 2016,
Vol. 25
2016,
Vol. 25



2. 上海市浦东新区农业委员会执法大队渔政中队, 上海 201308
杭州湾与长江入海口南侧相毗邻,杭州湾北岸水域主要受潮流、东南向强浪以及长江口径流影响[1]。闫欣等[2]研究表明该水域是多种海洋和河口性早期发育阶段鱼类的生活栖息地。同时,作为上海经济发展战略的重点地区,杭州湾北岸水域环境正遭受严重污染[3]。浮游动物是中上层鱼类和仔稚鱼的良好饵料[4],其群落结构变化会不同程度地影响渔业生物结构的变动[5]。在河口和近岸水域,受环境参数和人类活动影响,浮游动物群落结构和优势种发生相应的变化和演替[6-7]。
国内外关于河口近岸水域浮游动物的报道有很多,杭州湾和长江口附近水域浮游动物的报道已屡见不鲜[8-12]。杭州湾附近水域浮游动物种类、生态类型和分布特征已得到一定的了解,但是对于表层浮游动物情况未见报道。面对全球气候变暖,我国近海海表水温明显上升[13],表层水域浮游动物情况未曾可知。本文旨在从表层水域浮游动物研究出发,在全球气候变暖的大前提下,通过对该水域浮游动物种类组成、生物量和丰度、多样性以及与相关环境因子的关系等分析,为相关河口水域系统评价提供可靠依据。同时,初步探讨该水域表层浮游动物与仔稚鱼的相关性。
1 材料与方法 1.1 采样时间、地点及方法2014年8月(夏季)、11月(秋季)和2015年2月(冬季)、5月(春季),在杭州湾北岸水域(121°45′~122°09′E,30°45′~30°51′N)设置10个站点(相邻站点距离约为5 nmiles,水深6~10 m)见图 1。采用大型仔稚鱼网(口径1.3 m,网目0.5 mm)进行表层水平拖网调查,渔船(沪捕鱼No.49627,120马力,30 t)拖速为每小时2 nmiles,每个站点拖网10 mins,并用网口流量计(KC Denmark A/S Model 23.090)记录拖网流量值。所采样本现场用5%的甲醛固定,并测定水温、盐度。同时,按照《海洋监测规范》(GB 17378.4-2007)和(GB 17378.7-2007)现场采水样,用于室内测定溶氧和叶绿素a。

|
图 1 采样站点
Fig. 1 Sampling stations
|
将采集的样本进行垃圾清理,用湿重法测定每个样本的生物量。利用普通显微镜(OLYMPUS CX21)和体式显微镜(Nikon SMZ800),按照《中国海洋浮游桡足类图谱》、《中国海洋浮游生物图谱》、《中国动物志淡水桡足类》等参考书进行种类鉴定,并做好相关记录。
1.3 数据分析浮游动物丰度的计算公式:
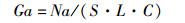 (1)
(1)
式中:Ga为单位体积内浮游动物的个体数(ind./m3);Na为每网浮游动物总个体数;S为网口面积(m3);L为流量计转数;C为流量计校正值(C=0.3)。
浮游动物优势度(Y)的计算公式:
 (2)
(2)
式中:ni是第i种的个体数,N是所有种类的个体数,fi为出现频率。Y≥0.02的种类为优势种。
优势种更替率(R)的计算公式:
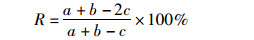 (3)
(3)
式中:a、b为相邻两季节优势种种数,c为相邻两季节共同拥有优势种种数。
Shannon-Wiener多样性指数(H′):
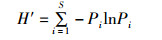 (4)
(4)
Pielou均匀度指数(J′):
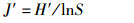 (5)
(5)
Margalef丰富度指数(D):
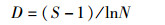 (6)
(6)
式中:H′为物种多样性指数,J′为物种均匀度指数,D为物种丰富度指数,Pi为第i种的个体数与样品中总个体数的比值,S代表该站浮游动物的种类数,N为样品中所有种类的个体数。
2 结果与分析 2.1 种类组成杭州湾北岸表层水域4季共鉴定浮游动物12类42种(表 1),浮游幼体5种(不计)。其中桡足类最多,共17种,其次为水母类,共6种。其他如端足类3种,糠虾类2种,涟虫类1种,等足类1种,翼足类1种,毛颚类2种,多毛类2种,十足类4种,磷虾类1种,樱虾类2种。春季鉴定浮游动物种类最多,共29种,冬季最少,只有16种。

|
表 1 杭州湾北岸表层水域浮游动物种类组成 Tab.1 Species composition of surface zooplankton in the north shore of Hangzhou Bay |
该水域浮游动物按照其对盐度适应性主要可分为4种生物类群[14-15]:淡水类群、河口类群、近岸低盐类群和广盐类群。淡水类群,有汤匙华哲水蚤(Neodiaptomus schmackeri)、近邻剑水蚤(Cyclops vicinus)等。河口类群,有中华华哲水蚤(Sinocalanus sinensis)、虫肢歪水蚤(Tortanus vermiculus)、火腿伪镖水蚤(Pseudodiaptomus poplesia)等。近岸低盐类群为主要类群,有真刺唇角水蚤(Labidocera euchaeta)、背针胸刺水蚤(Centropages dorsispinatus)、太平洋纺锤水蚤(Acartia pacifica)、克氏纺锤水蚤(Acartia clausi)、长额刺糠虾(Acanthomysis longirostris)、中华假磷虾(Pseudeuphausia sinica)、拿卡箭虫(Sagitta nagae)等。广盐类群,有近缘大眼水蚤(Corycaeus affinis)、中华哲水蚤(Calanus sinicus)、小拟哲水蚤(Paracalanus parvus)等。
2.2 优势种杭州湾北岸表层水域4季共出现优势种10种。分别是克氏纺锤水蚤、太平洋纺锤水蚤、背针胸刺水蚤、中华胸刺水蚤(Centropages sinensis)、真刺唇角水蚤、火腿伪镖水蚤、虫肢歪水蚤、针刺拟哲水蚤(Paracalanus aculeatus)、小拟哲水蚤(Paracalanus parvus)和中华华哲水蚤。四季均出现的优势种为真刺唇角水蚤、火腿伪镖水蚤、虫肢歪水蚤3种。
其优势种组成具有明显的季节变化(表 2)。秋季优势种最多(7种),夏季最少(4种)。夏季,太平洋纺锤水蚤和火腿伪镖水蚤平均丰度值达到508.56 ind./m3和493.40 ind./m3,占当季总丰度的39.41%和38.23%。秋季,中华胸刺水蚤和真刺唇角水蚤的平均丰度值达到19.77 ind./m3和22.01 ind./m3,占当季总丰度的26.45%和29.46%。冬季,虫肢歪水蚤和克氏纺锤水蚤的平均丰度值达到31.94 ind./m3和16.79 ind./m3,占当季总丰度的47.66%和25.05%。春季,虫肢歪水蚤和针刺拟哲水蚤的平均丰度值达到339.52 ind./m3和144.56 ind./m3,占当季总丰度的59.39%和25.29%。结合优势种更替率R,夏-秋R值为43%,秋-冬R值为67%,冬-春R值为57%,春-夏R值为50%,可见,季节变化对优势种组成影响很大。

|
表 2 优势种的平均丰度值、优势度以及丰度百分比的季节变化 Tab.2 The dominance (Y), average abundance (x) and the percentage of abundance (N%) of the dominant species |
杭州湾北岸表层水域浮游动物生物量和丰度有明显的季节变化且二者显著相关(P < 0.01),生物量水平分布无明显规律(图 2)。夏季平均生物量达到最大,值为120.17 mg/m3,各站点幅度范围为38.73~307.23 mg/m3。其次为春季,平均生物量为59.56 mg/m3,各站点幅度范围为21.74~134.61 mg/m3。秋、冬生物量相近,平均值分别为14.51 mg/m3和14.21 mg/m3,各站点幅度范围分别为7.62~27.36 mg/m3和5.94~43.12 mg/m3。夏季平均丰度达到最大,值为1 290.53 ind./m3,各站点幅度范围为524.03~2 131.85 ind./m3。其次为春季,平均丰度为571.69 ind./m3,各站点幅度范围为106.40~1 924.81 ind./m3。秋、冬两季丰度相近,平均值分别为74.72 ind./m3和67.03 ind./m3,各站点幅度范围分别为37.84~120.47 ind./m3和29.81~196.54 ind./m3。

|
图 2 四季浮游动物生物量和丰度水平分布
Fig. 2 The horizontal distribution of biomass and abundance in four seasons
|
该水域多样性指数、均匀度指数和丰富度指数也出现明显的季节性变化(表 3)。其中夏季因太平洋纺锤水蚤和火腿伪镖水蚤为绝对优势种,且物种个体分布极不均匀导致3种指数最低。四季之中,秋季3种指数最高。春季种类数最高,但因个体分布不均导致多样性指数偏低。冬季种类数最少,但因个体分布较均匀,其多样性指数较高。

|
表 3 四季多样性指数、物种均匀度指数和物种丰富度指数 Tab.3 The indexes of Shannon-Wiener, Pielou and Margalef in four seasons |
杭州湾北岸表层水域温度和盐度季节性变化明显(表 4)。温度在夏季达到最大,平均表层水温为27.33 ℃,最低温在冬季出现,平均表层水温仅为6.37 ℃。该水域盐度主要受长江径流、潮流和风浪等影响而波动。夏季(丰水期)受长江冲淡水影响,盐度最低,平均表层盐度为4.6。随着降雨量的减少,盐度随之上升,冬季达到最大值,平均表层盐度为20。表层叶绿素a的含量幅度在1.23~4.28 μg/L之间,三季中(夏季数据空缺)冬季叶绿素a含量最高。表层溶氧具有随温度上升而下降的趋势,三季中(夏季数据空缺)冬季溶氧最高,平均表层溶氧值为11.77 mg/L。

|
表 4 相关环境因子季节变化 Tab.4 Seasonal variation of related environmental factors |
利用SPSS软件,对丰度、生物量、种类数和多样性指数分别与环境因子进行双变量相关性分析。结果表明(表 5)浮游动物各项指标皆与相应的环境因子有相关性。其中温度分别与丰度、生物量和多样性极显著相关,R值分别为0.706,0.632,和-0.440,表明温度与丰度和生物量间中度相关且成正相关,与多样性低度相关成负相关。盐度与丰度、生物量和多样性极显著相关,与种类数显著相关,R值分别为-0.678,-0.595,0.311和0.472,表明盐度与丰度和生物量间中度相关,且成负相关,与种类数和多样性低度相关,且成正相关。叶绿素a仅仅与种类数低度相关且成负相关(R值为-0.495)。溶解氧与生物量和种类数相关,与生物量成低度负相关(R值为-0.362),但与种类数成高度负相关(R值为-0.870)。

|
表 5 浮游动物各项指标与环境因子的相关性 Tab.5 Pearson correlations of environmental factors and parameters of zooplankton community |
杭州湾北岸表层水域浮游动物包括12大类,其中以桡足类为优势类群,占总种数的40.5%,这符合河口水域浮游动物分布的基本特征,且已被国内外许多河口证实[8-12, 16-17]。杭州湾水域属于钱塘江河口下游段,与开阔的海洋自由相通,主要受两股水流影响:一是由长江径流和钱塘江径流与海水混合形成的江浙沿岸水流;另一是由向北移动的黑潮暖流表层水与来自台湾海峡的海水混合形成的台湾暖流表层水流[18]。从本研究浮游动物生态类群可以看出,该水域浮游动物很好地反映了水文环境特征,其中包括淡水指示种汤匙华哲水蚤、近邻剑水蚤等,河口半咸水指示种火腿伪镖水蚤、虫肢歪水蚤等,近岸低盐水系指示种真刺唇角水蚤、太平洋纺锤水蚤等,暖水指示种针刺拟哲水蚤、精致真刺水蚤等。然而,近岸低盐和河口类群为该水域主要类群,说明杭州湾北岸表层水域主要受江浙沿岸水流影响。
3.2 浮游动物季节性变化与环境因子相关性温度和盐度是影响浮游动物个体生长、发育和繁殖的关键因素,温度和盐度的变化直接影响浮游动物群落结构变动[19]。河口水域水文物理结构复杂,水体的水平移动和垂直混合也会影响浮游动物分布[20]。随着环境因子、水体运动的季节性变化,杭州湾北岸表层水域浮游动物种类、生物量、丰度、多样性等也随之改变。结果表明温度和盐度是影响浮游动物分布的两个关键因子。由于温度的适宜,该水域浮游动物种类数在春季达到最大。夏季因丰水期的到来,长江冲淡水影响力增大,再加上浙江近岸上升流的盛行[21],可能导致河口种火腿伪镖水蚤、虫肢歪水蚤和近岸低盐种太平洋纺锤水蚤在表层大量聚集,使总生物量和丰度达到4个季节最大值,同时,因物种个体分布极不均匀导致夏季多样性指数最低。优势种季节更替率,进一步说明环境因子变动导致优势种组成变化。此外,河口浮游动物因为受到剧烈的水体运动,其水平分布并不像河口外海域中的斑块分布,而是具有多变性[22]。杭州湾北岸表层水域浮游动物生物量水平分布无明显规律,但夏季站点间生物量水平波动最大,春季次之,秋冬最小,可见不同季节水体运动程度对该水域浮游动物水平分布有所影响。
3.3 浮游动物与仔稚鱼相关性浮游动物中毛颚类和水母类被视为仔稚鱼的敌害生物[23],结合同月份仔稚鱼捕获均值(由课题组提供),将浮游动物平均丰度,敌害生物平均丰度与仔稚鱼平均捕获量进行对比分析(图 3)。浮游动物平均丰度夏季最高,春季次之,敌害生物春季平均丰度最大,夏秋相当,占浮游动物比例范围在0.06%~5.43%之间,且秋季所占比例较高,仔稚鱼在春季捕获量最高,其次为夏季。关于浮游动物与仔稚鱼关系研究,MONTELEONE[24]发现仔稚鱼丰度与浮游动物的数量有一定的关系。而AZEITEIRO[25]研究表明,仔稚鱼在不同发育阶段对饵料有所选择导致二者之间并无相关关系。本研究结果表明浮游动物仔稚鱼数量与浮游动物和敌害生物丰度无明显相关性(P>0.05),且浮游动物和仔稚鱼最大值出现在不同季节,但4个季节浮游动物丰度远大于仔稚鱼丰度(仔稚鱼最大丰度小于0.05 ind./m3)。另外多个研究表明[26-29],火腿伪镖水蚤、真刺唇角水蚤、虫肢歪水蚤、长额刺糠虾等桡足类和糠虾类为刀鲚、凤鲚、鳀鱼、小黄鱼等多种仔稚鱼和成鱼的主要饵料生物。可见,无论浮游动物的种类还是丰度,均能满足该水域出现的仔稚鱼和成鱼摄食。此外,敌害生物丰度小表明该水域能为仔稚鱼提供很好的庇护场所。

|
图 3 仔稚鱼与浮游动物和敌害生物关系
Fig. 3 The relationship between larval and juvenile fishes, zooplankton and harmful organisms
|
本研究为表层水平拖网采样,相关水域浮游动物研究多采用垂直拖网[5, 30-34]。与历史数据结果比较可知,两种采样方式所得浮游动物种类组成以及生物量和丰度4个季节变化趋势大体一致,具体差异主要体现在不同季节生物量和丰度值、优势种组成以及多样性指数三个方面。
徐兆礼等[5]对于2001年杭州湾洋山岛等调查中的1号站位点在本研究调查范围内。数据显示2月1日站点生物量为14 mg/m3,8月1日站点生物量为72.50 mg/m3。经对比,本研究2月表层生物量偏低,而8月表层平均生物量偏高。纪焕红等[30]于2002年8月对长江口浮游动物监测调查中指出,南汇嘴附近水域浮游动物丰度比长江口南支、长江口混合区以及长江口外海区都大,幅度范围在30~1 078.33 ind./m3,而本调查8月丰度幅度范围在524.03~2131.85 ind./m3。张冬融等[31]2009, 2011, 2012三年针对杭州湾不同水域秋季浮游动物调查表明,秋季北岸东侧水域优势种有5种,分别为真刺唇角水蚤、针刺拟哲水蚤、虫肢歪水蚤,火腿伪镖水蚤和中华胸刺水蚤。而本文调查水域秋季共有7种优势种,除上述5种外,太平洋纺锤水蚤和背针胸刺水蚤也为优势种。纪焕红等[30]于2002年长江口浮游动物监测调查中指出,火腿伪镖水蚤、虫肢歪水蚤和长额刺糠虾为8月优势种,而本调查8月优势种为太平洋纺锤水蚤、真刺唇角水蚤、火腿伪镖水蚤和虫肢歪水蚤4种。多个水域历史数据[32-34]表明,多样性指数在夏季达到最大值,而本研究夏季却出现最低值,且四季多样性指数皆相对偏低。
以上差异对比可推断该水域浮游动物具有垂直分层现象,一方面体现在优势种种类组成,有研究表明,不同季节由于温度影响,浮游动物会出现垂直移动现象,从而导致优势种分层现象[35];另一方面体现在生物量和丰度值,有研究表明[36-37],水体中的物理变化,导致沿岸水域出现浮游动物薄层(thin layer)现象,这种生物聚集现象垂直范围在数厘米到几米,水平范围数公里并可以持续数小时甚至更久。本研究夏季表层生物量偏大,可能是因为长江冲淡水和浙江近岸上升流共同影响导致浮游动物在表层大量群聚。
| [1] | 李建佳.杭州湾岸线演化及稳定性研究[D].杭州:浙江大学, 2013. LI J J. Studies on the evolution and stability of the coast of Hangzhou Bay[D]. Hangzhou:Zhejiang University, 2013. http://cdmd.cnki.com.cn/Article/CDMD-10335-1013186005.htm |
| [2] | 闫欣, 钟俊生, 王明星, 等.杭州湾北部水域仔稚鱼种类组成和多样性研究[J]. 上海海洋大学学报, 2014, 23(5): 765–773. YAN X, ZHONG J S, WANG M X, et al.Study on the species composition and diversity of fish larvae and juveniles in the north of Hangzhou Bay[J]. Journal of Shanghai Ocean University, 2014, 23(5): 765–773. |
| [3] | 徐明德, 刘强.杭州湾金山嘴海域水环境质量分析[J]. 科技情报开发与经济, 2006, 16(23): 188–189. XU M D, LIU Q.Analysis on the quality of Jinshanzui water area environment of the Hangzhou Gulf[J]. Sci-Tech Information Development & Economy, 2006, 16(23): 188–189. |
| [4] | 徐兆礼, 沈新强, 袁骐, 等.杭州湾洋山岛周围海域浮游动物分布特征[J]. 水产学报, 2003, 27(S1): 69–75. XU Z L, SHEN X Q, YUAN Q, et al.Distribution characteristics of zooplankton in waters around Yangshan Islands in Hangzhou Bay[J]. Journal of Fisheries of China, 2003, 27(S1): 69–75. |
| [5] | REID P C, BATTLE E J V, BATTEN S D, et al.Impacts of fisheries on plankton community structure[J]. Ices Journal of Marine Science, 2000, 57(3): 495–502. DOI:10.1006/jmsc.2000.0740 |
| [6] | PARK G S, MARSHALL H G.Estuarine relationships between zooplankton community structure and trophic gradients[J]. Journal of Plankton Research, 2000, 22(1): 121–136. DOI:10.1093/plankt/22.1.121 |
| [7] | KENNISH M J. Ecology of estuaries:anthropogenic effects[M]. Boca Raton: CRC press, 1991. |
| [8] | 郭沛涌, 沈焕庭, 刘阿成, 等.长江河口浮游动物的种类组成、群落结构及多样性[J]. 生态学报, 2003, 23(5): 892–900. GUO P Y, SHEN H T, LIU A C, et al.The species composition, community structure and diversity of zooplankton in Changjiang estuary[J]. Acta Ecologica Sinica, 2003, 23(5): 892–900. |
| [9] | 章飞燕, 唐静亮, 李道季, 等.夏、秋季长江口及毗邻海域浮游动物的分布与变化[J]. 水生生物学报, 2009, 33(6): 1219–1225. ZHANG F Y, TANG J L, LI D J, et al.Zooplankton distribution and variation in the Yangtze estuary and its adjacent waters in summer and autumn[J]. Acta Hydrobiologica Sinica, 2009, 33(6): 1219–1225. |
| [10] | WOOLDRIDGE T H, CALLAHAN R.The effects of a single freshwater release into the Kromme Estuary. 3:Estuarine zooplankton response[J]. Water S A, 2000, 26(3): 311–318. |
| [11] | SWADLING K M, BAYLY I A E.Different zooplankton communities in confluent waters:comparisons between three marine bays in Victoria, Australia[J]. Proceedings of the Royal Society of Victoria, 1997, 107: 113–118. |
| [12] | 王丽, 王保栋, 陈求稳, 等.三峡三期蓄水后长江口海域浮游动物群落特征及影响因子[J]. 生态学报, 2016, 36(9): 2505–2512. WANG L, WANG B D, CHEN Q W, et al.Characteristics of the zooplankton communty and impactfactors in the Yangtze estuary coastal eara after third stage impoundment of the Three Gorges Dam[J]. Acta Ecologica Sinica, 2016, 36(9): 2505–2512. |
| [13] | 蔡榕硕, 陈际龙, 黄荣辉.我国近海和邻近海的海洋环境对最近全球气候变化的响应[J]. 大气科学, 2006, 30(5): 1019–1033. CAI R S, CHEN J L, HUANG R H.The response of marine environment in the offshore area of China and its adjacent ocean to recent global climate change[J]. Chinese Journal of Atmospheric Sciences, 2006, 30(5): 1019–1033. |
| [14] | 杜明敏, 刘镇盛, 王春生, 等.中国近海浮游动物群落结构及季节变化[J]. 生态学报, 2013, 33(17): 5407–5418. DOI:10.5846/stxb DU M M, LIU Z S, WANG C S, et al.The seasonal variation and community structure of zooplankton in China sea[J]. Acta Ecologica Sinica, 2013, 33(17): 5407–5418. DOI:10.5846/stxb |
| [15] | 朱根海, 徐汉祥, 陈全震, 等.舟山渔场及附近海域浮游动物指示种的研究[C]//中国环境科学学会, 2010学术年会论文集.上海:中国环境科学学会. 2010. ZHU G H, XU H X, CHEN Q Z, et al. Study on the zooplankton indicator species in the Zhou Shan ground and its adjacent area[C]//China environmental science society, academic essays. Shanghai, 2010. |
| [16] | 刘玉, 李适宇, 董燕红, 等.珠江口伶仃水道浮游生物及底栖动物群落特征分析[J]. 中山大学学报(自然科学版), 2001, 40(s3): 114–118. LIU Y, LI S Y, DONG Y H, et al.Community characteristics of plankton and benthos in the lingding waterway of the pearl river mouth[J]. Acta Scientiarum Naturalium Universitatis Sunyatseni, 2001, 40(s3): 114–118. |
| [17] | PALOMARES-GARCI R, GOMEZ-GUTIERREZ J.Copepod community structure at bahia magdalena, mexico during el niño 1983-84[J]. Estuarine, Coastal and Shelf Science, 1996, 43(5): 583–595. DOI:10.1006/ecss.1996.0089 |
| [18] | 黄备, 吴健平, 唐静亮, 等.杭州湾浮游动物群落与水团的相关性研究[J]. 海洋学报, 2010, 32(1): 170–175. HUANG B, WU J P, TANG J L, et al.The study on the correlation between zooplankton community and water mass in the Hangzhou Bay[J]. Acta Oceanologica Sinica, 2010, 32(1): 170–175. |
| [19] | 姜会超, 陈海刚, 宋秀凯, 等.莱州湾金城海域浮游动物群落结构及与环境因子的关系[J]. 生态学报, 2015, 35(22): 7308–7319. JIANG H C, CHEN H G, SONG X K, et al.Zooplankton community structure in Jincheng area of Laizhou Bay and its relationship with environmental factors[J]. Acta Ecologica Sinica, 2015, 35(22): 7308–7319. |
| [20] | DERISIO C, ALEMANY D, ACHA E M, et al.Influence of a tidal front on zooplankton abundance, assemblages and life histories in Península Valdés, Argentina[J]. Journal of Marine Systems, 2014, 139: 475–482. DOI:10.1016/j.jmarsys.2014.08.019 |
| [21] | 刘镇盛.长江口及其邻近海域浮游动物群落结构和多样性研究[D].青岛:中国海洋大学, 2012. LIU Z S. Community Structure and Biodiversity of Zooplankton in the Changjiang Estuary and its Adjacent Waters[D]. Qingdao:Ocean University of China, 2012. http://www.oalib.com/references/19619630 |
| [22] | 余兴光, 刘正华, 马志远. 九龙江河口生态环境状况与生态系统管理[M]. 北京: 海洋出版社, 2012. YU X G, LIU Z H, MA Z Y. The state of ecological environment and ecosystem management of Jiulong River Estuary[M]. Beijing: China Ocean Press, 2012. |
| [23] | 魏永杰, 王晓波, 张海波, 等.2004~2010年夏季杭州湾鱼类浮游生物种类组成与数量分布[J]. 台湾海峡, 2012, 31(4): 501–508. WEI Y J, WANG X B, ZHANG H B, et al.Ichthyoplankton species composition and distribution in Hangzhou Bay in summer during 2004~2010[J]. Journal of Oceanography in Taiwan Strait, 2012, 31(4): 501–508. |
| [24] | MONTELEONE D M.Seasonality and abundance of ichthyoplankton in Great South Bay, New York[J]. Estuaries, 1992, 15(2): 230–238. DOI:10.2307/1352697 |
| [25] | AZEITEIRO U M, BACELAR-NICOLAU L, RESENDE P, et al.Larval fish distribution in shallow coastal waters off North Western Iberia (NE Atlantic)[J]. Estuarine, Coastal and Shelf Science, 2006, 69(3/4): 554–566. |
| [26] | 庄平, 罗刚, 张涛, 等.长江口水域中华鲟幼鱼与6种主要经济鱼类的食性及食物竞争[J]. 生态学报, 2010, 30(20): 5544–5554. ZHUANG P, LUO G, ZHANG T, et al.Food comparison among juvenile Acipenser sinensis and other six economic fishes in the Yangtze estuary[J]. Acta Ecologica Sinica, 2010, 30(20): 5544–5554. |
| [27] | 郭斌, 张波, 金显仕.黄海海州湾小黄鱼幼鱼的食性及其随体长的变化[J]. 中国水产科学, 2010, 17(2): 289–297. GUO B, ZHANG B, JIN X S.Diet composition and ontogenetic variation in feeding habits of juvenile small yellow croaker Pseudosciaena polyactis Bleeker in the Yellow Sea[J]. Journal of Fishery Sciences of China, 2010, 17(2): 289–297. |
| [28] | 孟田湘.黄海中南部鳀鱼各发育阶段对浮游动物的摄食[J]. 海洋水产研究, 2003, 24(3): 1–9. MENG T X.Studies on the feeding of anchovy (Engraulis japonicus) at different life stages on zooplankton in the Middle and Southern Waters of the Yellow Sea[J]. Marine Fisheries Research, 2003, 24(3): 1–9. |
| [29] | TAKAGI K, YATSU A, ITOH H, et al.Comparison of feeding habits of myctophid fishes and juvenile small epipelagic fishes in the western North Pacific[J]. Marine Biology, 2009, 156(4): 641–659. DOI:10.1007/s00227-008-1115-8 |
| [30] | 纪焕红, 叶属峰.长江口浮游动物生态分布特征及其与环境的关系[J]. 海洋科学, 2006, 30(6): 23–30. JI H H, YE S F.Ecological distribution characteristics of zooplankton and its relationship with environmental factors in the Changjiang River estuary[J]. Marine Sciences, 2006, 30(6): 23–30. |
| [31] | 张冬融, 徐兆礼, 徐佳奕, 等.杭州湾不同水域秋季浮游动物群落结构及其影响因素[J]. 应用生态学报, 2015, 26(9): 2859–2866. ZHANG D R, XU Z L, XU J Y, et al.Zooplankton community structure in relation to influencing factors in different parts of Hangzhou Bay in autumn[J]. Chinese Journal of Applied Ecology, 2015, 26(9): 2859–2866. |
| [32] | 黄备, 王婕妤, 沈明富, 等.浙江北部海域春季浮游动物的群落结构研究[J]. 中国环境监测, 2012, 28(4): 64–68. HUANG B, WANG J Y, SHEN M F, et al.Community structure of zooplankton in the offshore water of the northern Zhejiang[J]. Environmental Monitoring in China, 2012, 28(4): 64–68. |
| [33] | 王婕妤, 唐静亮, 胡颢琰, 等.浙江近岸海域浮游动物的生态分布特征[J]. 浙江海洋学院学报:自然科学版, 2008, 27(4): 384–389. WANG J Y, TANG J L, HU H Y, et al.Ecological distributional characteristics of zooplankton in Zhejiang sea area estuary[J]. Journal of Zhejiang Ocean University:Natural Science, 2008, 27(4): 384–389. |
| [34] | 张丹, 王淼, 孙振中, 等.长江河口区浮游动物生态特征研究[J]. 水产科技情报, 2014, 41(2): 91–94. ZHANG D, WANG M, SUN Z Z, et al.The study on zooplankton ecological characteristics in the Changjiang river Estuaring area[J]. Fisheries Science & Technology Information, 2014, 41(2): 91–94. |
| [35] | 徐兆礼, 王云龙, 陈亚瞿, 等.长江口河口锋区浮游动物生态研究Ⅲ优势种的垂直分布[J]. 中国水产科学, 1995, 2(1): 64–70. XU Z L, WANG Y L, CHEN Y Q, et al.An ecological study on zooplankton in plume frontal zone of Changjiang (Yangtze) river estuarine area Ⅲ vertical distribution of dominant species[J]. China fisheries science, 1995, 2(1): 64–70. |
| [36] | MCMANUS M A, CHERITON O M, DRAKE P J, et al.Effects of physical processes on structure and transport of thin zooplankton layers in the coastal ocean[J]. Marine Ecology Progress Series, 2005, 301: 199–215. DOI:10.3354/meps301199 |
| [37] | CHERITON O M, MCMANUS M A, HOLLIDAY D V, et al.Effects of mesoscale physical processes on thin zooplankton layers at four sites along the west coast of the U. S.[J]. Estuaries and Coasts, 2007, 30(4): 575–590. DOI:10.1007/BF02841955 |
2. Law Enforcement Team, Agriculture Commission of Pudong District, Shanghai 201308, China