法进行聚类分析;在聚类结果中类别数的确定采用χ2统计量,即以方差分析统计学意义最大为原则。
根据聚类分析的结果,以同一类别平均中心距离最小作为类别的选取依据,计算该类别内所有样本的经、纬度重心,在8—11月四个月中,分别得出唯一经、纬度重心的柔鱼洄游重心G1*、G2*和G3*,将聚类后的洄游重心及其1°为半径的海域定义为柔鱼在该月份的洄游海域,按照时间顺序的各月份柔鱼洄游海域连结为柔鱼的洄游路径。
1.2.4 洄游重心年间变动趋势与海洋气候因子之间的关系通过聚类分析所得的固定洄游重心,将各月份洄游重心之间的重叠海域假设为柔鱼固有的洄游路径,再与通过渔获产量计算所得的洄游重心进行比较,比较其纬度南北向和经度东西向的偏移规律与海洋气候因子TNI之间的关系。
1.2.5 神经网络模型预测洄游重心误差反向传播网络(Error Backpropagation Network, BP)属于多层前向神经网络[25],采用误差方向传播的监督算法,能够学习和存储大量的模式映射关系,已被广泛的应该用于各个领域[26-27]。本研究中,以各月份的柔鱼洄游备选路径(G1*、G2*和G3*)分别作为输入因子,结合洄游重心海域环境因子SST、SSS和Chl-a作为输入因子,以实际G1、G2和G3及其对应产量占比作为输出因子,进行训练。选取2015年的数据进行预测,可分别得出三个洄游重心及其对应资源丰度,再分别计算该洄游重心1°海域范围内的实际产量占比,比较预报的准确性。
2 结果 2.1 各月份柔鱼洄游重心变化和产量占比分析2004—2015年8—11月各月份的柔鱼洄游重心G1、G2和G3的具体分布和各重心海域产量占该月份总产量的比值见图 1。从图 1中可以看出,各个月份洄游重心G1的产量占比均高于G2和G3,8月份除2012和2013年外,G1和G2的产量占比差距并不是很大,差值在1%~25%之间,G3在2006~2013年和2015年的产量占比主要分布在10%~25%之间。从9月份开始至11月份,G1的产量占比明显增加:9月份除去2012—2014年,G1和G2的产量占比的差值在25%~66%之间,G3的占比最高到达15%,主要分布在5%~10%之间;10月份除去2012—2014年,G1和G2的产量占比差值最高达98%(2007年),主要分布在25%~40%之间,G3在2012年和2014年达到10%以上,其余年份都在1%~8%范围内;11月份2004—2007年和2015年,G1和G2的产量占比的差值在50%~90%之间,G3除2005年外,产量占比均低于10%,主要分布在1%~5%之间。从年份来看,2004—2007年和2015年均满足G1的产量占比随着月份增加而增加,G2和G3的产量占比随着月份逐渐减少。2009—2014年G1的产量占比同样也有该趋势,只是增加趋势相对平缓。

|
图 1 2004—2015年各月份柔鱼洄游重心分布及产量占比
Fig. 1 Monthly distribution of production percentage and migration gravity of O.bartramii in 2004 to 2015
|
从分布来看,8月份,G1、G2和G3分布相对均匀,G1主要分布在152°E~156.5°E,41°N~43.5°N之间;G2主要分布在151.5°E~156.5°E,41.5°N~43.5°N之间;G3主要分布在151.5°E~157.5°E,41°N~43.5°N之间。9月份G1分布比较集中,主要分布在154.5°E~157°E,42.5N°~44.5°N之间,相对8月份,分布范围向东移动2°左右,向北移动1°左右。G2和G3分布相对分散,主要分布在151°E~160°E,40.5°E~44°N之间。10月份G1分布主要集中在154°E~156.5°E,42.5°N~43.5°N之间,相对9月份更为集中,且维度方向范围向北缩小0.5°,G2和G3分布相对分散,主要分布在151.5°E~157.5°E,40°N~43° N之间。11月份G1分布范围相对南移,主要分布在153°E~155.5°E,41.5°N~43°N之间,相对于10月份,分布范围向西、向南偏移1°左右,G2和G3则主要分布在150°E~156°E,40.5°N~43°N之间。
2.2 柔鱼洄游重心产量占比及其离散度分析按上述方法,由图 2可以看出各个月份洄游重心G1、G2和G3的三个海域的产量总和占该月份总产量的比值,除去个别极值分布在50~60%之间外,其余均在70%~98%之间,表明三个渔场重心指代柔鱼的洄游重心具有代表性。其中8月主要分布在78%~92%之间;9月份主要分布在72%~98%之间,其中一半分布在90%以上;10月份则主要集中在80%~90%之间;11月份主要分布在95%~98%之间;从月份变化来看,三个海域的产量占比总和逐月增加。

|
图 2 2004—2015年各月份洄游重心产量总和占比
Fig. 2 Monthly production percentage and distribution range of migration gravity in 2004 to 2015
|
由图 3a可以看出,9—11月的波动趋势大致相同, 8月份波动幅度较大,产量占比相对不稳定。2008—2014年各月份的产量占比均出现较大的波动,其余年份产量占比变化趋势相对稳定。其余年份11月份产量占比最为稳定,且产量占比值较高,8月份波动较大,9月和10月变化趋势相对平缓。

|
图 3 2004—2015年各月份洄游重心产量总和占比及离散度年间变化
Fig. 3 Annual production percentage anddispersion degreevariation tendency range of monthly migration gravity in 2004 to 2015
|
由图 3b可以看出各月份洄游重心离散度的年间变化趋势,其中9月份和11月份波动较大,且在2008—2014年期间各月份离散度波动较大,且变化趋势也不一致,11月份波动幅度最大。其余年份波动趋势一致,变化范围较小。各月份洄游重心产量占比和各月份离散度的相关分析结果表明10月份(r=-0.55, P < 0.01)和11月份(r=-0.92, P < 0.01)洄游重心产量占比和离散度之间存在着明显的负相关,而8月份和9月份则没有明显的相关性。
2.3 系统聚类分析系统聚类分析采用可变类平均法和卡方距离分别对各月份的3个洄游重心进行聚类,根据χ2统计量确定聚类谱系图中类别数;从2004—2015年的样本中,依据上述类别选取依据,分别选取各月份洄游重心的聚类类别,并将该类别内样本作为正常年份柔鱼洄游重心分布,各类别包含年份分布如表 1所示。

|
表 1 系统聚类分析结果分布 Tab.1 The distribution of the results of the hierarchical cluster analysis |
经上述结果计算统计意义上的柔鱼洄游重心分布,按照本文假设,以此作为柔鱼未受海洋环境影响下的固定洄游路径。各月份G1*、G2*和G3*及其海域范围的分布如图 4所示。

|
图 4 固定洄游重心海域范围分布
Fig. 4 The distribution and range of the inherent migration gravity
|
结合以上柔鱼洄游重心产量占比和离散度的结果,可将种群洄游分布模式分为两种,其中2004年至2007年和2015年分为一类,2008年至2014年可归为另一类。不同之处在于:类别1中8月份三个渔场重心的产量占比相对均匀,其余年份G1产量占比突出明显;9月份至11月份,类别1中G1产量占比明显增加,且随着月份的增加逐渐增大,G2和G3占比减少。其余年份的G1产量增加不明显。将以上得到的洄游重心的结果与各月份对应的尼诺转化指数进行相关性分析,结果如表 3所示。8—10月:2008—2014年,洄游重心纬度上的变化趋势与TNI之间呈现显著正相关,经度上并无相关性;其余年份仅9月份经度上呈现显著负相关。11月份:2008—2014年,相关性并不显著,其余年份却呈现显著正相关性。
2.5 洄游重心的预测依据上述所得柔鱼洄游重心,结合SST、SSS和Chl-a等海洋环境因素,作为神经网络的输入因子,每月份统计上的洄游重心G1、G2和G3作为输出因子。网络设计的参数为:隐藏层神经元根据经验公式取9,输出层神经元为3,学习速率0.1,动量参数0.5,输入层与隐含层,隐含层与输出层神经元之间的传递函数分别是S型正切函数tansig/线性函数purelin;网络训练的终止参数为:最大训练次数为1 000,最大误差定为0.001。通过计算预测重心与柔鱼标准洄游重心之间的距离来评定预报精度。
神经网络模型结构采用5-9-3进行训练。
利用上述方法构建的预测模型,以2004—2014年的样本作为拟合样本,以2015年的样本数据作为预测样本,各月份预测洄游重心如图 5所示。

|
图 5 预测洄游重心海域分布范围
Fig. 5 The distribution and range of the forecasting migration gravity
|
以上预测洄游重心海域产量占比如图 6所示:8月份和9月份预测海域产量占比总和为64%和68%;10月份和11月份预报产量占比总和为83%和89%,明显高于8、9月份。

|
图 6 预测海域产量总和占比
Fig. 6 Total production percentage in the forecasting sea area
|
柔鱼渔场重心的计算多采用固定物体中心计算方法[24],整个渔场所得的渔场重心,根据其公式意义可以表示鱼群整体的空间变化,而很难精确的计算出柔鱼具体的洄游重心,更难揭示柔鱼年间洄游路径之间的变化差异,且渔场海域越大,洄游路径的预测精度越低。本研究创新性的提出了洄游重心,根据柔鱼集群特性和鱿钓渔业的作业方式,利用产量指代柔鱼聚集程度,从而确定柔鱼在相应时间内出现的海域。从整个渔场中筛选出三个洄游重心,即G1、G2和G3,以1°为半径的洄游重心海域作为整个渔场的洄游重心。从图 2中可以看出,各月份洄游重心产量占比总和除个别值外,均稳定在70%~98%之间,因此,三个洄游重心可以代表整个渔场的鱼群的洄游路径变化的趋势。图 1和图 3可以看出,8月,各洄游重心产量占比相对均匀,柔鱼分布相对分散;9月到10月,G1产量占比开始逐渐增加,其余洄游重心产量占比开始减少,柔鱼分布相对8月开始聚集;11月,G1产量占比增大明显,柔鱼分布较为集中。此现象的成因可能跟柔鱼生活史相关。8月末,柔鱼向北或东北洄游进入35°N~40°N黑潮和亲潮交汇区,柔鱼这一移动路线主要是和黑潮暖水系分支方向有关[28-30]。9—10月性未成熟和性成熟的柔鱼主要分布在40°N~46°N、亲潮前锋区和周围海域,主要为索饵洄游期,分布相对分散。11月,柔鱼成长到性成熟的高峰期,随着亲潮冷水域的扩展,开始向南进行产卵洄游[7-8, 31],洄游具有目的性,种群分布较为集中,洄游重心离散度的变化趋势和柔鱼生活史中的索饵洄游和产卵洄游阶段相对应。
通过聚类分析得出8—11月的固定洄游重心,其洄游重心随着月份的改变,出现不同的变化。在纬度上呈现南-北-南的变化趋势,经度上呈现西-东-西的变化趋势(表 2和图 4)。分析G1、G2和G3与固定洄游重心G1*、G2*和G3*在经度和纬度上的发生的偏移与尼诺转化指数之间的关系可以看出:2008—2014年,洄游重心纬度上出现显著的正相关,即TNI数值增加,各年的洄游重心较固定洄游重心出现向北偏移,而在经度上并未出现相关性。其余年份,8月至10月纬度上并未出现相关的变化趋势,11月份纬度上则出现明显的正相关性。综合上述结果可以看出,尼诺转化指数对柔鱼洄游重心的影响主要体现在纬度上,即随着Nino 1+2与Nino 4海区的温差的增加,而出现洄游重心向北移动的变化趋势。11月份表现最为明显,主要原因是产卵洄游对于环境的要求更为严格,更容易受到海洋环境的影响而发生趋利性洄游。

|
表 2 洄游重心的偏移和尼诺转化指数之间的关系 Tab.2 The relationship between the shift of the migration gravity and TNI |
预测结果中,利用神经网络模型,结合固定洄游重心海域的海洋环境因素,来预测该年份洄游重心可能会发生的偏移及其对应的产量。预测洄游重心G1、G2和G3分布随着月份的变化,纬度和经度上也大致呈现南-北-南和西-东-西的变化趋势。预测产量结果(图 6)显示,8月和9月预测洄游重心海域的产量占比为64%和68%,柔鱼处于索饵期,分布海域较大且分布相对分散。在10月和11月,柔鱼种群开始聚集,进行产卵洄游,分布海域相对集中,故预测洄游重心海域产量占比明显提高,分别为83%和89%,预测精度明显提高。本研究在系统分析后得出的柔鱼固定洄游重心海域,结合对应时期的海洋环境,可以通过神经网络模型计算出实际的洄游重心相对于固定洄游重心在经纬度上发生的偏移,从而对实际渔业生产提出指导,作业渔船根据该信息在提高渔获的同时减少海上航行所需的燃油成本。
本研究按照渔场重心,从整个渔场中筛选出三个渔场重心,并定义为洄游重心,各月份洄游重心的连接,构成柔鱼的洄游路径。结果可以较好的反应柔鱼洄游路径的变化和海洋环境之间的关系,同时也可以很好的预测柔鱼资源丰度的具体分布。因数据限制,本研究在时间上采用月份,空间上采用1°为半径的圆形海域来对柔鱼的洄游重心进行分析和预测。如果以周为时间单位,以30′作为洄游重心的半径,还可以进一步提高时间和空间精度,从而达到更为准确的预测效果。
| [1] |
MURATA M. Oceanic resources of squids[J]. Marine Behaviour and Physiology, 1990, 18(1): 19-71. DOI:10.1080/10236249009378779 |
| [2] |
陈新军, 陈峰, 高峰, 等. 基于水温垂直结构的西北太平洋柔鱼栖息地模型构建[J]. 中国海洋大学学报, 2012, 42(6): 52-60. CHEN X J, CHEN F, GAO F, et al. Modeling of habitat suitability of neon flying squid (Ommastrephes bartramii) based on vertical temperature structure in the northwestern Pacific Ocean[J]. Periodical of Ocean University of China, 2012, 42(6): 52-60. |
| [3] |
陈新军, 钱卫国, 刘必林, 等. 利用衰减模型评估柔鱼西部冬春生群体资源[J]. 海洋湖沼通报, 2008(2): 130-140. CHEN X J, QIAN W G, LIU B L, et al. An assessment of western winter-spring stock of neon flying squid, Ommastrephes bartramii, in the northwest Pacific ocean using the depletion model[J]. Transactions of Oceanology and Limnology, 2008(2): 130-140. |
| [4] |
YATSU A, MORI J. Early growth of the autumn cohort of neon flying squid, Ommastrephes bartramii, in the North Pacific Ocean[J]. Fisheries Research, 2000, 45(2): 189-194. DOI:10.1016/S0165-7836(99)00112-5 |
| [5] |
WATANABE H, KUBODERA T, ICHⅡ T, et al. Feeding habits of neon flying squid Ommastrephes bartramii in the transitional region of the central North Pacific[J]. Marine Ecology Progress Series, 2004, 266: 173-184. DOI:10.3354/meps266173 |
| [6] |
ICHⅡ T, MAHAPATRA K, SAKAI M, et al. Differing body size between the autumn and the winter-spring cohorts of neon flying squid (Ommastrephes bartramii) related to the oceanographic regime in the North Pacific:a hypothesis[J]. Fisheries Oceanography, 2004, 13(5): 295-309. DOI:10.1111/fog.2004.13.issue-5 |
| [7] |
ICHⅡ T, MAHAPATRA K, SAKAI M, et al. Life history of the neon flying squid:effect of the oceanographic regime in the North Pacific Ocean[J]. Marine Ecology Progress Series, 2009, 378: 1-11. DOI:10.3354/meps07873 |
| [8] |
MURATA M, NAKAMURA Y. Seasonal migration and diel vertical migration of the neon flying squid, Ommastrephes bartramii in the North Pacific[C]//OKUTANI T ed. Contributed papers to International Symposium on Large Pelagic Squids, for JAMARC's 25th Anniversary of Its Foundation. Tokyo: Japan Marine Fishery Resources Research Center, 1998,
|
| [9] |
YATSU A, WATANABE T, MORI J, et al. Interannual variability in stock abundance of the neon flying squid, Ommastrephes bartramii, in the North Pacific Ocean during 1979-1998:impact of driftnet fishing and oceanographic conditions[J]. Fisheries Oceanography, 2000, 9(2): 163-170. DOI:10.1046/j.1365-2419.2000.00130.x |
| [10] |
陈新军. 北太平洋(160°E~170°E)大型柔鱼渔场的初步研究[J]. 上海水产大学学报, 1999, 8(3): 197-201. CHEN X J. The preliminary study on fishing ground of large-sized Ommastrephe bartramii in North Pacific waters between 160°E and 170°E[J]. Journal of Shanghai Fisheries University, 1999, 8(3): 197-201. |
| [11] |
陈新军, 曹杰, 田思泉, 等. 表温和黑潮年间变化对西北太平洋柔鱼渔场分布的影响[J]. 大连水产学院学报, 2010, 25(2): 119-126. CHEN X J, CAO J, TIAN S Q, et al. Effect of inter-annual change in sea surface water temperature and Kuroshio on fishing ground of squid Ommastrephes bartramii in the Northwest Pacific[J]. Journal of Dalian Fisheries University, 2010, 25(2): 119-126. |
| [12] |
RODHOUSE P G. Managing and forecasting squid fisheries in variable environments[J]. Fisheries Research, 2001, 54(1): 3-8. DOI:10.1016/S0165-7836(01)00370-8 |
| [13] |
余为. 西北太平洋柔鱼冬春生群对气候与环境变化的响应机制研究[D]. 上海: 上海海洋大学, 2016. YU W. Response mechanism of winter-spring cohort of neon flying squid to the climatic and environmental variability in the Northwest Pacific Ocean[D]. Shanghai: Shanghai Ocean University, 2016. http://cdmd.cnki.com.cn/Article/CDMD-10264-1016912194.htm |
| [14] |
余为, 陈新军, 易倩. 不同气候模态下西北太平洋柔鱼渔场环境特征分析[J]. 水产学报, 2017, 41(4): 525-534. YU W, CHEN X J, YI Q. Analysis of variations in the environmental conditions on the fishing ground of neon flying squid (Ommastrephes bartramii) in the Northwestern Pacific Ocean under different climate modes[J]. Journal of Fisheries of China, 2017, 41(4): 525-534. |
| [15] |
余为, 陈新军, 易倩, 等. 西北太平洋柔鱼传统作业渔场资源丰度年间差异及其影响因子[J]. 海洋渔业, 2013, 35(4): 373-381. YU W, CHEN X J, YI Q, et al. Annual difference of abundance index and its influencing factors of Ommastrephes bartramii in traditional fishing grounds in the Northwest Pacific[J]. Marine Fisheries, 2013, 35(4): 373-381. |
| [16] |
陈新军, 田思泉, 许柳雄. 西北太平洋海域柔鱼产卵场和作业渔场的水温年间比较及其与资源丰度的关系[J]. 上海水产大学学报, 2005, 14(2): 168-175. CHEN X J, TIAN S Q, XU L X. Analysis on changes of surface water temperature in the spawning and feeding ground of Ommastrephes bartrami and its relationship with abundance index in the Northwestern Pacific Ocean[J]. Journal of Shanghai Fisheries University, 2005, 14(2): 168-175. |
| [17] |
徐兆礼, 陈佳杰. 依据大规模捕捞统计资料分析东黄渤海白姑鱼种群划分和洄游路线[J]. 生态学报, 2010, 30(23): 6442-6450. XU Z L, CHEN J J. Analysis to population division and migratory routine of populations and migratory routines of Argyrosomus argentatus in the north China waters[J]. Acta Ecologica Sinica, 2010, 30(23): 6442-6450. |
| [18] |
徐兆礼, 陈佳杰. 东黄海大黄鱼洄游路线的研究[J]. 水产学报, 2011, 35(3): 429-437. XU Z L, CHEN J J. Analysis of migratory route of Larimichthys crocea in the East China Sea and Yellow Sea[J]. Journal of Fisheries of China, 2011, 35(3): 429-437. |
| [19] |
陈佳杰, 徐兆礼, 陈雪忠. 我国沿海小黄鱼渔场的空间格局[J]. 水产学报, 2010, 34(2): 236-244. CHEN J J, XU Z L, CHEN X Z. The spatial distribution pattern of fishing ground for small yellow croaker in China Seas[J]. Journal of Fisheries of China, 2010, 34(2): 236-244. |
| [20] |
牛明香, 李显森, 徐玉成. 智利外海竹筴鱼中心渔场时空变动的初步研究[J]. 海洋科学, 2009, 33(11): 105-109. NIU M X, LI X S, XU Y C. Preliminary study on spatio-temporal change of central fishing ground of chilean jack mackerel (Trachurus murphyi) in the offshore waters of Chile[J]. Marine sciences, 2009, 33(11): 105-109. |
| [21] |
陈新军, 钱卫国, 许柳雄, 等. 北太平洋150°E-165°E海域柔鱼重心渔场的年间变动[J]. 湛江海洋大学学报, 2003, 23(3): 26-32. CHEN X J, QIAN W G, XU L X, et al. Comparison among annual positions of fishing grounds for Ommastrephe bartrami from 150°E to 165°E in the North Pacific[J]. Journal of Zhanjiang Ocean University, 2003, 23(3): 26-32. |
| [22] |
汪金涛, 陈新军. 中西太平洋鲣鱼渔场的重心变化及其预测模型建立[J]. 中国海洋大学学报, 2013, 43(8): 44-48. WANG J T, CHEN X J. Changes and prediction of the fishing ground gravity of skipjack (Katsuwonus pelamis) in western-central Pacific[J]. Periodical of Ocean University of China, 2013, 43(8): 44-48. |
| [23] |
汪振华, 赵静, 王凯, 等. 马鞍列岛岩礁生境鱼类群落结构时空格局[J]. 生态学报, 2013, 33(19): 6218-6226. WANG Z H, ZHAO J, WANG K, et al. Fish community ecology in rocky reef habitat of Ma'an Archipelago Ⅱ. Spatio-temporal patterns of community structure[J]. Acta Ecologica Sinica, 2013, 33(19): 6218-6226. |
| [24] |
蔡建堤, 陈方平, 吴建绍, 等. 渔场重心信度测算及渔场重心修正理论构建——以闽南-台湾浅滩渔场二长棘鲷为例[J]. 生态学报, 2015, 35(6): 1929-1937. CAI J T, CHEN F P, WU J S, et al. The theoretical construction of measuring fishing ground gravity reliability and correcting fishing ground gravity:a case study on Paerargyrops edita Tanaka in Minnan-Taiwan Bank Fishing Ground[J]. Acta Ecologica Sinica, 2015, 35(6): 1929-1937. |
| [25] |
汪金涛, 高峰, 雷林, 等. 基于主成分和BP神经网络的智利竹筴鱼渔场预报模型研究[J]. 海洋学报(中文版), 2014, 36(8): 65-71. WANG J T, GAO F, LEI L, et al. Application of BP neural network based on principal component analysis in fishing grounds of Chilean jack mackerel (Trachurus murphyi) in the southeast Pacific Ocean[J]. Acta Oceanologica Sinica, 2014, 36(8): 65-71. |
| [26] |
BENEDIKTSSON J A, SWAIN P H, ERSOY O K. Neural network approaches versus statistical methods in classification of multisource remote sensing data[J]. IEEE Transactions on Geoscience and Remote Sensing, 1990, 28(4): 540-552. DOI:10.1109/TGRS.1990.572944 |
| [27] |
HUSH D R, HORN0 E B G. Progress in supervised neural networks[J]. IEEE Signal Processing Magazine, 2002, 1(1): 8-39. |
| [28] |
CHEN X J, CAO J, CHEN Y, et al. Effect of the kuroshio on the spatial distribution of the red flying squid Ommastrephes bartramii in the Northwest Pacific Ocean[J]. Bulletin of Marine Science, 2012, 88(1): 63-71. DOI:10.5343/bms.2010.1098 |
| [29] |
YATSU A, CHIBA S, YAMANAKA Y, et al. Climate forcing and the Kuroshio/Oyashio ecosystem[J]. ICES Journal of Marine Science, 2013, 70(5): 922-933. DOI:10.1093/icesjms/fst084 |
| [30] |
FAN W, WU Y M, CUI X S. The study on fishing ground of neon flying squid, Ommastrephes bartrami, and ocean environment based on remote sensing data in the Northwest Pacific Ocean[J]. Chinese Journal of Oceanology and Limnology, 2009, 27: 408. DOI:10.1007/s00343-009-9107-1 |
| [31] |
VIJAI D, SAKAI M, KAMEI Y, et al. Spawning pattern of the neon flying squid Ommastrephes bartramii (Cephalopoda:Oegopsida) around the Hawaiian Islands[J]. Scientia Marina, 2014, 78(4): 511-519. DOI:10.3989/scimar.2014.78n4 |
2. The Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources, Shanghai Ocean University, Ministry of Education, Shanghai 201306, China;
3. National Distant-water Fisheries Engineering Research Center, Shanghai Ocean University, Shanghai 201306, China;
4. Key Laboratory of Oceanic Fisheries Exporation, Ministry of Agriculture, Shanghai 201306, China