 2018,
Vol. 27
2018,
Vol. 27



 (Acheilognathus chankaensis)、大鳍(Acheilognathus macropterus)为主要种类、异速生长,其他鱼类等速生长。聚类分析显示:太湖鳑鲏亚科鱼类组成有显著时空差异,时间为2组,5—12月为一组,N%、W%和IRI指数显示兴凯占优势,其余时间大鳍占优势;空间为两组,贡湖、漫山为一组,大鳍占优势,其余站点兴凯占优势。分析显示多样性指数均低于一般水平(1.5~3.5),其鱼类群落结构的时空差异与湖泊营养盐、环境因子、浮游生物、水生植物等分布格局密切相关。
(Acheilognathus chankaensis)、大鳍(Acheilognathus macropterus)为主要种类、异速生长,其他鱼类等速生长。聚类分析显示:太湖鳑鲏亚科鱼类组成有显著时空差异,时间为2组,5—12月为一组,N%、W%和IRI指数显示兴凯占优势,其余时间大鳍占优势;空间为两组,贡湖、漫山为一组,大鳍占优势,其余站点兴凯占优势。分析显示多样性指数均低于一般水平(1.5~3.5),其鱼类群落结构的时空差异与湖泊营养盐、环境因子、浮游生物、水生植物等分布格局密切相关。太湖是长江流域典型的大型浅水湖泊,水域面积2 338 km2,平均水深1.89 m,是我国第三大淡水湖,其地处亚热带,气候温和,河网密布,渔业资源十分丰富,历史上曾报道有鱼类103种[1-3]。有关太湖鱼类资源的调查始于20世纪50年代,内容主要集中于鱼类生物学特征、区系组成及食性等方面[4-6],倪勇等[7]在汇总前人研究的基础上,对太湖鱼类区系进行了系统调研,共计记录鱼类107种,分别隶属于14目25科73属。鱼类作为湖泊水生生物的重要类群,其群落结构与湖泊生态系统的稳定性息息相关[8],科学掌握湖泊鱼类群落结构现状并辨明其变动趋势,对于湖泊生态系统研究具有重要意义。
鳑鲏亚科(Acheilognathinae)隶属于鲤形目(Cypriniformes)鲤科(Cyprinidae),为典型小型鱼类,一般体长小于80 mm,最大不超过180 mm,大多栖息于江河、湖泊、水库等浅静水区,营杂食性生活,以浮游生物、碎屑等为食[9]。由于独特的形态、繁殖策略及细胞核学的特征,其在鲤科鱼类中显得极为特别,吸引了研究者极大的兴趣。目前对鳑鲏亚科鱼类的研究主要集中于繁殖习性、形态特征及分子系统学等几个方面[10-12],对其群落结构的研究却鲜有介绍。鳑鲏亚科鱼类是太湖鱼类的重要组成部分,对维护太湖生态系统的平衡起重要作用[6-7]。然而近几十年来,太湖水域生态环境不断恶化,江湖阻隔、围网养殖、过度捕捞及富营养化不断加剧、水草植被的大幅度减少等生境变化导致太湖鱼类资源量和区系组成发生了显著变化,鱼类小型化趋势十分明显[4, 6],进而鳑鲏亚科鱼类的种群特征及其在太湖鱼类群落中的生态地位也将发生相应的变化。本研究于2013年3月至2014年2月逐月对太湖鳑鲏亚科鱼类进行调查,了解其物种组成、数量分布及多样性特征等,以期掌握太湖鳑鲏亚科鱼类群落特征的时空变化规律等,为太湖鱼类生态学研究积累素材。
1 材料与方法 1.1 样点设置太湖位于长江下游,处于北纬30°55′40″~31°32′58″和东经119°52′32″~120°36′10″。依据太湖水域的实际情况,按照《内陆水域渔业自然资源调查手册》样点设置原则[13],共设置8个监测点,分别为大浦、贡湖、横山、鲤山湾、漫山、平台山、汤溇及竺山湖(图 1)。

|
图 1 太湖鱼类调查采样点分布
Fig. 1 The fish sampling stations in Lake Taihu
|
本次调查于2013年3月至2014年2月使用多网目复合刺网、地笼2种渔具对太湖8个样点进行逐月调查。其中:多网目复合刺网长125 m(每条刺网包含7种网目,网目1.2 cm、2 cm、4 cm各15 m;网目6 cm、8 cm、10 cm、14 cm各20 m),高1.5 m;地笼长、宽、高分别为25 m、0.3 m、0.3 m,网目0.8 cm,每次各样点分别放置3条刺网及3条地笼,12 h后收集所有渔获物,带回实验室鉴定到种[7, 14],同时按照Hubbs & Lagler描述的方法,使用电子数显卡尺和电子天平测定每一尾鱼的全长(0.1 cm)、体长(0.1 cm)及体质量(0.1 g)等生物学指标[15]。
1.3 数据处理 1.3.1使用PINKAS相对重要性指数(IRI)描述各鱼类重要性[16-17],公式如下:
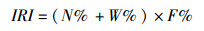
式中:N%为某一鱼类的尾数占总尾数的百分比,W%为某一鱼类的质量占总质量的百分比,F%为某一鱼类出现的次数占总调查次数的百分比,其中IRI>1 000,为群落优势种;IRI>100,为群落主要种;IRI<10,为群落少见种或偶见种。
1.3.2Margalef物种丰富度指数(D):
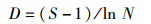
Shannon-Wiener多样性指数(H′):
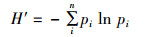
Evenness均匀度指数(E):
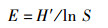
Simpson优势度指数(D′):
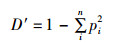
式中:S为鳑鲏亚科鱼类种类数;N为鳑鲏亚科鱼类总尾数;pi为第i种鱼所占的数量百分比,群落多样性指数差异采用单因素方差分析,多重比较采用Duncan氏检验。
采用组平均聚类法对鳑鲏亚科鱼类进行时间、空间聚类分析,探讨太湖鳑鲏亚科鱼类群落组成的时空变化,以d=1-r(r为Pearson相关系数)作为相异性指标,将d=0.3作为存在显著差异的依据[20]。
1.3.3使用幂函数描述鱼类体长与体质量的相互关系[21],公式如下:
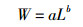
式中:W表示鱼类体质量(g),L表示鱼类全长(cm),a、b为相关参数,其中a为生长的条件因子,b值判断鱼类的生长情况,表示鱼类生长发育的不均匀性,当鱼类b值与b=3时无显著差异时,表示鱼类的体长和体质量关系符合欧氏几何中的体积随长度的变化关系,即等速生长;当b值与b=3呈显著差异时,表示鱼类体长和体质量关系不符合欧氏几何中的体积随长度的变化关系,即非等速生长。
2 结果 2.1 种类组成共采集鱼类52种10 358尾,隶属于8目23科。数量百分比(N%)显示鳑鲏亚科(24.44%)、亚科(18.33%)、鳀科(14.37%)占优势,其中刀鲚(14.37%)、兴凯

|
图 2 太湖渔获物数量、质量及相对重要性指数百分比
Fig. 2 The number, weight and index of relative importance of catch in Lake Taihu
|
共采集到2属7种2 531尾鳑鲏亚科鱼类,占总数的24.44%,生物量仅占1.63%。其中兴凯

|
表 1 太湖鳑鲏亚科鱼类组成 Tab.1 The composition of Acheilognathinae fish in Lake Taihu |
分析显示太湖鳑鲏亚科鱼类数量及生物量呈相似波浪形时间变化,4月、10月及1月较高,6月、7月及11月较低(图 3)。对鱼类数量进行聚类显示,其组成有显著时间差异,5月至12月为一类,兴凯

|
图 3 太湖鳑鲏亚科鱼类组成的月度变化
Fig. 3 The monthly variations of Acheilognathinae in Lake Taihu
|

|
图 4 太湖鳑鲏亚科鱼类组成时间变化的聚类分析
Fig. 4 Dendrogram of the time variation of Acheilognathinae in Lake Taihu
|

|
表 2 太湖鳑鲏亚科鱼类数量及质量百分比的时间变化 Tab.2 The time variation of the percentage by number and weight of Acheilognathinae in Lake Taihu |
分析显示太湖鳑鲏亚科鱼类数量及生物量呈相似波浪形空间变化,北部贡湖、竺山湖等较高,东部横山、鲤山湾等较低(图 5)。对鱼类数量进行聚类显示,其组成有显著空间差异,漫山、贡湖为一类,大鳍

|
图 5 太湖鳑鲏亚科鱼类组成的空间变化
Fig. 5 The spatial variations of the Acheilognathinae in Lake Taihu
|

|
图 6 太湖鳑鲏亚科鱼类组成空间变化聚类分析
Fig. 6 Dendrogram of the spatial variation of Acheilognathinae in Lake Taihu
|

|
表 3 太湖鳑鲏亚科鱼类数量及质量百分比的空间变化 Tab.3 The spatial variation of the percentage by number and weight of Acheilognathinae in Lake Taihu |
太湖鳑鲏亚科鱼类丰富度(R)、多样性(H′)、均匀度(E)、优势度(D′)等指数没有显著的时间差异(R:X2=19.04,P=0.06;H′: X2=16.82,P=0.11;E:X2=16.67,P=0.12;D′:X2=19.04,P=0.06)。前三种指数呈现相似的月度间变化,均与优势度指数变化相反,6月多样性指数较高(图 7)。太湖鳑鲏亚科鱼类指数R、H′没有显著的空间差异(R:X2=11.32,P=0.13;H′: X2=12.38,P=0.89),而指数E、D′呈现显著空间差异(E:X2=14.98,P=0.036;D′:X2=21.75,P=0.003),Duncan氏检验显示横山、鲤山湾、贡湖、漫山没有差异,均与平台山、竺山湖处有差异(图 8)。

|
图 7 太湖鳑鲏亚科4种多样性指数的时间变化
Fig. 7 The time variation of four kinds of diversity index of Acheilognathinae in Lake Taihu
|

|
图 8 太湖鳑鲏亚科4种多样性指数的空间变化
Fig. 8 The spatial variation of four kinds of diversity index of Acheilognathinae in Lake Taihu
|
相关性显示多样性指数与丰富度(H′=0.985 9 R+0.032 1,R2=0.824 9,P<0.001)、均匀度(H′=0.947 5 E+0.009 3,R2=0.854 1,P<0.001)、物种数(H′=0.219 8S-0.0281,R2=0.803 7,P<0.001)等均呈显著正相关性,与优势度(H′=-1.4859 D′+1.5178,R2=0.795 2,P<0.001)呈显著负相关性,物种多样性指数依靠各指数变化而变化,与均匀度指数相关性最显著。
2.4 体长、体质量特征 2.4.1 体长组成如图 9为各鳑鲏亚科鱼类全长分布,结果显示全长在1.9~12.9 cm之间,平均7.5 cm,其中6~8 cm为鳑鲏亚科鱼类全长优势组。大鳍

|
图 9 太湖鳑鲏亚科鱼类全长组成
Fig. 9 The composition of the total length of Acheilognathinae in Lake Taihu
|
如图 10为各鳑鲏亚科鱼类体质量分布,结果显示体质量在0.3~31.9 g之间,平均5.9 g,其中30%的鱼类体质量在2~4 g;51.5%的鱼类体质量小于5 g,86.7%的鱼类体质量小于10 g。大鳍

|
图 10 太湖鳑鲏亚科鱼类体质量组成
Fig. 10 The composition of the weight of Acheilognathinae in Lake Taihu
|
使用幂指数公式(W=aLb)描述鱼类体长和体质量相互关系,结果显示,相关系数R2在0.724 1~0.910 2之间,均值为0.822 1,相关性高,幂指数b值为2.703 5~3.139 1,均值为2.901 6,与b=3没有显著性差异(P=0.26)。统计每种鱼类的b值与b=3的差异性显示,大鳍

|
表 4 太湖鳑鲏亚科鱼类体长和体质量相互关系 Tab.4 The relationship between length and weight of Acheilognathinae in Lake Taihu |
资料显示,20世纪60~80年代,太湖鱼类记录有82~106种[5, 7],而21世纪以来,其发生了较大变化。一是种类数明显减少,本世纪的几次调查显示鱼类有50~60种[4-6],减少了将近50.0%,本次调查亦如此,仅采集到鱼类50种,其中洄游性鱼类(鲟形目、鲻形目等)、溪流性鱼类(马口鱼、宽鳍
据记录太湖鳑鲏亚科鱼类共2属11种(中国3属30种[14]),其中兴凯
太湖鳑鲏亚科鱼类组成有明显的时空差异。其中5月至12月主要为兴凯
高强度的捕捞、环境污染及人类活动等都能导致鱼类多样性指数的降低[32]。太湖鳑鲏亚科鱼类多样性指数在各时间段(0.576 4 ~1.428 2)和各湖区(0.668 2 ~1.124 2)尺度上均表现偏低,低于Magurran提出的鱼类多样性指数的一般范围(1.5~3.5),表明太湖鳑鲏亚科鱼类资源衰退及优势种单一化等。有结果显示,鳑鲏类、虾类等以浮游动物为食物的小型水生生物类群,可能鲤、鲫数量的增加对其有促进作用,而与鲚鱼间有食物上的相互竞争[33]。本次调查显示,鳑鲏亚科鱼类数量与鲤亚科鱼类(P=0.042)、鲢亚科鱼类(P=0.001)均呈显著正相关性,这种浮游生物食性鱼类产量随着鲤、鲫产增加而增加的现象和一些研究结论接近[34]。对鳑鲏亚科鱼类间进行相关性分析显示,大鳍
| [1] |
秦伯强, 胡维平, 陈伟民, 等. 太湖水环境演化过程与机理[M]. 北京: 科学出版社, 2004.
QIN B Q, HU W P, CHEN W M, et al. Process and mechanism of environmental changes of the Taihu Lake[M]. Beijing: Science Press, 2004. |
| [2] |
朱松泉. 2002-2003年太湖鱼类学调查[J].
湖泊科学, 2004, 16(2): 120–124.
ZHU S Q. Ichthyological survey of Lake Taihu during 2002-2003[J]. Journal of Lake Sciences, 2004, 16(2): 120–124. |
| [3] |
谷孝鸿, 白秀玲, 江南, 等. 太湖渔业发展及区域设置与功能定位[J].
生态学报, 2006, 26(7): 2247–2254.
GU X H, BAI X L, JIANG N, et al. Fishery development, regional classification and functional positioning of Lake Taihu[J]. ActaEcologicaSinica, 2006, 26(7): 2247–2254. |
| [4] |
刘恩生, 刘正文, 陈伟民, 等. 太湖鱼类产量、组成的变动规律及与环境的关系[J].
湖泊科学, 2005, 17(3): 251–255.
LIU E S, LIU Z W, CHEN W M, et al. Changes in the yield and composition of the fish catches and their relation to the environmental factors in Lake Taihu[J]. Journal of Lake Sciences, 2005, 17(3): 251–255. |
| [5] |
朱松泉, 刘正文, 谷孝鸿. 太湖鱼类区系变化和渔获物分析[J].
湖泊科学, 2007, 19(6): 664–669.
DOI:10.18307/2007.0607 ZHU S Q, LIU Z W, GU X H. Changes of the fish fauna and fish yield analysis in Lake Taihu[J]. Journal of Lake Sciences, 2007, 19(6): 664–669. DOI:10.18307/2007.0607 |
| [6] |
毛志刚, 谷孝鸿, 曾庆飞, 等. 太湖鱼类群落结构及多样性[J].
生态学杂志, 2011, 30(12): 2836–2842.
MAO Z G, GU X H, ZENG Q F, et al. Community structure and diversity of fish in Lake Taihu[J]. Chinese Journal of Ecology, 2011, 30(12): 2836–2842. |
| [7] |
倪勇, 朱成德. 太湖鱼类志[M]. 上海: 上海科学技术出版社, 2005.
NI Y, ZHU C D. Fishes of the Taihu Lake[M]. Shanghai: Shanghai Scientific & Technical Publishers, 2005. |
| [8] | SCHEFFER M, CARPENTER S, FOLEY J A, et al. Catastrophic shifts in ecosystems[J]. Nature, 2001, 413(6856): 591–596. DOI:10.1038/35098000 |
| [9] |
陈欣. 我国鳑鲏亚科鱼类遗传多样性及系统发育研究[D]. 新乡: 河南师范大学, 2011.
CHEN X. The study of genetic diversity and molecular phylogenetic of Acheilognathinae[D]. Xinxiang: Henan Normal University, 2011. http://cdmd.cnki.com.cn/Article/CDMD-10476-1012281635.htm |
| [10] |
陈校辉, 倪勇, 伍汉霖. 江苏省鳑鲏属(Rhodeus)鱼类的研究[J].
海洋渔业, 2005, 27(2): 89–97.
CHEN X H, NI Y, WU H L. Fishes of genus Rhodeus Agassiz in Jiangsu Province, China[J]. Marine Fisheries, 2005, 27(2): 89–97. |
| [11] |
朱玉蓉, 刘焕章. 大鳍基于细胞色素b基因序列的遗传变异及生物地理过程[J].
水生生物学报, 2006, 30(2): 134–140.
ZHU Y R, LIU H Z. Genetic diversity and biogeographical process of Acheilognathus macropterus revealed by sequence variations of mitochondrial cytochrome bgene[J]. Acta Hydrobiologica Sinica, 2006, 30(2): 134–140. |
| [12] | BOHLEN J, ÁLECHTOVÁ V, BOGUTSKAYA N, et al. Across Siberia and over Europe:phylogenetic relationships of the freshwater fish genus Rhodeus in Europe and the phylogenetic position of R. sericeus from the River Amur[J]. Molecular Phylogenetics and Evolution, 2006, 40(3): 856–865. DOI:10.1016/j.ympev.2006.04.020 |
| [13] |
张觉民, 何志辉. 内陆水域渔业自然资源调查手册[M]. 北京: 农业出版社, 1991.
ZHANG J M, HE Z H. A manual for investigation of natural resources in inland waters[M]. Beijing: Agriculture Press, 1991. |
| [14] |
倪勇, 伍汉霖. 江苏鱼类志[M]. 北京: 中国农业出版社, 2006.
NI Y, WU H L. Fishes of Jiangsu province[M]. Beijing: China Agriculture Press, 2006. |
| [15] | HUBBS C L, LAGLER K F. Fishes of the Great Lakes region[M]. Michigan: University of Michigan Regional, 2004. |
| [16] | PINKAS L, OLIPHANT S M, IVERSON I L K. Food habits of albacore, bluefin tuna, and bonito in California waters[R]. State of California: California Department of Fish and Game, Fish Bulletin. 1971: 1-105. |
| [17] |
程济生, 俞连福. 黄、东海冬季底层鱼类群落结构及多样性变化[J].
水产学报, 2004, 28(1): 29–34.
CHENG J S, YU L F. The change of structure and diversity of demersal fish communities in the Yellow Sea and East China Sea in winter[J]. Journal of Fisheries of China, 2004, 28(1): 29–34. |
| [18] | LUDWIG J A, REYNOLDS J F. Statistical ecology, a primer on methods and computing[J]. Journal of Wildlife Management, 1990, 54(1): 197–198. DOI:10.2307/3808926 |
| [19] | KREBS C J. Ecological methodology[M]. New York: Harper Collins Publishers, 1989. |
| [20] | HU Z J, WANGS Q, WU H, et al. Temporal and spatial variation of fish assemblages in Dianshan Lake, Shanghai, China[J]. Chinese Journal of Oceanology and Limnology, 2014, 32(4): 799–809. DOI:10.1007/s00343-014-3193-4 |
| [21] | RICKER W E. Linear regressions in fishery research[J]. Journal of the Fisheries Research Board of Canada, 1973, 30(3): 409–434. DOI:10.1139/f73-072 |
| [22] |
孙明波, 谷孝鸿, 曾庆飞, 等. 基于水声学方法的太湖鱼类空间分布和资源量评估[J].
湖泊科学, 2013, 25(1): 99–107.
DOI:10.18307/2013.0113 SUN M B, GU X H, ZENG Q F, et al. Assessment of fish spatial distribution and biomass in Lake Taihu using hydroacoustic method[J]. Journal of Lake Sciences, 2013, 25(1): 99–107. DOI:10.18307/2013.0113 |
| [23] |
张蓉, 杨龙元, 谷孝鸿, 等. 东太湖典型湖区水环境特征时空变化分析[J].
水利学报, 2007, 38(S1): 597–605.
ZHANG R, YANG L Y, GU X H, et al. Study on the water environmental characters for spatial and temporal variation in the typical districts of Lake East Tai[J]. Journal of Hydraulic Engineering, 2007, 38(S1): 597–605. |
| [24] |
史赟荣, 晁敏, 沈新强. 长江口张网鱼类群落结构特征及月相变化[J].
海洋学报, 2014, 36(2): 81–92.
SHI Y R, CHAO M, SHEN X Q. Characteristics and monthly variations of set net fish community structure in the Changjiang River estuary[J]. Acta Oceanologica Sinica, 2014, 36(2): 81–92. |
| [25] | QIN B Q, XU P Z, WU Q L, et al. Environmental Issues of Lake Taihu, China[J]. Hydrobiologia, 2007, 581(1): 3–14. DOI:10.1007/s10750-006-0521-5 |
| [26] |
冯露露, 李正魁, 周涛. 太湖浮游植物和各形态无机氮的时空分布特征[J].
湖泊科学, 2012, 24(5): 739–745.
DOI:10.18307/2012.0515 FENG L L, LI Z K, ZHOU T. Temporal and spatial distributions of phytoplankton and various forms of inorganic nitrogen in Lake Taihu[J]. Journal of Lake Sciences, 2012, 24(5): 739–745. DOI:10.18307/2012.0515 |
| [27] |
何俊, 谷孝鸿, 刘国锋. 东太湖水生植物及其与环境的相互作用[J].
湖泊科学, 2008, 20(6): 790–795.
DOI:10.18307/2008.0618 HE J, GU X H, LIU G F. Aquatic macrophytes in East Lake Taihu and its interaction with water environment[J]. Journal of Lake Sciences, 2008, 20(6): 790–795. DOI:10.18307/2008.0618 |
| [28] |
谷孝鸿, 张圣照, 白秀玲, 等. 东太湖水生植物群落结构的演变及其沼泽化[J].
生态学报, 2005, 25(7): 1541–1548.
GU X H, ZHANG S Z, BAI X L, et al. Evolution of community structure of aquatic macrophytes in Lake East Taihu and its wetlands[J]. Acta Ecologica Sinica, 2005, 25(7): 1541–1548. |
| [29] |
朱广伟. 太湖富营养化现状及原因分析[J].
湖泊科学, 2008, 20(1): 21–26.
DOI:10.18307/2008.0103 ZHU G W. Eutrophic status and causing factors for a large, shallow and subtropical Lake Taihu, China[J]. Journal of Lake Sciences, 2008, 20(1): 21–26. DOI:10.18307/2008.0103 |
| [30] |
赵凯, 李振国, 魏宏农, 等. 太湖贡湖湾水生植被分布现状(2012年)[J].
湖泊科学, 2015, 27(3): 421–428.
DOI:10.18307/2015.0308 ZHAO K, LI Z G, WEI H N, et al. The distribution of aquatic vegetation in Gonghu Bay, Lake Taihu, 2012[J]. Journal of Lake Sciences, 2015, 27(3): 421–428. DOI:10.18307/2015.0308 |
| [31] | KE Z X, XIE P, GUO L G, et al. In situ study on the control of toxic Microcystis blooms using phytoplanktivorous fish in the subtropical Lake Taihu of China:a large fish pen experiment[J]. Aquaculture, 2007, 265(1/4): 127–138. |
| [32] | WASHINGTON H G. Diversity, biotic and similarity indices:a review with special relevance to aquatic ecosystems[J]. Water Research, 1984, 18(6): 653–694. DOI:10.1016/0043-1354(84)90164-7 |
| [33] |
刘恩生, 刘正文, 鲍传和, 等. 太湖鲚鱼和鲢、鳙鱼的食物组成及相互影响分析[J].
湖泊科学, 2007, 19(4): 451–456.
DOI:10.18307/2007.0414 LIU E S, LIU Z W, BAO C H, et al. Food content and the mutual effects between Hypophthalmich thysmolitrix (Cuvier etValenciennes) and Aristichthys nobilos (Richardson) and Coilia ectenes taihuensis Yen et Lin in Lake Taihu[J]. Journal of Lake Sciences, 2007, 19(4): 451–456. DOI:10.18307/2007.0414 |
| [34] | RICHARDSON W B, WICKHAM S A, THRELKELD S T. Foodweb response to the experimental manipulations of a benthivore (Cypinus carpio), zooplanktivore (Menidia beryllina) and benthic insects[J]. ArchivFür Hydrobiologie, 1990, 119(2): 143–165. |